長臂蝦科幾種重要經濟蝦類的繁殖生物學研究進展
2021-08-10 09:06:26胡潤豪史文軍萬夕和王李寶楊澤禹吳旭干
海洋漁業 2021年4期
胡潤豪,史文軍,王 盼,萬夕和,沈 輝,黎 慧,王李寶,楊澤禹,吳旭干
(1.上海海洋大學,水產科學國家級實驗教學示范中心,上海 201306;2.江蘇省海洋水產研究所,江蘇南通 226007;3.中國科學院煙臺海岸帶研究所,山東煙臺 264003)
長臂蝦科(Palaemonoidea)隸屬甲殼綱(Crustacea)十 足 目(Decapoda)真 蝦 次 目(Caridea),其下屬長臂蝦亞科現生種屬共計21屬373種,隱蝦亞科現生種屬共計117屬604種[1]。常見長臂蝦科蝦類主要隸屬于長臂蝦亞科的沼蝦屬(Macrobrachium)或白蝦屬(Exopalaemon),其余各屬雖在我國也有分布,但群體數量較小或未實現大規模人工培育,如古潔蝦屬(Palaemonetes)的廣東古潔蝦(P.tonkinensis)、瘦蝦屬(Leander)的細腳瘦蝦(L.tenuicornis)等[2]。
長臂蝦科蝦類是我國漁業重要的一類經濟蝦類[2],主要的養殖或捕撈品種有:日本沼蝦(M.nipponense)、羅氏沼蝦(M.rosenbergii)、脊尾白蝦(E.carinicauda)、秀麗白蝦(E.modestus)、葛氏長臂蝦(Palaemon gravieri)、海南沼蝦(M.hainanense)、細螯沼蝦(M.superbum)以及安氏白蝦(E.annandalei)[3-4]等,其中養殖規模較大且研究報道較多的有淡水品種日本沼蝦、羅氏沼蝦和海水品種脊尾白蝦。日本沼蝦自然分布于亞洲各地,除中國外,日本、朝鮮半島、越南、馬來西亞以及俄羅斯遠東地區都有分布[5];羅氏沼蝦原產于泰國、印度、緬甸等的河口處,自1976年引入我國后,分別于廣東和廣西的水產研究所進行試養[6]。2018年我國日本沼蝦產量23.4萬t、羅氏沼蝦產量13.3萬t,在淡水養殖蝦類產量排行中分別位列第三和第四位,江蘇省日本沼蝦與羅氏沼蝦產量為全國最高,分別為11.5萬t和6.4萬t[7]。脊尾白蝦主要分布于中國大陸沿岸和朝鮮半島西岸的淺海低鹽水域[5],目前全國養殖面積約3.7萬hm2,其中江蘇省養殖面積最大、產量最高。
