沿海木麻黃防護林天然更新種群結構和空間分布
2021-08-11 03:20:14江傳陽
福建林業科技 2021年2期
關鍵詞:高峰
江傳陽
(福建省晉江坫頭國有防護林場,福建 晉江 362200)
植物種群結構和空間分布格局是生態學研究的重要內容和核心問題[1-2],能夠反映種群現存狀態、個體組配情況和動態變化趨勢[3-4]。通過徑級結構劃分與統計,可有效分析種群內部的結構和所處階段:幼苗數量多表明種群處于增長的初始期,而當大量中樹出現時,種群處于穩定發展期[5];而幼苗數量少,以中樹、大樹為主,種群結構呈現倒金字塔型,表明種群處于衰退期[2,6-7]。采用生命表研究種群結構,特別是Deevey存活曲線能夠反映種群的動態變化。此外,種群的空間分布格局反映了該物種在漫長的自然選擇過程中每一種群在特定空間會呈現出特有的分布形式[4,8]。木麻黃(Casuarinaequisetifolia)是東南沿海主要防護林樹種,生長迅速、抗強風、耐干旱和鹽堿,不怕沙埋,萌芽力強。早期沿海防護林人工栽植方式通常采用苗栽穴種均勻種植,以快速得到生長狀態相似的成片林分,但因是固定間隔,同齡苗成林,不能反映林分在沿海環境下種群結構和空間分布以及林分的更新規律。因沿海環境惡劣,木麻黃雖能生存,但實生苗在林下出現和存活困難,天然更新防護林更是不易出現,導致沿海地區天然更新防護林種群結構和空間分布研究較少。前人對木麻黃林種群和群落的研究大多集中在凋落物、土壤理化性質和養分等[9],這既不利于掌握木麻黃種群結構和空間分布格局以及生長特征,也不利于預測種群動態。
面對頻繁的沿海災害造成的防護林不同程度的破壞,漫長海岸線上防護林的維護、補植等每年均要消耗大量的人力、物力和財力[10-11],實現沿海防護林的自我天然更新可以節約國家大量寶貴資源。因此,為更好地了解天然更新沿海防護林內部的種群結構和分布格局,以福建省惠安赤湖國有防護林場內天然更新木麻黃林為研究對象,分析徑級結構、編制種群靜態生命表,探討其空間分布,以期為沿海防護林的保護和可持續經營提供參考。
1 材料與方法
1.1 研究區概況
研究區設在福建省惠安赤湖國有防護林場,24°56′N、118°54′E(圖1),屬海洋性季風氣候,年均氣溫20 ℃,最高氣溫35 ℃,年均無霜期306 d,年均降水量1033 mm,年均日照時間2122 h;土壤以紅壤為底的風沙土為主,沙層厚,肥力低,地形較平坦,平均海拔15 m,是典型的沿海沙荒防護林場。防護林主要樹種為木麻黃,伴有桉樹、相思樹、濕地松等,其中木麻黃種植面積達到220.7 hm2。林場建有全球較大的木麻黃種質資源庫,目前引種世界各地木麻黃共200多種。
圖1 研究樣地位置示意圖
1.2 樣地設置
