山生柳不同優樹無性系光合特性比較
2021-08-11 03:31:38馬成俠李強峰
福建林業科技 2021年2期
馬成俠,李強峰
(1.西寧市南山公園,青海 西寧 810000; 2.青海大學農牧學院農林系,青海 西寧 810016)
山生柳(SalixoritrephaSchneid.)也稱高山柳、毛枝山居柳[1],是中國特有種,落葉矮小灌木,高60~120 cm[2]。生于海拔3000~4700 m的高山地帶,是青藏高原地區高寒柳灌叢的建群種和優勢種。以山生柳作為優勢種所構成的灌木林是青海省主要的灌木林類型之一[1,3],分布廣,面積大,在保持水土、涵養水源等方面具有重要作用[1],是我國干旱半干旱地區優秀的生態防護樹種[4-6]。
青海省地處江河源頭,氣候寒冷、干旱,生態環境脆弱,森林覆蓋率低,多數地方喬木生長受到限制,而山生柳作為青海高寒地區灌木林的建群種或優勢樹種,處于喬木林分布上線與高寒草甸之間,具有生活力強,耐高寒,植株叢生,根系發達的特性。開展山生柳不同無性系的葉綠素含量、凈光合速率和葉綠素熒光等指標的測定與對比分析,揭示無性系間光合特性的規律及其差異性,為山生柳高光效育種篩選優良無性系提供參考,對青海高寒地區植樹造林及生態恢復和生態保護等具有重要意義。
1 試驗材料
本試驗在西寧市大通植物園試驗田進行,所選材料為大通、祁連、果洛、天峻 4個分布區中初選的50個山生柳優良單株的1年生扦插苗。
2 試驗方法
2.1 不同山生柳無性系生長量及成活率測定
2018年3月15日,采用完全隨機區組設計,將50個山生柳無性系制成插條并編號,每個無性系各制30個插條,3次重復,統一用GGR(1∶2000)浸泡3 h后扦插。2019年6—10月進行成活率測定,選出生長穩定的山生柳無性系作為優良無性系。