1 繁殖生物學特征
1.1 繁殖期
長臂蝦科幾種重要經濟蝦類的繁殖期[8-20]見表1。長臂蝦科蝦類(以下簡稱長臂蝦類)繁殖期較長,一般在5個月以上,繁殖盛期一般在5—8月份,但不同地區的長臂蝦類繁殖期和繁殖盛期存在一定差異,這可能主要是由于溫度和生物餌料量等因素的差異導致。
1.2 繁殖力
產卵量和產卵次數是蝦類繁殖力的主要體現,長臂蝦類產卵量從幾百粒到幾萬粒不等,產卵次數也存在較大差異。長臂蝦類產卵量不僅受 到 卵 巢 發 育 狀 況[11]、體 長[15,21-22]和 體 質量[23-24]等內在因素影響,也容易受到外界環境因素的影響,如溫度[13]、鹽度[25]和季節變化[26]等。多數長臂蝦類一年內可以多次產卵[15,22,27]。長臂蝦類產卵量一般少于對蝦類而多于螯蝦類(除秀麗白蝦外)[18,20,23,26,28-34],這可能是由于雌蝦個體大小以及卵粒直徑差異造成的(表2)。
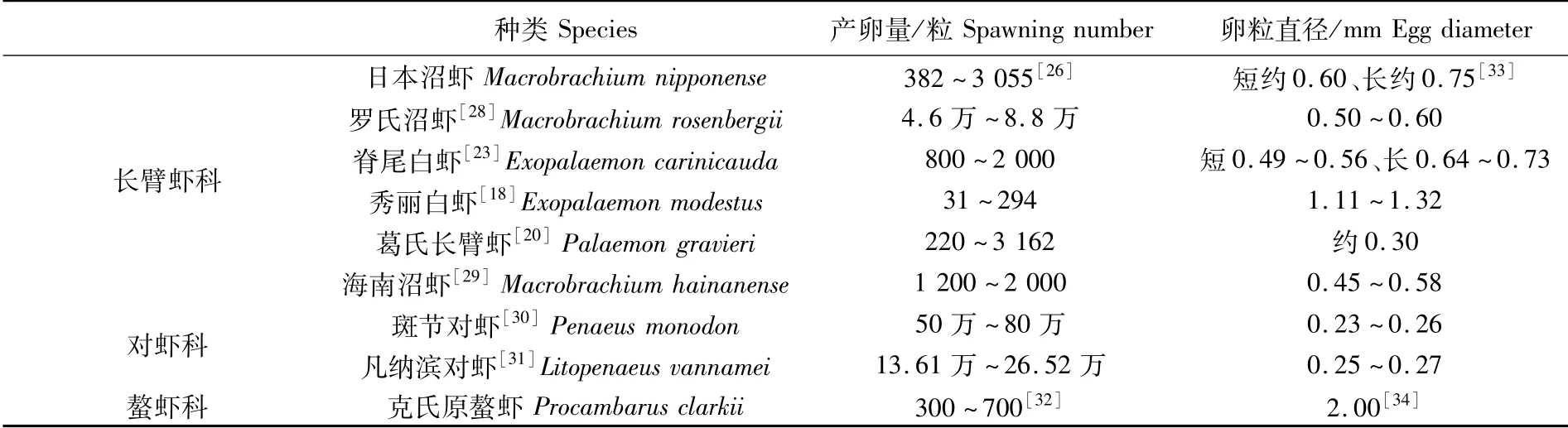
表2 長臂蝦科蝦類與其他科蝦類產卵量及卵徑對比Tab.2 Comparison of spawning number and egg diameter of Palaemonoidea and other families
1.3 雌雄性比
雌雄性比是動物群體構成的重要評價指標,也是繁殖生物學關注的群體特征之一,長臂蝦科幾種重要經濟蝦類雌雄性比見表3。盧敬讓等[21]認為,繁殖季節雌蝦多于雄蝦是因為抱卵雌蝦繁殖洄游,以及越冬期間雄蝦死亡率高于雌蝦,從而導致越冬后群體中雌蝦占比較大,這與何緒剛等[8]和孫建貽等[35]對日本沼蝦性比變化的研究結果一致。另外,在長臂蝦類繁殖期,部分成蝦壽命已經達到上限,群體中雄蝦在交配完成生物學使命后很快死亡,雌蝦則要等待受精卵孵化后死亡[8,36];而雌蝦死亡后,大量當年幼蝦加入群體,緩沖了雌蝦死亡導致的性比變化趨勢。與其他野生長臂蝦類群體相比,引進物種羅氏沼蝦主要為池塘養殖,雖然雌雄性比均大于1,但相對變化較小,這可能是由于自然群體的捕撈在時間和空間上跨度較大,導致數據統計的波動性大,而池塘養殖時間和空間跨度小,數據統計的波動較小。