沿海地區普遍實行對受損的防護林進行每年定期補植,天然更新木麻黃林十分稀少,即便有,面積也較小。本研究所調查木麻黃天然林處于沿海基干林帶,距海岸線約150 m,長勢良好,并未進行人工干擾,屬天然更新的木麻黃純林,更新時間始于2012年末。天然更新方式主要起源于種子萌發,超過總數的90%,低于10%的苗由根孽萌發。在全面踏查的基礎上,于2018年8月采用相鄰格子法選擇典型林分設置20塊樣地,每塊樣方10 m×10 m,總面積2000 m2。考慮到木麻黃處于幼苗階段居多,把每個10 m×10 m的樣地再劃分為4個5 m×5 m的小樣方,共80個小樣方。采用每木檢尺法,記錄樣方內所有個體的胸徑、樹高、冠幅。研究地近海岸,地勢相對平坦,高程差忽略,以初始樣方外側2條相互垂直的邊界為橫、縱坐標軸,首樣方外頂點為原點建立平面直角坐標系,記錄每株木麻黃的相對坐標值(x,y)。
1.3 種群結構分析

1.4 空間分布格局分析
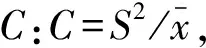
各指標判定種群分布類型見表1。
2 結果與分析
2.1 種群徑級結構和靜態生命表
天然更新木麻黃種群的胸徑最大值≤14 cm,徑級分布可劃分為7級,無間斷,大致趨于反“J”型,各徑級間過渡光滑,無明顯波動(圖2)。其中第Ⅰ級數量最多,占45.5%,有528棵,第Ⅱ

表1 6個指數對種群分布的判定
徑級次之,占27.5%,有319棵,第Ⅰ、Ⅱ徑級共占73%;第Ⅴ、Ⅵ、Ⅶ徑級比例小,僅占6%,其中第Ⅶ徑級占0.7%,只有8棵。

圖2 木麻黃種群徑級結構
死亡率整體上呈波動上升,第Ⅳ級出現第一個死亡高峰,死亡率達到0.54;第Ⅴ級較低,只有0.3;從第Ⅴ級之后死亡率增幅較大;第Ⅵ級達到第2個高峰0.69(表2)。生命期望反映某齡級內個體的生命期望壽命,隨著木麻黃齡級增加呈反比例形式,從第Ⅰ齡級的1.70下降到第Ⅶ齡級的0.69。Ⅶ級不是天然更新木麻黃林最高等級,故死亡率、消失率未作統計。
消失率與死亡率變化趨勢一致,第Ⅳ級出現第一個高峰,達到0.78;最小是第Ⅴ級,為0.35;第Ⅵ級為最高峰達到1.18。其2個消失率高峰與死亡率對應,分別出現在第Ⅳ級和第Ⅵ級。幼苗個體多而大樹少,種群年齡結構呈金字塔形,表明其為增長種群。

表2 木麻黃靜態生命表
存活曲線能反映種群在各個齡級的數目和動態變化,是種群統計中最直觀和常見的方法。一般以齡級為橫坐標,lnlx為縱坐標繪制種群存活曲線。按照Deevey[14]的劃分,種群存活曲線有3種基本類型:Ⅰ型為凸型、Ⅱ型為對角線型、Ⅲ型為凹型[15]。由圖3可知,木麻黃幼苗存活率高,第Ⅰ到第Ⅵ齡級存活數量下降趨勢基本穩定,第V齡級死亡率最低為0.3。木麻黃種群存活曲線屬于Ⅱ型和Ⅲ型中間過渡型。
2.2 種群空間分布格局