2.2 不同山生柳無性系光合參數的測定
選擇天氣晴朗的日子,采用LCpro便攜式光合儀測定不同山生柳無性系的光合參數,為減小試驗誤差,每個無性系各測10株,每個所選單株測定3次,分別取上、中、下3個部位中同一朝向的葉片,用均值表示。
2019年9月28日,測定光合有效輻射為1500 μmol·m-2·s-1下的凈光合速率、蒸騰速率、氣孔導度、胞間CO2濃度。9月29日測定所選山生柳無性系的光合參數日變化。測定的時間從7∶00開始,每隔2 h測定1次,直至19∶00結束。
2.3 不同山生柳無性系葉綠素含量的測定
2019年9月28日,在凈光合速率測定后,及時采摘葉片,分別裝袋并帶回實驗室,用研磨法測定。每袋樣品重復3次。葉綠素含量以全部讀數的平均值表示。
2.4 不同山生柳無性系葉綠素熒光參數測定
采用JUNIOR-PAM熒光儀測定山生柳無性系的熒光參數。2019年10月1日,從每個山生柳無性系中選取3株長勢基本一致的植株,選取山生柳枝條第5對葉片,用濕毛巾包裹,放到加有冰袋的保溫箱中,帶回實驗室測定。測定指標包括:初始熒光Fo,最大熒光產量Fm、Fv/Fm、Fv/Fo、qP、qN、NPQ、ETR,每個葉片測定3次,取平均值。
3 數據處理及綜合分析
采用Excel統計軟件對測定數據進行統計和圖表繪制,采用SPSS 24.0軟件進行數據分析。
4 結果與分析
4.1 不同山生柳無性系的生長特性比較