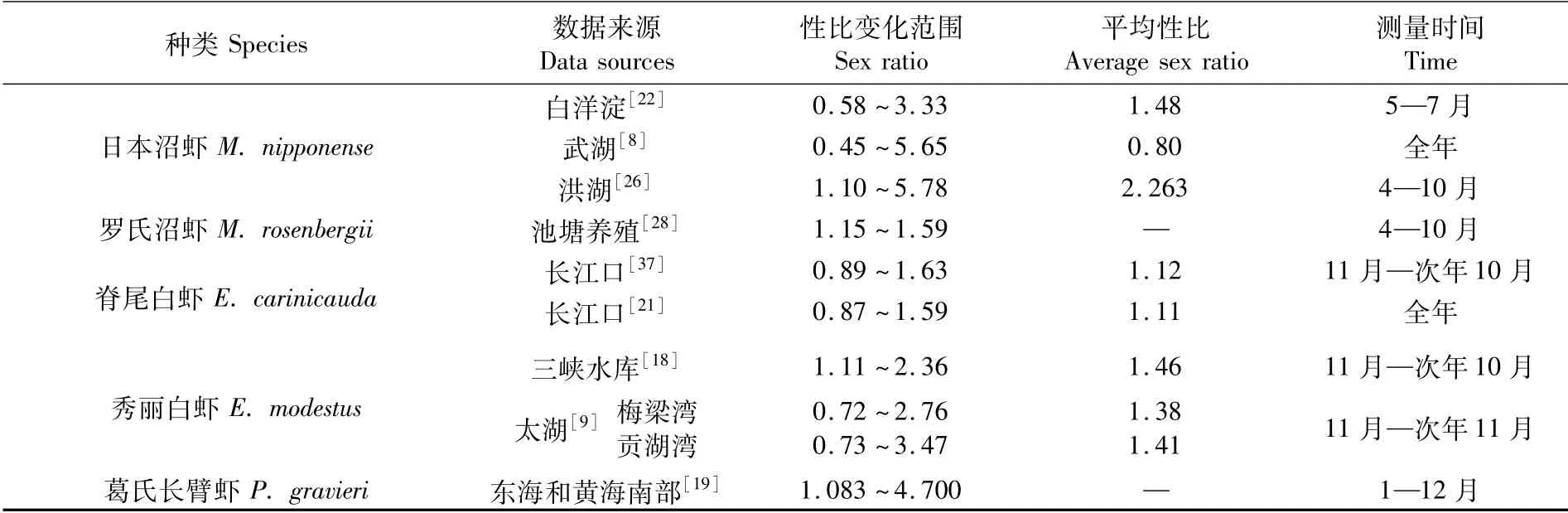
表3 長臂蝦科幾種重要經濟蝦類雌雄性比Tab.3 Sex ratio of some important economic kinds of Palaemonidea
2 生殖系統發育
2.1 雌性生殖系統
2.1.1 雌性生殖系統結構
長臂蝦類雌性生殖系統組成基本相似,都有卵巢與輸卵管,但不同種之間存在一定差異,主要表現在卵巢左右兩葉的連接方式上[36],目前還未見關于這種結構差異可能導致的影響的報道。脊尾白蝦雌性生殖系統由1對卵巢和2根輸卵管組成,整個卵巢呈“O”字型,前后端愈合在一起,中 間 形 成 一 個 空 隙[36],與 粗 糙 沼 蝦(M.asperulum)[38]卵巢相似。羅氏沼蝦卵巢左右兩葉呈桑葚狀,前端愈合,后端分離,卵巢兩側分別有短的輸卵管與生殖孔相連[39-40],與哈氏仿對蝦(Parapenaeopsis hardwickii)[41]卵巢相似。日本沼蝦的卵巢呈橢圓形,前端略尖,后端圓鈍,前部成對,后部合二為一,在卵巢的兩側各有一條短而直的輸卵管[42],與克氏原螯蝦(Procambarus clarkii)[43]的卵巢結構類似。
2.1.2 卵巢發育
研究人員主要根據卵巢顏色、大小或相對位置及內部卵細胞成熟情況將日本沼蝦[42]和脊尾白蝦[15]的卵巢發育分為5個時期,羅氏沼蝦[40]的卵巢發育分為4個時期,見表4。總結3種長臂蝦類卵巢各時期的特點,筆者認為脊尾白蝦卵巢5個發育時期的劃分和命名更為合理。
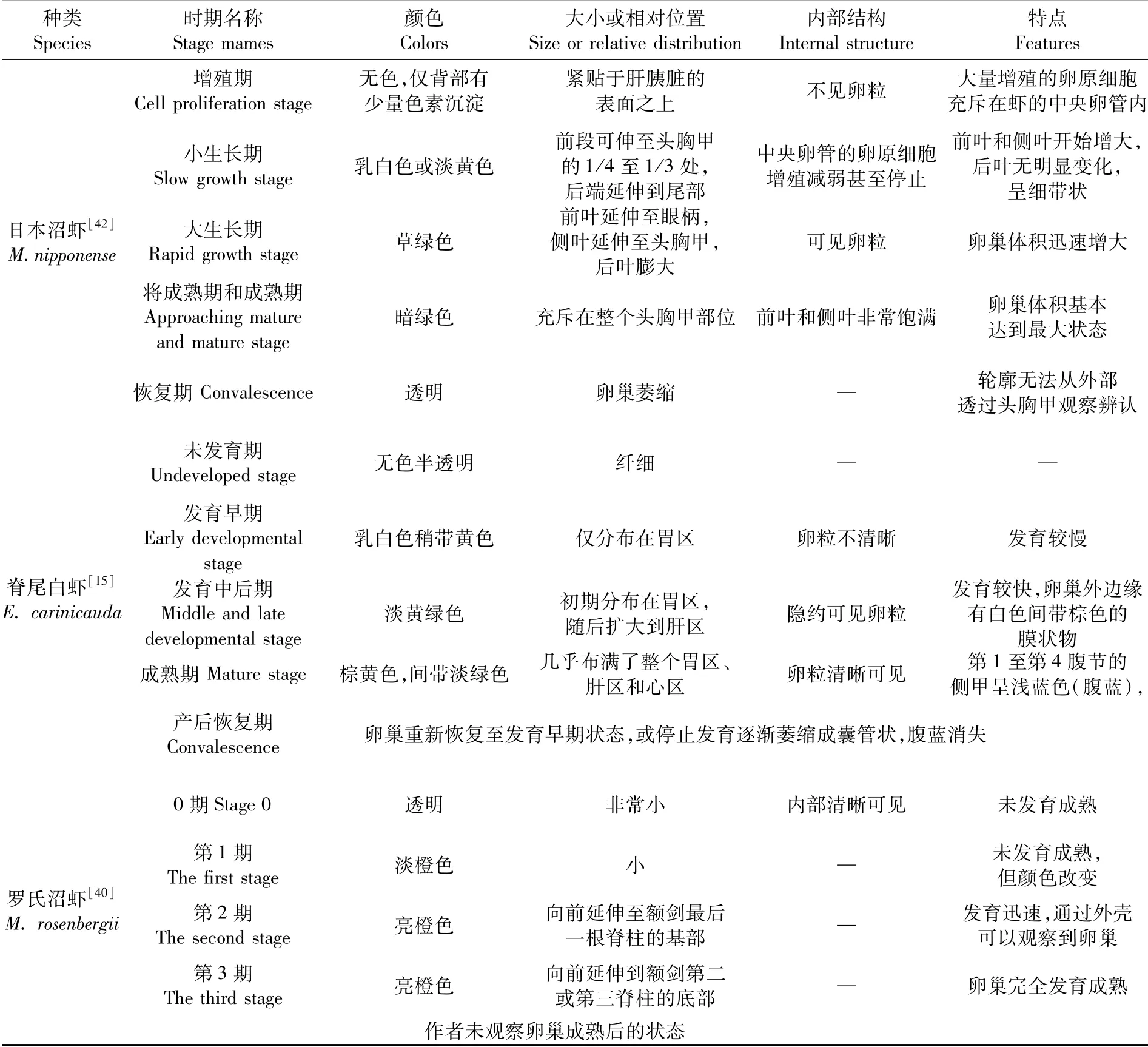
表4 3種長臂蝦類卵巢發育時期及特征Tab.4 Ovarian development stages and characteristics of three species of Palaemonidae
2.2 雄性生殖系統
2.2.1 雄性生殖系統結構
