我國豬源和雞源大腸桿菌抗生素耐藥性相關基因研究進展
2021-08-13 02:37:34周芷錦沈紅霞倪柏鋒張恩寶曲道峰趙靈燕
中國獸藥雜志 2021年7期
關鍵詞:耐藥
周 煒,周芷錦,沈紅霞, 倪柏鋒, 王 彬, 陳 凱, 穆 琳, 陳 勇, 張恩寶,曲道峰, 趙靈燕*
(1.浙江省動物疫病預防控制中心, 杭州 311119;2:浙江工商大學食品與生物工程學院,杭州 310018)
大腸桿菌(Escherichiacoli,E.coli)是腸桿菌科細菌中分布最廣泛的革蘭氏陰性菌,也是獸醫臨床中常見病原菌之一。自1885年德國小兒科醫師Escherich首次發現大腸桿菌以來,先后有279種血清型被證實[1]。正常情況下,大腸桿菌是動物腸道的共生菌,但當動物機體免疫力低下或菌群失調時,腸侵襲性(EIEC)、腸出血性(EHEC)、腸致病性(EPEC)、腸黏附性(EAEC)、彌散粘附性(DAEC)和腸產毒性(ETEC)大腸桿菌等致病性大腸桿菌可引發宿主腸道、呼吸道、血液系統、生殖系統等多器官系統疾病,例如受到廣泛關注的腸出血性家族的O157:H7型、O2血清型等。由于血清型復雜,且極易產生變異和耐藥性[2],大腸桿菌引發的疾病并無理想的疫苗可用于預防,獸醫臨床多用抗菌藥物進行治療。按照《中華人民共和國獸藥典 獸藥使用指南 化學藥品卷》的分類,目前獸醫臨床常用于豬、雞大腸桿菌病防治的藥物主要有β-內酰胺類(阿莫西林、頭孢噻呋)、氨基糖苷類(新霉素、安普霉素)、喹諾酮類(恩諾沙星)、磺胺類(磺胺氯噠嗪鈉)、多肽類(硫酸黏菌素)和酰胺醇類(氟苯尼考),以及其他合成抗菌類藥物(乙酰甲喹)。在長期抗菌藥選擇進化過程中,大腸桿菌通過非遺傳性耐藥/藥物耐受[3]和耐藥基因遺傳/轉移[4],從而產生水解酶/鈍化霉、抗菌藥物靶位改變、胞吞/胞吐作用改變、形成生物被膜及細胞膜通透性改變等變化,進而產生抗生素耐藥性。因而,大腸桿菌被視作耐藥性指示菌和耐藥基因庫被廣泛研究。目前,已有大量關于動物源大腸桿菌耐藥性相關基因的研究,但由于各研究者的關注點不同,研究中所檢測的耐藥基因也不相同。本文擬根據耐受抗菌藥物種類,對這些基因作用機理及分布情況等方面進行綜述,闡明大腸桿菌的耐藥基因類型,分析比較各類耐藥基因在全國各地的分離檢出率。
1 介導β-內酰胺類抗菌藥物耐受的相關基因
表達產生超廣譜β-內酰胺酶(Extended-spectrum β-lactmases,ESBLs),是大腸桿菌對β-內酰胺類藥物耐藥最主要的機制,還可引起對氨基糖苷類、喹諾酮類藥物、多西環素和氟苯尼考等藥物耐藥,引發了科學界最廣泛的關注[5]。國內獸醫界關注的產ESBLs基因型主要有:TEM、CTX-M、SHV、OXA。
1.1TEM型ESBLsTEM型ESBLs是由TEM-1和TEM-2廣譜酶的編碼基因發生1~4個氨基酸突變而形成的一系列蛋白酶。目前,已知的TEM型酶有115種,是目前數量最多的ESBLs[6-7]。根據徐英春等[8]的綜述,TEM-1、TEM-2、TEM-13和屬窄譜β-內酰胺酶,而TEM-30~36、38~40、44~45、51、59、65、73~78和80屬耐酶抑制劑的β-內酰胺酶,并非ESBLs。關于動物源大腸桿菌中的TEM基因檢測,國內的研究通常不細分至亞型。劉雅妮等[9]對2010~2011年間上海地區296株豬源大腸桿菌的檢測結果表明,52株產ESBLs菌株中TEM基因和CTX-M基因檢出率分別為88.5%和28.8%。曲志娜等[10]發現,2012年山東萊西地區的43株雞源產ESBLs大腸桿菌中CTX-M和TEM基因的檢出率分別為93%和72%。2019年,張明亮等[11]針對豫北地區21株豬源大腸桿菌開展了TEM-1基因檢測,發現檢出率為100%。