2019年6月對山生柳無性系生長情況進行調查,結果見表1。其中苗高最大的無性系(T11)為113.4 cm,最小的無性系(Q33)為23.5 cm,極差近5倍,總平均值為60.6 cm;地徑最大的無性系(T10)為14.33 mm,最小的無性系(T5)為5.09 mm,極差近3倍,總平均值為8.20 mm。從表中看出,有18個山生柳無性系的成活率大于80%,且苗高和地徑均大于區組平均值,因此將此18個無性系作為光合特性研究對象。同一無性系的生長表現整齊一致,各類形態特征完全統一。
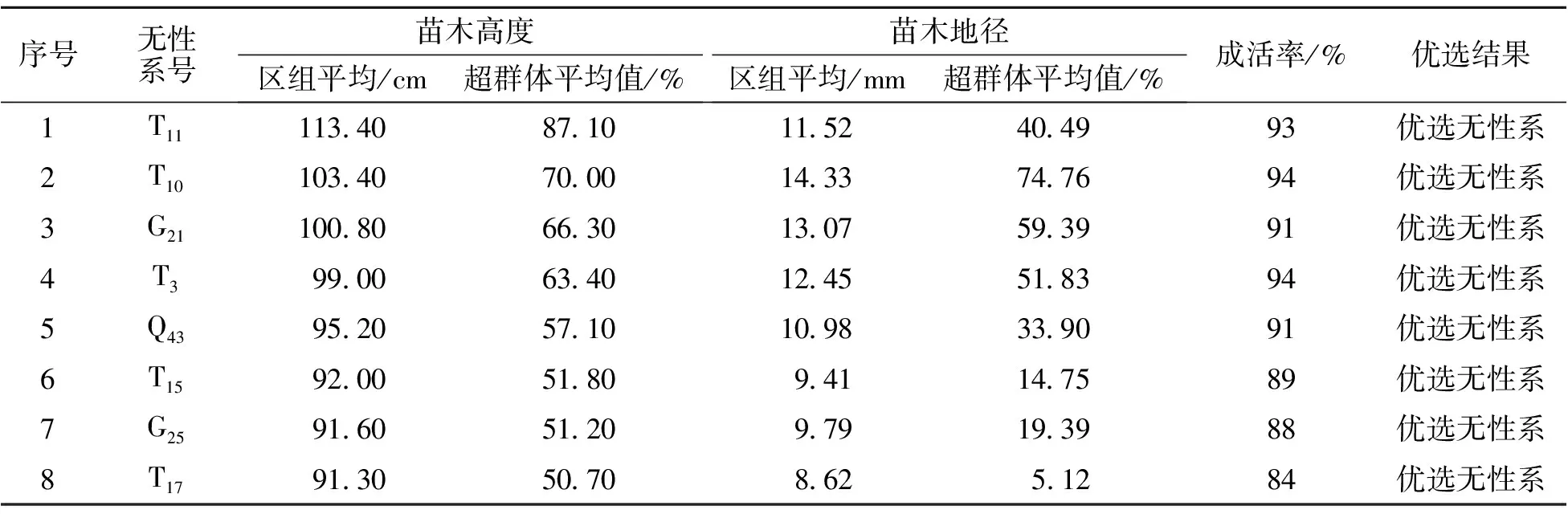
表1 不同山生柳無性系生長狀況
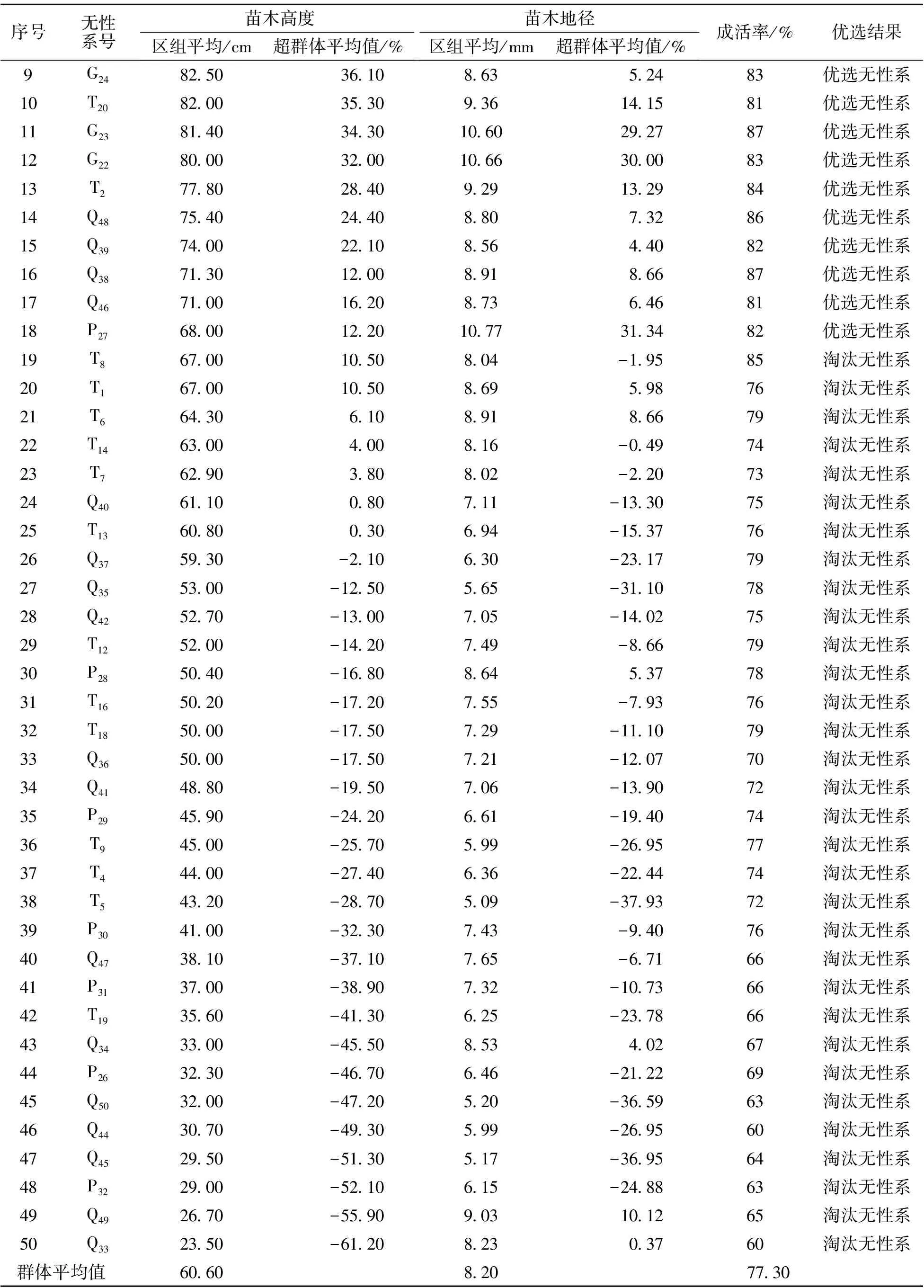
表1(續)
4.2 不同山生柳無性系的光合特性比較
4.2.1 不同山生柳無性系葉綠素含量比較 18個山生柳無性系的葉綠素含量間存在極顯著差異(P<0.01)。對18個山生柳無性系進行多重比較(表2)可知,葉綠素含量較高的是T11、T10、T3、G21、Q43號無性系,分別為7.65、7.55、7、6.91、6.8 mg·g-1,顯著高于T15~T18號無性系。其中T10、T11的葉綠素含量顯著高于其它山生柳無性系,說明T10、T11具有更大的光合潛力。
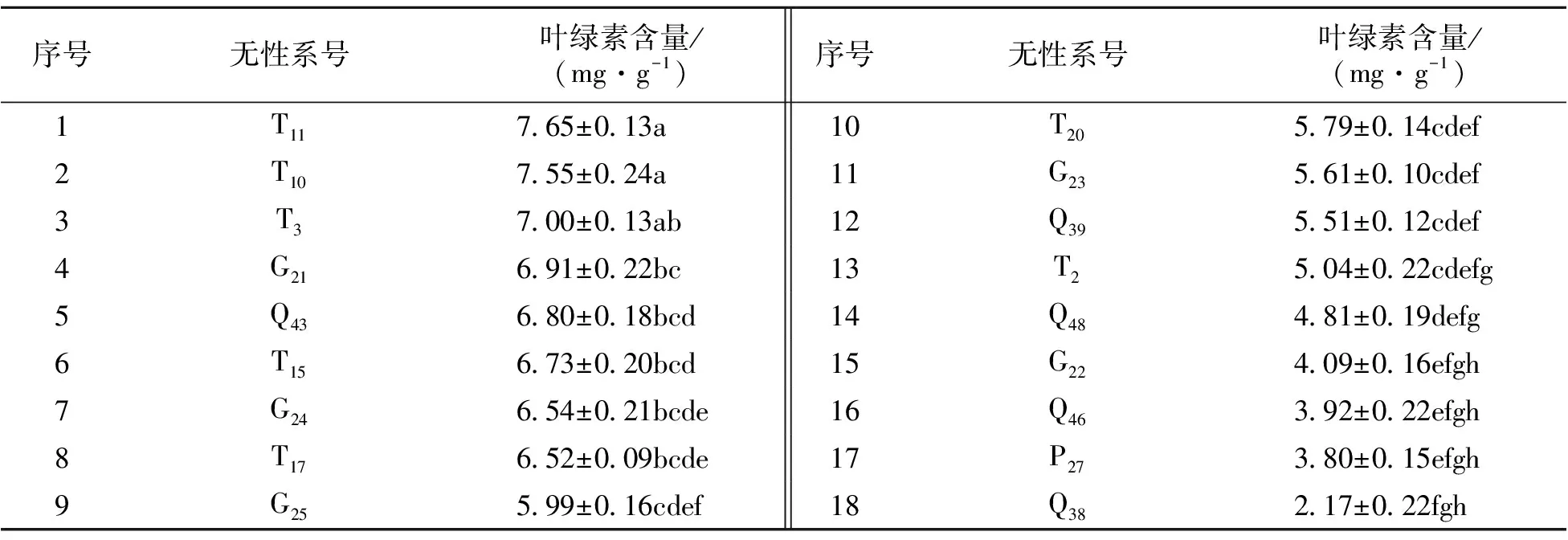
表2 不同山生柳無性系葉綠素含量多重比較
4.2.2 不同山生柳無性系光合參數比較 對18個無性系的光合參數進行多重比較(表3)可知,T11、T10、T3、G21、Q43的凈光合速率Pn值顯著高于其它無性系,分別為5.57、5.38、5.24、5.13、5.04 μmol·m-2·s-1,其中T11仍居于首位,說明這5個山生柳無性系對光能的利用率較高,其中T11的光能利用率最高,且T11與其它無性系間凈光合速率存在顯著性差異(P<0.05)。