長臂蝦類雄性生殖系統的研究主要集中在精巢結構、精子發生等方面。長臂蝦類的雄性生殖系統由精巢和輸精管組成[44-45],與對蝦類雄性生殖系統存在明顯差異,沒有分泌管和精囊,而這兩種結構往往在對蝦生殖系統中發揮重要作用[47](圖1)。精巢內為無數的生精小管,其管壁由位于基膜上的生精上皮和管壁上皮構成,生精上皮細胞主要為精原細胞,經有絲分裂,一部分精原細胞發育成不同階段的雄性生殖細胞,形成生殖帶[45]。同一區段的生精小管內精子發生的同步率較高,可以保證在較短時間內產生出大量成熟精子;不同區段的精子的發生不同步,使得生殖帶能在較長時期內源源不斷地產生精子,適應于當年生雄蝦繁殖期的多次繁殖[46]。長臂蝦類輸精管末端膨大為壺腹,肌肉層發達,可以暫時貯存包裹著精子的精莢,同時發達的肌肉層能在射精時給予所需的力量,其作用可能類似于對蝦的精囊,起到儲精和射精的雙重功效[44-46]。
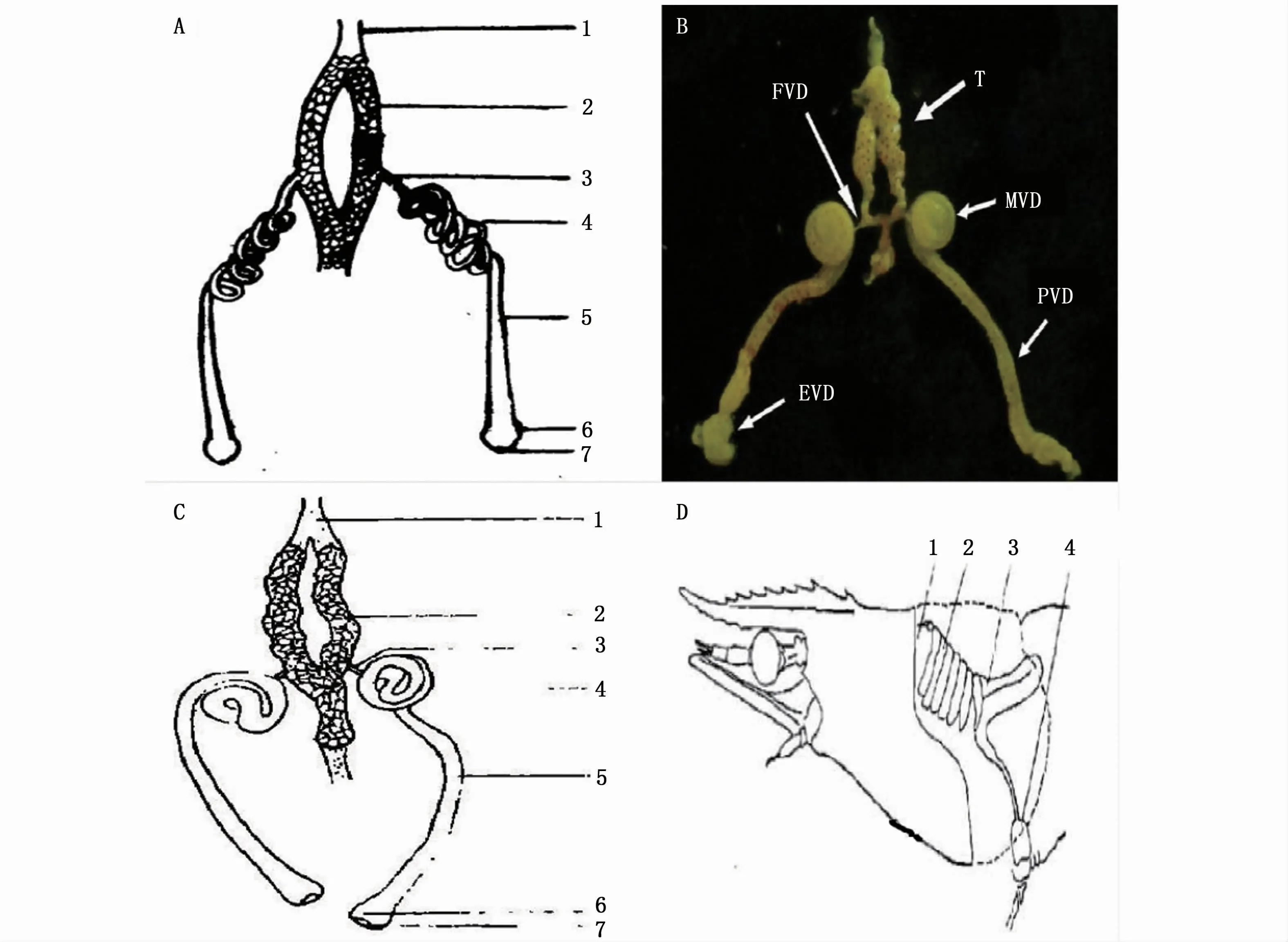
圖1 日本沼蝦[45](A)、脊尾白蝦[44](B)、秀麗白蝦[46](C)、中國明對蝦[47](D)雄性生殖系統Fig 1 Male reproductive system of M.nipponense[45](A),E.carinicauda[44](B),E.modestus[46](C)and Fenneropenaeus chinensis[47](D)
2.2.2 精子發生
長臂蝦類的精莢呈索狀,由中央的精子群、粘液團及包被在其外的精莢壁組成[48-49]。早期的精莢主要由精莢基質構成,由輸精管上皮分泌并逐漸積累到精子團外部;隨后輸精管上皮分泌形成精莢壁,最終產生完整精莢[50]。日本沼蝦、脊尾白蝦、秀麗白蝦的精莢形成過程見表5。

表5 3種長臂蝦類的精莢形成Tab.5 Spermatophore formation of three species of Palaemonidae
3 胚胎和幼體發育
3.1 胚胎發育過程
胚胎發育過程是繁殖生物學重要的研究內容,胚胎發育過程中,卵粒的大小、體積和含水量隨胚胎發育而增加。長臂蝦類胚胎發育過程基本一致,以雌雄蝦抱對受精產生的受精卵為起始,卵內僅1個細胞;卵裂期受精卵內的細胞開始分裂,形成分裂球;細胞經過第8次分裂數量增加到256個,進入囊胚期,此時細胞核全部移近受精卵表面,構成囊胚層;受精卵動物極細胞密集,植物極內陷形成原口,標志著原腸期的開始;胚胎卵黃含量減少,進入前無節幼體期,兩對觸角原基和一對大顎原基的形成是前無節幼體期的主要事件;后無節幼體期,形成兩對顎足原基和兩對小顎原基;進入前溞狀幼體期的胚胎,透明區內出現一對由復眼原基內色素細胞分泌的黑色顆粒狀物;復眼原基形成體積更大的橢圓形復眼,此時稱為溞狀幼體期或后溞狀幼體期,胚胎頭胸部13對附肢發育基本完成,隨后幼體破膜而出,胚胎期結束[51-54]。