20個樣地種群多度方差范圍0~33.54,均值范圍0.25~3.68;擴散系數(C)最大值10.32,最小值0.27。樣地2~5、樣地9~20擴散系數(C)均>1,t檢驗結果顯著,是典型的聚集分布;樣地4、5、9、10、15、17、20擴散系數(C)>1,t檢驗結果不顯著,但負二項分布指數K介于0~8之間,當個體平均密度極高或極低時,t檢驗的可靠性下降,而負二項指數K值不受樣方內個體密度的影響[7],所以它們也屬聚集分布。平均擁擠度M*各樣地均大于x的均值,且聚塊性指數(PAI)各樣地也均>1,僅由平均擁擠度判斷,則各樣地屬聚集分布;但樣地1、6、8的擴散系數(C)<1,聚集指數(CA)<0,負二項分布指數(K)<0,所以應屬均勻分布;而樣地7擴散系數(C)≈1,聚集指數(CA)≈0,負二項分布指數(K)>8,所以屬隨機分布(表3)。

表3 木麻黃種群空間分布格局
3 小結與討論
3.1 天然更新木麻黃種群結構特征
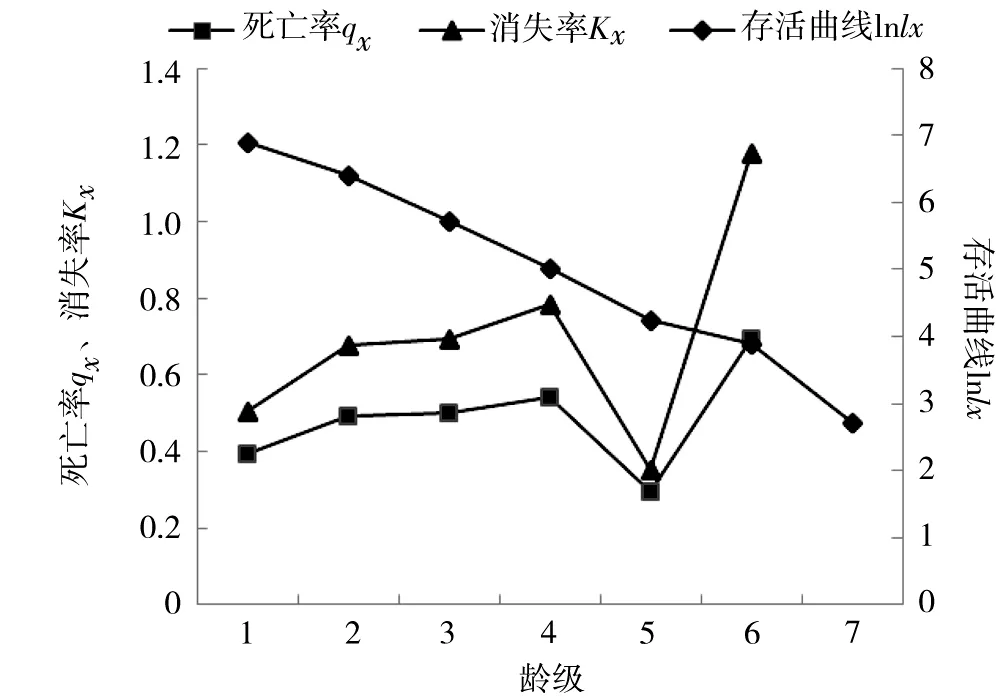
圖3 木麻黃種群死亡率(qx)、消失率(Kx)、存活曲線(lnlx)
年齡結構的分析是表達種群特征的有效方法,也能反映其與環境的相互關系、發展階段、群落地位。本研究中赤湖國有防護林場木麻黃第Ⅰ、Ⅱ齡級占73%,幼苗比例大,后備資源豐富;第Ⅶ齡級比例最小,僅占0.7%。喬木樹種同一環境下同一樹種的齡級與徑級對環境的反應規律具有一致性[16]。種群內中、幼樹多于大樹,種群正處于生命旺盛的中齡期[5]。若種子難以萌發,或種群缺乏幼苗,則呈衰退趨勢[6]。本研究木麻黃徑級結構完整,種群屬于旺盛的增長期。
靜態生命表表達了種群的出生率、死亡率等基本內容,用于壽命長的木本植物種群結構的統計具有特殊意義[17]。所調查木麻黃種群死亡率和消失率變化趨勢基本相同,有2個死亡高峰,這與純木麻黃人工林研究結果[18]一致,但天然更新的木麻黃幼苗死亡率更低,生命力比人工林頑強。木麻黃死亡率第1個小高峰在第IV齡級,幼苗期的死亡高峰往往與種間競爭密切相關,也可能源于環境壓力[19]。研究地多風災鹽蝕,土壤貧瘠,且林分郁閉度高,幼苗面臨光照少、營養缺乏且競爭烈的不利局面,出現死亡高峰并不意外。木麻黃死亡率第2個高峰在Ⅵ齡級,種群個體在第Ⅵ齡級面臨較強的競爭和營養缺乏,水分、空間等生態因子對其生存起到限制作用,個體大量消亡。生命表結果表明:木麻黃對立地要求不高,萌芽能力強,在第1齡級生命期望值最高(表2、圖3)。木麻黃出現第2個死亡高峰可能是由于木麻黃種群在進入中齡期過程中種內競爭加速的結果,本研究木麻黃天然更新始于2012年末,所調查樣地木麻黃種群存活年齡有限,也可能是處于中樹和大樹的分界階段。調查樣地木麻黃幼苗充足,說明其自我更新能力強,在一定條件下可實現天然更新。
3.2 沿海天然更新木麻黃種群空間分布