氣孔導度Gs最大的無性系是T11、T10、T3、G21、Q43,分別為230.5、222.7、218.4、185.7、185.4 μmol·m-2·s-1,顯著高于其它無性系,其中T11、T10、T3之間氣孔導度Gs差異不顯著、G21與Q43之間氣孔導度Gs差異不顯著(P>0.05),但T11、T10、T3與G21、Q43及其它無性系之間的氣孔導度Gs存在顯著性差異(P<0.05)。蒸騰速率Tr較大的無性系是T11、T10、T3、G21,分別為3.16、2.94、2.79、2.74 mmol·m-2·s-1;蒸騰速率較小的無性系是Q38,僅0.86 mmol·m-2·s-1。各山生柳無性系的胞間CO2濃度Ci在129~385 μmol·m-2·s-1之間,其中Q38、P27、Q46、G22、Q48的胞間CO2濃度較高,T11的胞間CO2濃度最低;T11、T10、T3之間胞間CO2濃度差異不顯著(P>0.05),但T11、T10、T3與其它無性系之間的胞間CO2濃度存在顯著性差異(P<0.05)。
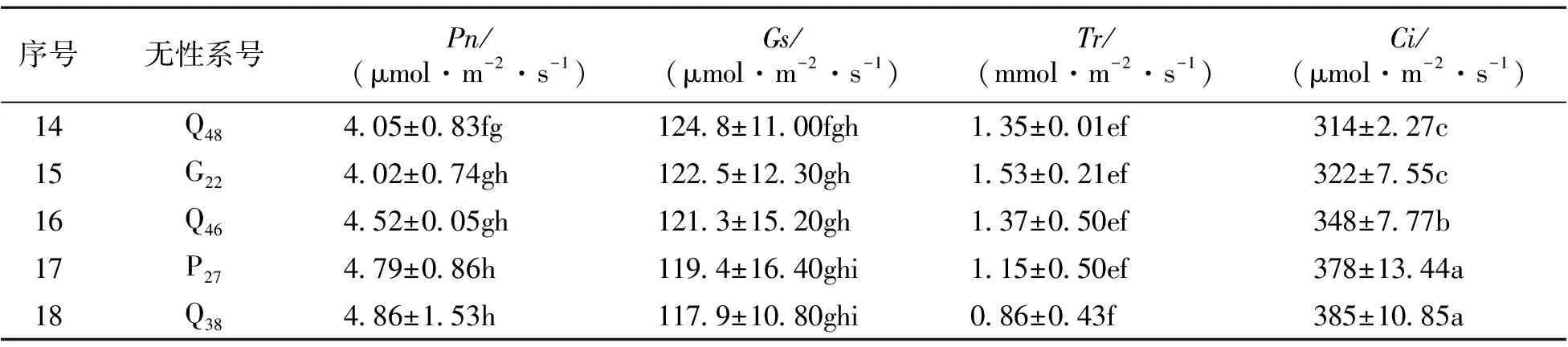
表3(續)
4.2.3 不同山生柳無性系葉綠素含量與凈光合速率聚類分析 葉綠素含量、凈光合速率測量結果表明,山生柳葉片的葉綠素含量與凈光合速率呈顯著正相關(P<0.05)(表4),即在一定范圍內,葉綠素含量越高凈光合速率越強。把葉綠素含量、凈光合速率作為主要性狀指標,對18個山生柳無性系進行多指標系統聚類分析,結果見圖1、表5。當距離為2時,18個山生柳無性系可分為5大類群:類群1包括T3、Q43、G21、T11、T10、G24共6個山生柳無性系,平均葉綠素含量最高,平均凈光合速率最大,分別為7.03 mg·g-1、33.85 μmol·m-2·s-1。