3種蝦類的胚胎發育的劃分方法見表6。

表6 胚胎發育分期Tab.6 Stages of embryonic development
目前根據胚胎發育的進程,長臂蝦類胚胎發育分期主要有7期法[52]、8期法[51,53]和12期法[54]。12期法對卵裂期的劃分過于詳細、工作量大且實用性較低;7期法以受精卵卵裂開始為胚胎發育第1期,這與事實不相符。與前兩者相比,8期法更加簡潔且實用,其第6期命名為“后無節幼體期”,與第5期“前無節幼體期”可以鮮明對比;第8期命名為“膜內溞狀幼體期”,相比“后溞狀幼體期”的命名更能概括這一時期的特征。
3.2 卵裂方式
幾種重要長臂蝦類的卵裂方式基本一致,但與對蝦科、螯蝦科蝦類(以下簡稱對蝦類、螯蝦類)的卵裂方式存在差異,3科蝦類的卵裂方式及區別見表7[51-56]。卵裂方式主要受到卵黃含量影響,3科蝦類的卵主要屬于中黃卵(日本沼蝦為多黃卵),但實際卵黃含量仍存在差異:對蝦類單次產卵量大,卵黃含量較少,分裂面可將卵完全分割開;長臂蝦類產卵量較少,卵黃含量較多,分裂面不能將卵完全分割開;螯蝦類產卵量極少,卵黃含量較多,卵裂以不完全卵裂為主。此外,相關文獻雖未明確指出脊尾白蝦的卵裂方式,但結合日本沼蝦和羅氏沼蝦卵裂方式的描述及筆者的觀察研究,認為脊尾白蝦卵裂方式應屬于完全卵裂和不完全卵裂之間的過渡類型。
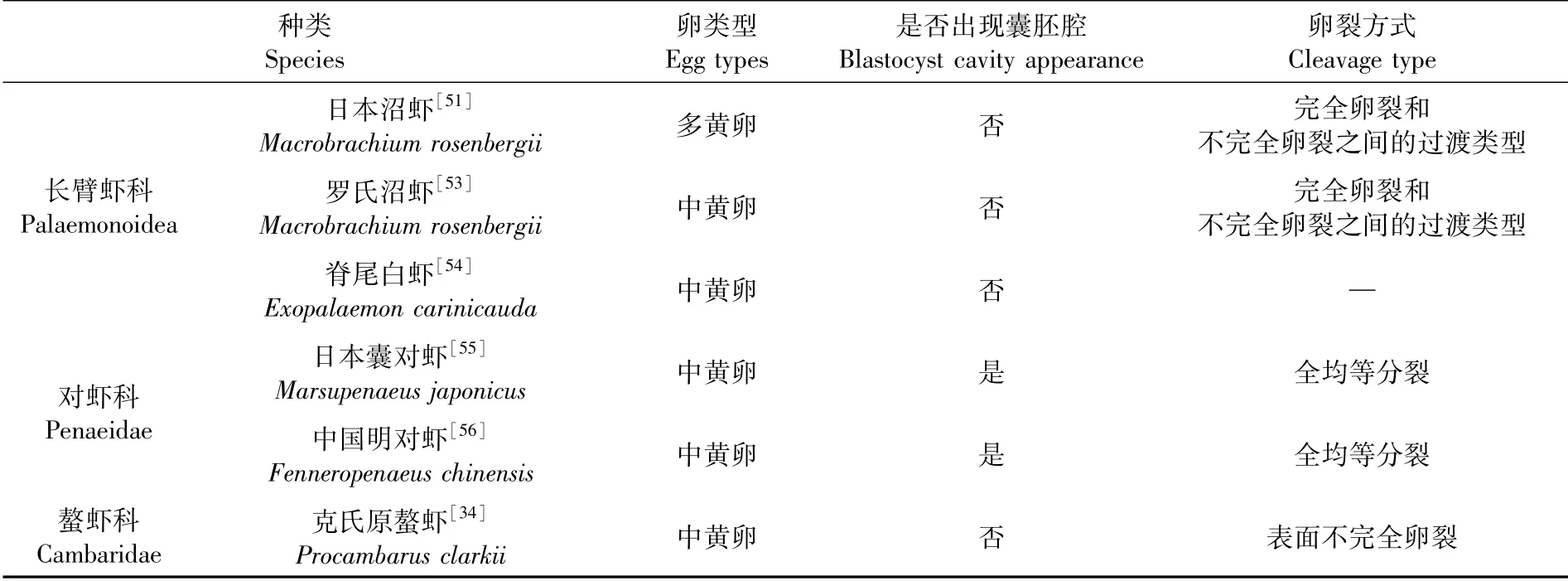
表7 長臂蝦類、對蝦類、螯蝦類的卵裂方式Tab.7 Cleavage patterns of Palaemonidae,Penaeidae and Cambaridae
3.3 幼體發育過程