1.2CTX-M基因型ESBLsCTX-M基因型ESBLs,是20世紀90年代發現的一類主要作用于頭孢類藥物的ESBLs。其對頭孢噻肟的水解能力最高,同時可水解氨曲南和頭孢曲松,而對頭孢吡肟的水解力很弱。截至2015年,已知的CTX-M基因型ESBLs已達160種(http:∥www.lahey.org/studies/,2015年7月關閉),按基因同源性其可分為5組[12]:CTX-M-1、CTX-M-2、CTX-M-8、CTX-M-9和CTX-M-25。張成東等[13]對2018年從河南省規模化豬場分離得到的704株大腸桿菌的耐藥基因進行了研究,CTX-M、CTX-M-G9和CTX-M-G2的檢出率分別為78.4%、38.4%和69.9%,而CTX-M-G1、CTX-M-G8和CTX-M-G25均未檢出。2015年,楊守深[14]等報道了在213株閩西地區豬源大腸桿菌中17.4%的菌株檢出CTX-M-9G基因,且均與耐喹諾酮類基因共存。李進福等[4]對河南地區31株豬源大腸桿菌的研究表明,CTX-M-U基因檢出率為25.8%,其中CTX-M-1和CTX-M-9亞群菌株均檢出3株。杜向黨等[15]2009年的研究發現:94株雞源大腸桿菌中,CTM-X型基因攜帶率分別為56.4%,CTX-M-9組26株、CTX-M-2組14株、同時攜帶CTX-M-9和CTX-M-2組13株;46株豬源大腸桿菌中,25株檢出CTM-X型基因,CTX-M-9組17株、CTX-M-2組7株、同時攜帶CTX-M-9和CTX-M-2組1株;CTX-M-1、CTX-M-8、CTX-M-25組均未檢出。2016年,張利鋒[16]等發現山東某雞場及其加工場的373株大腸桿菌中,有54.7%的菌株攜帶CTX-M系列耐藥基因,其中CTX-M-55(屬于CTX-M-G1組)、CTX-M-65和CTX-M-14(屬于CTX-M-G9組)檢出率最高,并檢出同于CTX-M-G9組的CTX-M-27基因。張冬冬等[17]2018年的研究結果表明,四川地區29株產ESBLs大腸桿菌中CTX-M的檢出率為100%,如圖1所示,7種亞型的檢出率依次為:CTX-M-65>CTX-M-55>CTX-M-14>CTX-M-79>CTX-M-27>CTX-M-15>CTX-M-123。
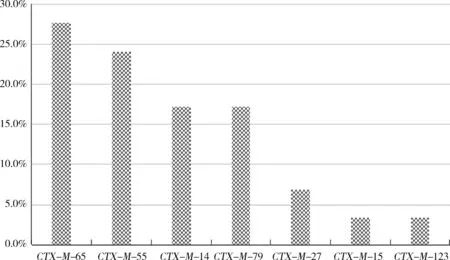
圖1 四川地區7種CTX-M基因亞型的檢出率Fig 1 Ratios of the 7 kinds of CTX-M gene in Sichuan
1.3SHV型基因型ESBLsSHV型β-內酰胺酶是由廣譜的SHV-1酶編碼基因發生1~4個氨基酸突變而形成的,一系列可不同程度水解頭孢菌素類和氨曲南的蛋白酶。目前已經發現39種SHV型酶,其中SHV-1屬窄譜β-內酰胺酶。李進福等[4]發現2015年間河南地區31株豬源大腸桿菌中SHV基因檢出率為61.3%,曲志娜等[10]對2013年山東萊西地區分離得到的49株豬源和53株雞源大腸桿菌的研究發現,產ESBLs大腸桿菌的流行基因主要為CTX-M和TEM,而SHV基因非主要流行基因。2016年,張炳亮等[18]等在洛陽某豬場分離得到的17株致病性大腸桿菌中均未檢出SHV基因。