說明這6個無性系的光合利用率強,生理活性高;類群2包括T15、T17、G25、T204個無性系,這4個無性系的平均葉綠素含量較高,平均凈光合速率較大,分別為6.26 mg·g-1、28.46 μmol·m-2·s-1;類群3僅有G231個無性系,其葉綠素含量、凈光合速率處于中等水平,分別為5.61 mg·g-1、23.58 μmol·m-2·s-1;類群4包括G22、Q46、Q39、T2、Q485個無性系,這5個無性系的平均葉綠素含量較低、平均凈光合速率較小,分別為4.67 mg·g-1、19.94 μmol·m-2·s-1;類群5包括P27、Q38共2個無性系,這2個無性系的平均葉綠素含量及平均凈光合速率最小,分別為2.99 mg·g-1、14.83 μmol·m-2·s-1,說明這2個山生柳無性系的生理活性較弱。
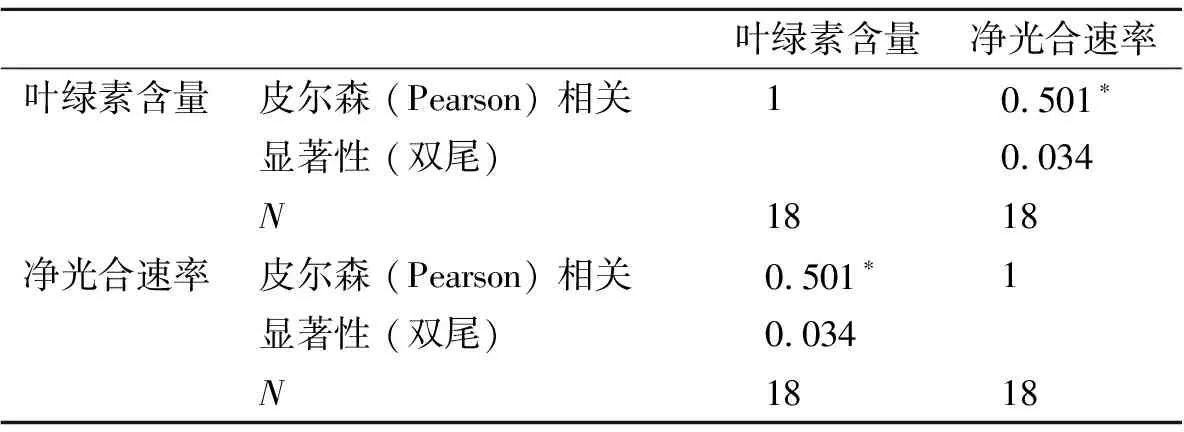
表4 葉綠素含量與凈光合速率相關性分析
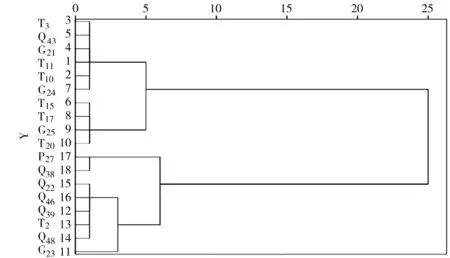
圖1 不同山生柳無性系葉綠素含量與凈光合速率聚類分析
綜上所述,T3、Q43、G21、T11、T10、G24葉綠素含量和凈光合速率最大,說明T3、Q43、G21、T11、T10、G24的生理活性及光合作用較強,有利于植物的物質積累。結合生長特性指標,發現T3、Q43、G21、T11、T10這5個無性系的生長指標及光合指標均高于其它無性系。