種群的空間分布格局是物種與環境相互適應的結果,在很大程度上由種群的生物學、生態學特性決定,并受自然環境的影響[20]。在小尺度下,天然更新木麻黃種群與大多數自然種群相同[6,21],空間分布整體屬于聚集分布。木麻黃喜陽性,種子萌芽力強,天然更新主要通過種子繁殖,種子的傳播量與其傳播距離呈反比,易導致種群聚集分布。木麻黃種子較小呈球狀,受海風等外力作用,更易聚集在地面坡度小的凹槽處,不同樣地地表起伏差異不同,種子落地后不易形成均勻分布;坡度和土壤的空間異質性也有可能影響其分布形式,樣地坡度大種群更易聚集分布。此外,木麻黃的根系特點是成網連片,提供強大防護能力的同時可能也是種群聚集分布形成的原因之一。樣地1、6、8的均勻分布和樣地7的隨機分布可能是與相鄰人工林距離過近受到干擾或與土壤養分分布有關。
本研究天然更新木麻黃林在小尺度空間上呈聚集分布,種群個體是斑塊狀分布,有利于充分利用資源,增強種群的競爭力,便于存活,是種群的生存策略和適應機制。木麻黃種群存在2個死亡高峰。成林初期的死亡高峰需格外注意,積極采取保護措施;后期的死亡高峰必要時應加以人工干預,改善生存環境,促使其更好地發揮生態效益。對其種群結構和空間分布格局的研究有利于了解種群所處階段預測未來發展趨勢,可為木麻黃種群的保護和可持續經營提供參考。
天然更新木麻黃林需要一定的條件才能實現,以本研究為例,在其外圍還保存有人工基干林,為其更新提供了一個屏障;林分距海岸線有一定的距離且平均海拔高于海面10多m,不會直接受強海風的強侵蝕,為其更新提供了相對較好的外部環境。四周的人工基干林的存在還是其更新種子的主要來源。由于仍處于更新初期,木麻黃種群徑級分布趨于反“J”形,幼苗數量足,種群處上升階段;但種群存活曲線屬于DeeveyⅡ型和Ⅲ型過渡型;種群具有2個死亡高峰,因此,天然更新的種群動態仍需長期跟蹤監測。小尺度下的聚集分布符合多數天然林種群特征,但由于所調查木麻黃生存年限較短,天然更新林稀缺、面積小,幼樹、中樹、大樹空間分布格局動態變化以及其它尺度下的分布規律仍需進一步研究。
猜你喜歡
瘋狂英語·新讀寫(2021年6期)2021-08-05 07:49:08
今日農業(2021年8期)2021-07-28 05:56:04
書香兩岸(2020年3期)2020-06-29 12:33:45
中華詩詞(2017年1期)2017-07-21 13:49:54
中國核電(2017年1期)2017-05-17 06:09:54
知識經濟·中國直銷(2016年4期)2016-11-07 09:34:15
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:12
衛星與網絡(2016年12期)2016-02-05 09:23:22
中國當代醫藥(2015年33期)2015-03-01 02:09:30
時文博覽(2009年2期)2009-01-01 00:00:00