1.4OXA型基因型ESBLs 目前已知的OXA型β-內酰胺酶有40種,僅16.1%的亞型是從大腸桿菌中分離得到的[11]。OXA-10和OXA-35屬窄譜β-內酰胺酶,OXA-23~27屬碳青酶烯酶。2018年,張成東等[13]在河南地區704株豬源大腸桿菌中,檢出173株攜帶OXA基因的菌株,檢出率低于攜帶CTM-X(552株)和TEM(231株)基因株,高于SHV基因株(18株)。王豪舉[1]對2014~2016年間重慶地區分離到854株雞源和493株豬源大腸桿菌進行的ESBLs基因分析,發現盡管僅1株豬源菌單純攜帶OXA-10基因,但攜帶OXA-10基因的菌株占產ESBLs菌株的9.8%,低于TEM株(占比94.5%)和CTX-M株(占比26.7%),高于SHV株(占比6.9%)。
盡管各研究中時間、地域、動物種類和探討的基因亞型各有不同,總體而言,我國產ESBLs動物源大腸桿菌中TEM和CTX-M為優勢基因型。
除上述4類基因型外,還有關于PER、VEB、GES、CARB、IMP、VIM、DHA、PSE(PSE-1)、BES、STO、TLA等基因的研究報道[1]。此外,李慶周等[19]在2013~2015年間四川地區20個規模化養雞場分離得到的367株雞源大腸桿菌進行了研究,發現33株產CMY-2的菌株,在這些菌株中TEM-1等耐藥基因的檢出率均在70%以上,且表現出對氨芐西林、阿莫西林/克拉維酸100%的耐藥性。盧亞蘭等[20]分析了2018年浙江和陜西地區分離得到的2株攜帶NDM-5質粒的禽源大腸桿菌的耐藥性,2菌株檢出10種和16種耐藥基因,分別在體外表現出對10種和12種抗菌藥物耐藥,耐受的藥物包括頭孢他定、頭孢噻呋和氨芐西林等β-內酰胺。
2 介導氨基糖苷類抗菌藥物耐受的相關基因
大腸桿菌對氨基糖苷類藥物耐受的基因研究,主要集中在16S rRNA甲基化酶和3種產氨基糖苷鈍化酶的編碼基因。16S rRNA甲基化酶可使菌體30S核糖體亞單位中16S rRNA位點的某一個或多個堿基發生甲基化,阻斷了氨基糖苷類藥物與作用靶點的結合,從而產生耐藥性。目前,至少已有rmtA~E、npmA和armA等7種16S rRNA甲基化酶基因被證實。產氨基糖苷鈍化酶的編碼基因是:產O-腺苷轉移酶(aminoglycoside adenylase,AAD)基因、產N-乙酰轉移酶(aminoglycoside acetytranserase,AAC)基因和產O-磷酸轉移酶(aminoglycoside phosphotransferase,APH)基因。國內獸醫學界關注的AAC編碼基因亞型最多,包括:aac(3)-Ⅰ、aac(3)-Ⅱ、aac(3)-Ⅲ、aac(3)-Ⅳ、aac(6')-Ⅰ、aac(6')-Ⅱ、aacA4、aacC2。AAD編碼基因的研究主要集中于aadA1和aadB中,APH編碼基因則主要關注aph(3’)系列。其中,aph(3’)系列包括aph(2'')、aph(3')-Ⅰ、aph(3')-Ⅱa、aph(3')-Ⅲ、aph(3')-Ⅵ等亞型。
鄧向東[21]分析了2012-2013年間四川地區134株豬源和雞源大腸桿菌中rmtA~rmtE、npmA和armA等7種16S rRNA甲基化酶基因,僅5株菌株檢出rmtB基因,其它6種16S rRNA甲基化酶基因均未檢出。劉河冰[22]檢測了2008年間河南鄭州地區120株雞源大腸桿菌中6種16S rRNA甲基化酶基因,結果表明僅檢出armA和rmtB基因,兩者檢出率分別為2.5%和7.5%。
2009年,孫金福等[23]對遼寧地區27株禽源大腸桿菌進行了檢測,發現apH(3’)-Ⅱ、aadA1和aacA4基因檢出率分別為55.6%、44.4%和27.8%。2005年,馬孟根等[24]分析比較了四川地區22株豬源和雞源大腸桿菌中aadA1、aadA2、aadB、apH(3’)-Ⅱ和acc(3)-Ia五種基因的攜帶率,結果表明aadA1和apH(3’)-Ⅱ為優勢基因,兩者檢出率分別為59.1%和54.5%,acc(3)-Ia未檢出。朱永江等[25]所檢測的2015-2016年間揚州地區分離得到的40株豬雞源大腸桿菌中,aac(3)-Ⅱ和aac(6’)-Ib的檢出率分別為17.5%和2.5%。