不同種蝦類在胚胎內度過的時間長短不同,這導致幼體出膜形態以及后續發育進程呈現出不同的類型。對蝦類在無節幼體期前就破膜而出[57],長臂蝦類無節幼體期在胚胎中度過,出膜的幼體一般是溞狀幼體[58]。長臂蝦類剛出膜的溞狀幼體運動能力較弱,自然條件下,早期溞狀幼體為植食性,主要濾食各種藻類來獲取營養,人工養殖條件下會投喂鹵蟲無節幼體等為其提供營養[58]。蝦類的幼體發育為變態發育,不同時期的幼體之間形態特征差異較大,幼體的形態特征及生活習性與成蝦有很大不同[36]。日本沼蝦幼體發育存在中間蛻皮現象,從Z5(經過4次蛻皮后的溞狀幼體)開始,凡需蛻二次皮方可完成一期形態遞進的幼體,其第一次蛻皮后,第二觸角外鞭形態無遞進變化,幼體形態、大小變化不大,即為中間蛻皮狀態,則需再蛻皮一次[58];其他長臂蝦類中未見相關報道,因此日本沼蝦蛻皮次數通常高于其他長臂蝦類。羅氏沼蝦、日本沼蝦與脊尾白蝦的幼體發育過程見表8[58-62]。研究人員對脊尾白蝦的蛻皮次數和發育時間存在2種觀點[60-61],這主要是由于對進入仔蝦期的判定不同,一種觀點認為幼體需經過6次蛻皮才能發育完好進入仔蝦期[60];而另一種觀點則認為經5次蛻皮后,幼體即發育完好進入仔蝦期[61]。

表8 長臂蝦類幼體發育過程Tab.8 Larval development of Palaemonidae
3.4 環境因子對胚胎和幼體發育的影響
環境因子對長臂蝦類幼體孵化、發育、存活等有很大的影響[63-66]。溫度對于水生生物生長和存活有明顯影響,在適溫范圍內,溫度越高,水產動物胚胎與幼體發育速率越快,生長速度亦相應加快。當環境溫度超過動物自身的調節能力,例如,高于適溫范圍時,其代謝強度加大,能量無法積累或已經積累的能量被消耗;低于適溫范圍時,其代謝活動降低,都會導致生長速率減慢[63-66]。徐慈浩等[53]在對羅氏沼蝦的研究中指出,溫度的升高有利于加快胚胎發育的速度,但是過快的發育速率可能導致胚胎結構分化不夠完整,最終影響羅氏沼蝦幼體的孵化率和孵化質量。脊尾白蝦各期幼體在16~36℃范圍內,溫度越高發育速度越快,發育持續時間越短,幼體體長日增長量越大[67]。日本沼蝦[68]、羅氏沼蝦[69]的幼體發育也受溫度影響,這種影響主要是通過影響蝦類幼體有效積溫的累計來影響幼體發育的速率。