4.2.4 不同山生柳無性系葉綠素熒光參數比較 由表6可知,Fo與Fv/Fo、NPQ呈極顯著負相關,與Y(II)、qN、qP呈顯著負相關,與Fm、Fv/Fm、ETR相關性不顯著;Fm與Fv/Fo、NPQ呈極顯著正相關,與Fv/Fm、Y(II)、qN、qP呈顯著正相關,與ETR相關性不顯著;Fv/Fm與Fv/Fo、Y(II)呈極顯著正相關,與NPQ呈顯著正相關,與qN、qP、ETR相關性不顯著;Fv/Fo與Y(II)、NPQ、qN、qP及ETR呈極顯著正相關;Y(II)與NPQ、qP和ETR呈極顯著正相關,與qN呈顯著正相關;NPQ與qN、qP呈極顯著正相關,與ETR相關性不顯著;qN和qP呈極顯著正相關,與ETR相關性不顯著;qP和ETR呈極顯著正相關。
把實際光化學量子產量(Yield)、表觀電子傳遞速率(ETR)和非光化學猝滅系數(NPQ)作為主要熒光參數,對18個山生柳無性系進行多指標系統聚類分析,結果見圖2、表7。當距離為2時,18個山生柳無性系可分為5大類群:類群1包括T11、T10共2個山生柳無性系,其Yield、ETR、NPQ最高,說明這2個無性系光合能力及吸收能力最好,光合活性和對光能利用率最強;類群2包括T3、Q43、G21、T15、G245個無性系,這5個無性系的Yield、ETR、NPQ值相比其它無性系較高,說明光合能力及吸收能力較好,光合活性和光能利用率較強;類群3有T17、G25、T203個無性系,光合能力及吸收能力一般,光合活性和光能利用率一般;類群4包括G23、G22、Q46、Q39、T2、Q486個無性系,這6個無性系的光合能力及吸收能力較差,光合活性和光能利用率較小;類群5包括P27、Q382個無性系,這2個無性系的光合能力及吸收能力最差,其光合活性和光能利用率最低 。