在多族耐藥基因比較的研究中,張炳亮[18]的研究結果顯示2016年洛陽地區17株豬源大腸桿菌aac(3)-Ⅱ基因檢出率88.2%,高于其他所測試的β-內酰胺類、四環素類和大環內酯類藥物的耐藥基因。2019年,張明亮[11]等的研究發現aadA1和aph(3’)-Ⅱa的檢出率分別為57.1%和76.2%,低于其所試的2種四環素和1種β-內酰胺類耐藥基因,但高于酰胺醇類和磺胺類耐藥基因。
3 介導喹諾酮類抗菌藥物耐受的相關基因
對喹諾酮類耐藥基因的研究主要集中于:質粒介導的喹諾酮類耐藥基質(plasmid mediated quinolone resistance,PMQR)和喹諾酮作用靶位基因決定區的突變基因(Quinolone resistance determining regions,QRDQ)。其中,PMQR基因族又主要包括qnr系列、aac(6’)-Ib-cr、oqxA/B和qepA。嚴格地,oqxAB和qepA均是質粒介導的外排泵基因[26]。
qnr基因通過保護DNA促旋酶和拓撲異構酶Ⅳ不受藥物的抑制來降低菌體對喹諾酮類藥物敏感性,已有報道的qnr主要包括qnrA、B、S、C、D、VC等。楊守深等[14]的研究發現,在2015年福建地區分離得到的213株豬源大腸桿菌中37株檢出CTX-M-9G基因,97.3%的菌株還攜帶有qnr基因,其中qnrS、qnrB和qnrA基因檢出率分別為73.0%、37.8%和5.4%,oqxA、oqxB和qepA的檢出率則依次為70.3%、75.7%和18.9%。aac(6’)-Ib-cr基因易被誤讀為EBSLs編碼基因,其中“-cr”系指環丙沙星耐藥(Ciprofloxacin resistance),是氨基糖苷乙酰轉移酶的新型變異基因編碼的滅活酶。南海辰等[27]對2010~2013年間新疆地區的183株畜源大腸桿菌耐藥基因的檢測結果表明,aac(6’)-Ib-cr的檢出率呈現出一定的動物種屬差異,豬、羊和牛源的檢出率分別為5.0%、22.9%和12.5%。由于豬源、羊源和牛源大腸桿菌菌株數分別為79株、96株和8株,aac(6’)-Ib-cr在不同物種間檢出率的差異可能受樣本量影響。
喹諾酮類藥物作用靶位是DNA促旋酶和拓撲異構酶Ⅳ,DNA促旋酶由gyrA和gyrB基因編碼,拓撲異構酶Ⅳ則由parC和parE基因編碼。因而,QRDQ系列基因的研究圍繞這4個基因開展。張艷芳等[28]的研究發現,DNA促旋酶的GyrA和GyrB亞基上發生的氨基酸替代與喹諾酮類藥物產生耐藥性有關。張秀英[29]等利用中藥處理耐喹諾酮類藥物的大腸桿菌,可使GyrA和ParC亞基從耐藥狀態恢復至敏感狀態。
4 介導四環素類抗菌藥物耐受的相關基因
已知的四環素類耐藥基因,根據其耐藥機制可分為3大類[30-31]:①外排泵基因,通過編碼膜相關蛋白,加速四環素排出細胞漿外。包括:tet(A)~tet(E)、tet(G)、tet(H)、tet(J)、tet(K)、tet(L)、tet(V)、tet(Y)、tet(Z)、tet(30)、tet(31)、tet(33)、tet(35)、tet(38)、tet(39)、otr(B)、otr(C)、tcr和tetA(P)。②核糖體保護蛋白基因,通過編碼核糖體保護蛋白,使核糖體免受四環素作用。包括:tet(M)、tet(O)、tet(Q)、tet(S)、tet(T)、tet(W)、tet(32)、otr(A)和tetP(B)。③滅活/鈍化四環素基因,通過編碼滅活或鈍化四環素的酶而發揮作用。包括:tet(X)、tet(34)和tet(37)。