鹽度是影響長臂蝦類胚胎和幼體發育的另一重要環境因子。脊尾白蝦胚胎發育時間與發育時的鹽度成正相關[70]。通常蝦類胚胎和幼體對鹽度變化更為敏感[71],羅氏沼蝦幼體發育的適宜鹽度為10~14[69],成體則能在鹽度0~40的水中生存[72]。日本沼蝦幼體在鹽度為12時生長速度最快,成活率最高[73]。梁俊平等[74]認為,不同鹽度下(2~30)脊尾白蝦溞狀幼體的變態率和仔蝦存活率均無顯著差異,但鹽度15和20下仔蝦干質量顯著大于其他鹽度組,鹽度對仔蝦個體干質量影響顯著。
水體環境的pH是蝦類繁殖的重要環境因素之一,最為直觀的是影響幼體的存活率,過高或者過低的pH值都會降低幼體的存活率[69,75]。浦蘊惠[76]報道,脊尾白蝦胚胎適宜的pH在7.0,堿性環境對胚胎的影響更大,胚胎更適宜偏酸性的環境。pH值的變化能夠引起蝦體內酶活性的改變,從而影響蝦的各種生理活動,幼體處于身體迅速發育、各種組織逐漸形成與成熟的狀態,pH值變化的影響更加明顯。KAWAMURA等[77]研究了低pH水環境對羅氏沼蝦幼體觸覺的影響,指出較低pH值下,幼體觸覺剛毛中的甲殼素可能被降解,表面機械感受器被抑制。
此外,硫代硫酸鈉脫氯的自來水飼養羅氏沼蝦幼體會導致其存活時間縮短,發育遲緩[78];隨著飼料中蛋白含量的升高,斑節對蝦增重率先升高后下降,但不影響存活率[79],而飼料中添加β-胡蘿卜素能夠促進脊尾白蝦的生長,β-胡蘿卜素濃度為0.5 g·kg-1時脊尾白蝦的生長效果最佳[80]。
4 繁殖相關調控基因
長臂蝦類的繁殖過程受到多種基因調控,包括調控性腺發育、性別分化、胚胎發育、幼體發育等生理過程的基因,已見報道的長臂蝦類中發現的參與繁殖過程調控的基因見表9[82-87,90-107]。

表9 長臂蝦類繁殖相關調控基因Tab.9 Regulation genes related to reproduction of Palaemonidae
性腺發育相關基因的研究報道較多,傳統繁殖生物學觀點認為,甲殼綱動物的性別決定是由性染色體主導的,成熟的性腺能夠分泌各種性激素作用于身體中的其他組織,從而引起非生殖組織產生性別差異。最近的研究否定了這種觀點,研究人員發現中樞神經系統性別分化的出現早于性激素的分泌[81],這說明在發育初期,性別分化由基因單獨決定的,隨后激素開始分泌,性別分化才會逐漸受到激素的影響。目前已發現如卵巢特 異 基因[79]、叉頭盒蛋 白L2基 因[83]、vasa[84]等能夠影響性腺發育,參與蝦類發育的性別分化。