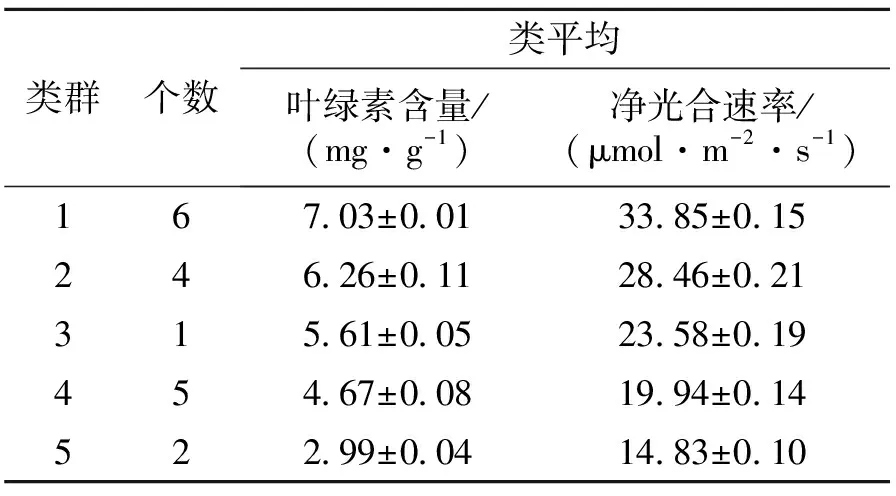
表5 葉綠素含量與凈光合速率聚類分析結果
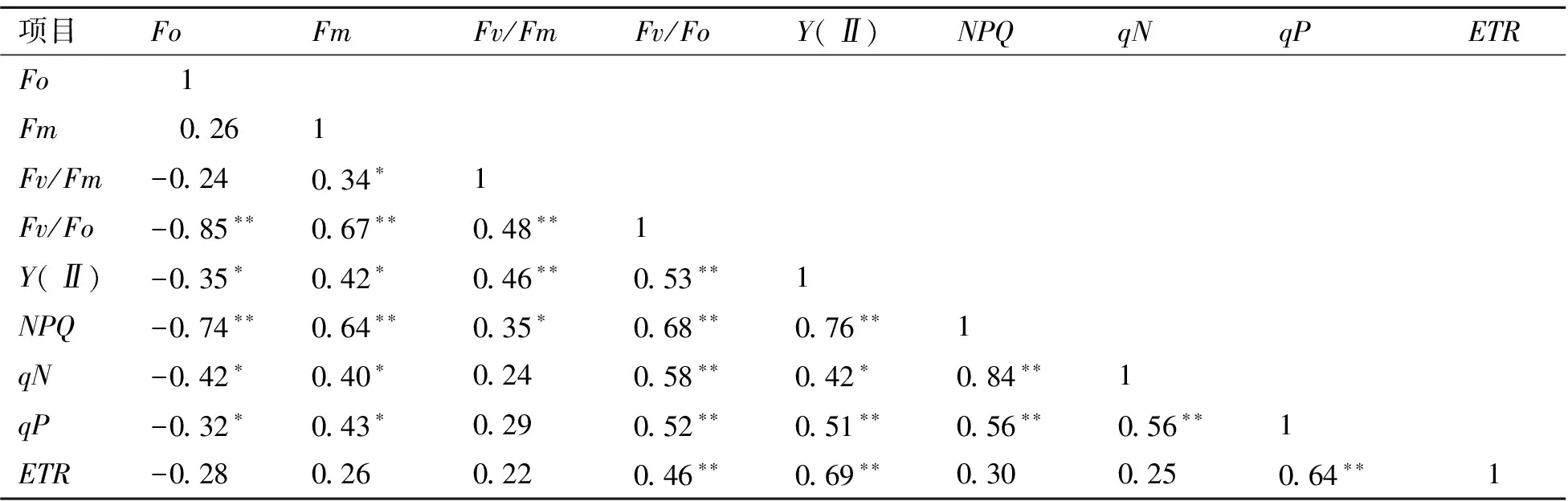
表6 葉綠素熒光參數間相關關系矩陣表
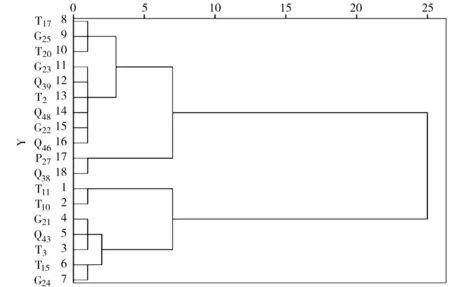
圖2 不同山生柳無性系葉綠素熒光聚類分析
綜上所述,Yield、ETR、NPQ最高的山生柳無性系是T11、T10,最低的山生柳無性系為P27、Q38,與光合特性和生長指標的聚類結果相吻合。說明葉綠素熒光、葉綠素含量和凈光合速率對植物生長具有一定影響,它們之間有一定的內在關系。
4.2.5 不同山生柳無性系光合特性比較 對18個山生柳無性系的葉綠素含量、光合參數及葉綠素熒光參數進行分析可知,T11、T10、T3、G21及Q435個無性系生長指標及光合指標均高于其它無性系且排名基本穩定,可將其作為光合生理育種中的高光合特性無性系資源,故僅對這5個無性系進行光合特性分析即可。由表8可知,不同山生柳無性系的光合參數存在一定差異,而T11的凈光合速率、氣孔導度、蒸騰速率均位居首位,分別為5.19 μmol·m-2·s-1、192.89 μmol·m-2·s-1、3.11 mmol·m-2·s-1,其次分別為T10、T3、G21、Q43。而胞間CO2濃度最大的則是Q43,達到306.29 μmol·m-2·s-1,最小的是T11,僅為207.65 μmol·m-2·s-1。說明T11的光合作用最強,光能利用率也最強。
5 結論與討論