2018年,王學君[31]對貴州地區783株豬源大腸桿菌中四環素耐藥基因攜帶率進行了研究,發現tetA、tetB、tetC和tetD等4種外排泵基因檢出率分別為92.6%、41.5%、58.8%和58.6%,核糖體保護蛋白基因tetM的檢出率為70.1%,滅活/鈍化四環素基因tetX的檢出率為78.8%。李進福等[4]對2015年間河南地區31株豬源大腸桿菌的外排泵基因(tetA、tetB、tetC、tetK、tetL)和核糖體保護蛋白基因(tetM、tetO、tetW)兩類耐藥基因進行檢測,tetA和tetM基因為優勢基因,兩者檢出率分別為87.1%和54.8%。6株tetL基因檢測陽性,tetC和tetO各有1株呈陽性,未檢出tetB、tetK和tetW基因。在張炳亮[18]等2015年的研究中,豬源大腸桿菌中tetA檢出率為70.6%,而tetC則未檢出。2018年,李蘊玉等[32]對唐山和秦皇島地區22株雞源大腸桿菌中tetA和tetB基因進行了檢測,兩者的檢出率均在80%以上。2016年,張雅為等[33]在遼寧阜新地區33株雞源大腸桿菌中tetB、tetC、tetM、tetK和tetL等5種四環素耐藥基因分析中,33.3%菌株檢出tetK基因,tetL、tetB、tetC、tetM檢出率在15%~30%之間,tetA基因均未檢出。
5 介導酰胺醇類抗菌藥物耐受的相關基因
關于酰胺醇類耐藥基因的研究主要集中在cat、clmA和flor三種基因,此外還有一些關于fexA、cfr、pexA和estdl136的報道。cat基因負責編碼氯霉素乙酰轉移酶,使菌體對氯霉素和甲砜霉素耐藥,但對氟苯尼考沒有作用。cat基因可以分為A型和B型,其中A型又有catⅠ~Ⅳ、catA~D、catP、catS、catQ等16種亞型,B型有cat和catB2種亞型。cmlA和flor均是外排泵蛋白基因,前者只介導氯霉素和甲砜霉素耐藥,而后者則還可以介導對氟苯尼考耐藥[34]。
2019年,張明亮[11]等在豫北地區21株豬源大腸桿菌中發現4株cat1基因陽性,檢出率19.0%。同年,吳方達[35]發現福建地區的17株耐氟苯尼考的大腸桿菌,floR基因檢出率為100%。羅永乾[34]對2014~2016年間重慶地區393株雞源、194株豬源和108株牛源大腸桿菌中cat、clmA、flor、pexA、cfr、fexA和fexB基因進行了檢測,695株菌中各基因總的檢出率依次為:flor(58.7%)>cat(37.0%)>cfr(0.9%),pexA、fexA和fexB均未檢出。
6 介導多粘菌素耐受的相關基因
大腸桿菌耐多粘菌素基因包括:arnBCDTEF基因和mcr系列基因。arnBCDTEF(亦作pmrHFIJKLM)操縱子,其通過脂質A的共價修飾,減少菌體細胞外膜的凈負電荷,從而降低對帶正電荷的多粘菌素的吸附。mcr系列基因屬磷酸乙醇胺轉移酶編碼基因,同樣通過減少菌體細胞外膜的凈電荷而產生耐藥性[36]。自2015年沈建忠院士團隊首次報道mcr-1以來,先后又發現了mcr2~mcr5等基因[37],但獸醫學界尚主要關注mcr-1基因。祝瑤[36]對2016年東三省732株動物源大腸桿菌進行了mcr-1基因攜帶率檢測,發現mcr-1基因在294株雞源大腸桿菌中的檢出率為15.3%,255株豬源菌株中檢出率為9.4%。
7 介導磺胺類抗菌藥物耐受的相關基因