胚胎與幼體的生長速率以及存活率也是蝦類繁殖能力的重要反映。這一過程受多種基因調節,目前在長臂蝦科已見報道的EcR[85]、WNT4[86]、TRY[87]等都對胚胎發生、幼體生長或者這兩者的發育有著調控作用。幼體生長過程中,需要經歷多次蛻皮,蛻皮速率一定程度上會影響生長速率。無脊椎動物缺乏真正的抗體和特異性的免疫細胞,機體防御反應依靠非特異的固有免疫結構[88],因此,蝦類僅能通過被動的先天免疫抵御外界影響,幼體期發育經過多次生長蛻皮,蛻皮時幼體先天性免疫能力減弱,且幼蝦身體結構發育不成熟,對外界影響耐受能力較弱,免疫能力是幼體生長尤其蛻皮時期存活率的關鍵影響因素之一。一些與長臂蝦類免疫功能相關的基因可能也在幼體發育過程中發揮作用,如α2-巨球蛋白(alpha2-macrogloblin,α2M)、C-型凝集素(C-type lectins,CTL)[89]等,這些基因對幼體生長期間免疫功能的影響有待進一步研究。
5 總結與展望
長臂蝦科幾種重要經濟蝦類的繁殖期一般為5個月以上,繁殖盛期一般在5—8月份,一般有多次繁殖盛期。繁殖力方面,羅氏沼蝦的單次產卵量最高,而秀麗白蝦單次產卵量較少。大部分長臂蝦類自然群體年平均雌雄性比高于1,但波動較大。
長臂蝦類胚胎發育過程包括多種生理活動,將長臂蝦類的胚胎發育過程分為8個時期的分期方法較為合理。長臂蝦類孵出的溞狀幼體需發育至Ⅲ期溞狀幼體方具備運動捕食的能力,因此該階段需注意保證幼體的營養供應以及提供合適的開口餌料。胚胎和幼體發育過程受到溫度、鹽度、pH等多種環境因子的影響,提供適宜的環境條件能夠顯著提高胚胎孵化率、幼體存活率以及生長速率等。
多種基因參與調控長臂蝦類繁殖過程,其中一些基因或參與早期的性別分化,或參與性腺的發育。長臂蝦類雌雄個體的大小、生長速率存在明顯差異,通過研究性別分化基因,使得人工控制種群性別成為可能,單性化養殖可能是性別分化基因研究未來的發展方向。
目前養殖規模較大且經濟價值高的日本沼蝦、羅氏沼蝦和脊尾白蝦等已有較為全面的繁殖生物學研究。秀麗白蝦、葛氏長臂蝦、海南沼蝦、細螯沼蝦等有一定的產量,但主要依靠野外捕撈,且相關研究較少。繁殖相關基因與繁殖過程的速率與成功率密切相關,同時其研究也是性成熟、胚胎與幼體發育、生長蛻皮等過程的重要理論基礎。