本試驗結果表明,18個山生柳無性系的葉綠素含量間存在極顯著差異,葉綠素含量較高的是T11、T10、T3、G21、Q43號無性系,顯著高于T15之后的其它山生柳無性系。T11的葉綠素含量顯著高于其它山生柳無性系,說明T11具有更大的光合潛力。18個山生柳無性系的葉綠素含量變化及光合特性的聚類結果發現T11、T10、T3、G21、Q43號山生柳無性系的光合活性及光能利用率均顯著大于其他山生柳無性系,因此,可將其作為光合生理育種中的高光合特性無性系資源。不同山生柳無性系的光合日變化參數具有差異,而T11的凈光合速率、氣孔導度及蒸騰速率均位居首位,其次分別為T10、T3、G21、Q43。而胞間CO2濃度最大的則是Q43,最小的是T11,說明T11的光合作用最強,光能利用率也最強。
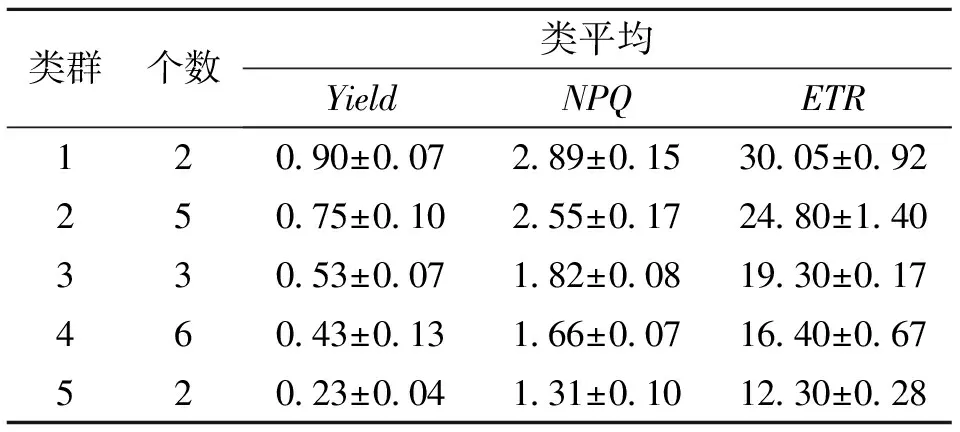
表7 葉綠素熒光聚類分析
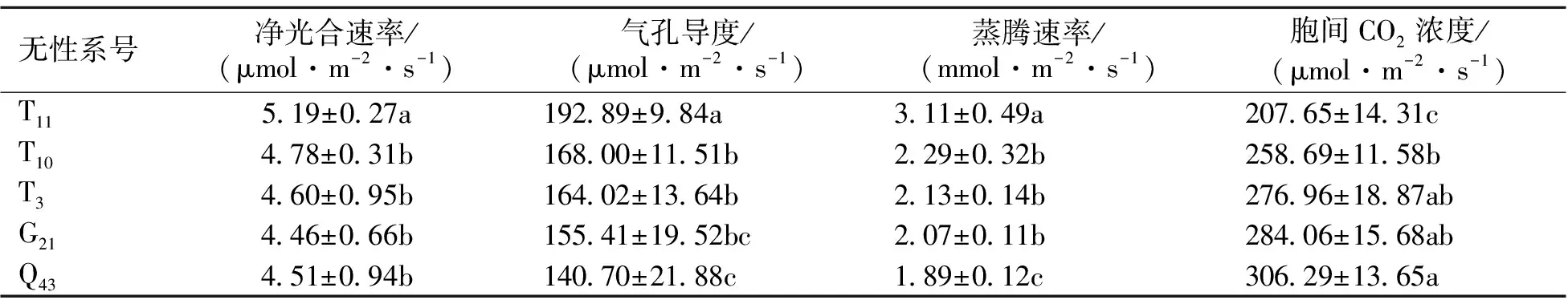
表8 不同山生柳無性系光合特性比較
目前,有關葉綠素含量的研究中,多為鹽脅迫或干旱脅迫對柳樹葉綠素含量影響,而對葉綠素含量與光合特性的相關性研究不多。本試驗通過研究山生柳無性系的生長特性、葉綠素含量及光合特性,發現在同一階段下的葉綠素含量與光合速率呈正相關關系,即在一定范圍內,葉綠素含量越高凈光合速率隨之越強。檉柳的葉綠素含量隨著凈光合速率的升高逐漸增強[7],研究結果與本試驗較為一致。
有關葉綠素熒光分析技術的研究多集中于逆境脅迫[8]、光抑制[9]、干旱脅迫[10]、水分脅迫[11]、鹽分脅迫[12]和其它脅迫[13]等。在山生柳生長發育方面未見相關報道。本文通過葉綠素熒光技術對18個山生柳無性系進行測定可知,葉綠素熒光參數可反映山生柳無性系的生長性狀及光合特性。此研究結果為今后選育山生柳無性系奠定了理論基礎。