磺胺類藥物主要作用于細菌的二氫蝶酸合成酶(Dihydropteroate synthase,DHFAS)。因而,對大腸桿菌磺胺類藥物耐藥的研究主要集中于DHFAS突變基因folp及DHFAS替代基因。獸醫學界主要關注于后者中的sul1、sul2和sul3基因亞型。2018年,方結紅[38]對浙江地區生豬屠宰環節分離得到的350株大腸桿菌的檢測發現,34株菌檢出sul1、sul2和sul3基因。其中,17株菌存在2~3種基因共存,sul1、sul2和sul3基因對陽性菌株的貢獻率分別為58.8%、47.1%和38.2%。李慶周[19]等發現在2013~2015年間四川地區20個規模化養雞場分離得到的產CMY-2的大腸桿菌中,sul1、sul2和sul3基因檢出率為84.8%、72.7%和36.4%。
8 介導多重藥物耐受的相關基因
除前述的7類藥物的耐藥基因外,國內獸醫學界還對cfr(chloramphenicol-florfenicol resistance)基因、外膜通道蛋白和被膜等有關的耐藥性編碼基因進行了研究。
cfr基因,是因耐受氟苯尼考和氯霉素而得名的2kb左右的小基因,但后經證實其可通過產物-甲基化轉移酶使藥物結合位點發生改變,而可同時介導五類抗生素耐藥。除cfr基因外,大腸桿菌中還存AcrR和Mar等與多重耐藥有關的基因[39]。AcrAB、ACREF、EmrAB、Ecr和QacE等均需于大腸桿菌的主動外排系統,其中AcrAB-TolC外排系統是最主要的。AcrR、MarA、MppA、SdiA、Rob和SoxS等調控基因因子,可作用于AcrAB-TolC外排系統,調節大腸桿菌的外排效能,形成耐藥。馬紅霞等對rob[40]、SdiA[41]基因的研究表明,這2種基因的高水平表達時,均可形成多重耐藥。
MdtM編碼一種大腸桿菌細胞膜蛋白,可非特異性增加抗菌藥物的泵出效率,從而介導多重耐藥。劉嬋嬋[37]在81株人源大腸桿菌中,發現54株攜帶有MdtM基因的菌株,表現出多重耐藥性。ompF編碼大腸桿菌外膜蛋白(OMPs)中的一種親水性非特異孔道蛋白,王藝凝等[42]的研究表明ompF基因通過對被膜及排出泵的作用,而影響大腸桿菌的耐藥性水平。
9 展 望
綜上所述,大腸桿菌不同的耐藥表型對應不同的耐藥基因,同樣的耐藥表型也可能對應不同的耐藥基因,因而在研究目標區域流行的耐藥基因時,需注意三方面因素的影響:首先,各類藥物耐藥機制和基因亞型均具多樣性,選擇哪些基因進行研究分析就顯得尤為重要。其次,從微觀層面的基因、蛋白表達,到宏觀的耐藥表型,還有很多因素在發揮影響,尤其是當目標菌株已表現出多重耐藥時,耐藥基因與表型是否完全一一對應,需審慎地總結。其三,因指示菌與致病菌/繼發感染菌的差異、藥物劑型/佐劑的差異等諸多因素影響,體外試驗的耐藥性與體內試驗結果并不完全一致。
國內獸醫學界對動物源大腸桿菌各類耐藥性相關基因進行深入研究。其中,β-內酰胺類和氨基糖苷類的相關耐藥基因研究較多。氨基糖苷類和四環素類藥物很少用于畜禽大腸桿菌感染的防治,但兩類藥物的耐藥基因檢出率很高,這可能與2方面原因有關:1、依據原《中華人民共和國農業部公告第168號》,氨基糖苷類的越霉素A預混劑、潮霉素B預混劑、硫酸安普霉素預混劑、鹽酸林可霉素及硫酸大觀霉素預混劑、硫酸新霉素預混劑、硫酸安普霉素;四環素類的土霉素鈣和金霉素(飼料級)預混劑,允許作為促動物生長使用。在長期低劑量的四環素類藥物壓力作用下,與動物共生的大腸桿菌自身產生了耐藥性。2、其他種類細菌產生的相關耐藥基因,通過質粒、整合子、轉座子等可移動遺傳元件橫向遷移至大腸桿菌內,再通過繁殖擴增,使這些耐藥基因得以縱向傳播。
隨著國家獸藥規范化使用行動和遏制動物源細菌耐藥性行動的逐步實施與推進,《中華人民共和國農業農村部公告第194號》等法規的相繼頒布與實施,以及養殖主體生物防控意識的不斷增強,動物源細菌耐藥性必將逐步減輕。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
