黃土丘陵區退耕草地群落優勢種葉片光合生理對氮磷添加的響應
2021-08-06 10:24:02賴帥彬簡春霞陳志飛周俊杰徐炳成
生態學報 2021年13期
賴帥彬,徐 升,簡春霞,陳志飛,周俊杰,楊 全,王 智,,徐炳成,,*
1 西北農林科技大學 黃土高原土壤侵蝕與旱地農業國家重點實驗室, 楊凌 712100 2 吉林農業科技學院, 吉林 132101 3 中國科學院水利部水土保持研究所, 楊凌 712100
近年來,大量礦物燃料燃燒以及農業中氮肥施用等致使大氣氮沉降遞增[1]。長期氮沉降在消除或緩解氮限制的同時,會造成陸地生態系統轉為磷限制或氮磷共同限制[1-3]。氮素是蛋白質、核酸、酶和葉綠素等重要組分,是限制植物光合作用的關鍵因子[4]。磷是光合過程中核酸、ATP和磷脂等化學物質的主要成分,也是土壤中最易被固定和轉化的元素,但即使向土壤中施加磷肥絕大部分會被有機物或金屬離子固定,轉化成緩效態和難效態,能為植物利用的有效磷非常少[5]。研究表明,氮限制下適宜添加氮能提高植物光能利用效率,促進植物生長和增加地上生物量[6],但當氮素超過某一閾值后,氮添加會轉變為抑制植物生長[6-7]。磷素能提高扁豆根瘤固氮菌活性,從而增加葉片氮磷含量和提高光合速率[8]。因此土壤氮磷供應狀況會對植物光合生理過程產生影響[9-10]。
不同植物由于其光合途徑、養分敏感性和分配策略差異等導致其對氮磷添加響應存在差異[11-13]。光合氮利用效率(PNUE)和光合磷利用效率(PPUE)是表征植物葉片養分利用及光合生理特征的重要參數[14]。一般來說,C4植物的PNUE和PPUE高于C3植物,主要是C4植物的CO2濃縮機制使得1,5-二磷酸核酮糖羧化酶/加氧酶(ribulose-1,5-bisphosphate carboxylase/oxygenase,Rubisco)周圍CO2達到飽和,使其在有效氮磷條件下擁有較高的凈光合速率[11];另一方面,PNUE和PPUE的差異受葉片氮和磷分配影響[11]。氮素充足時,相比C3植物,C4植物分配少量N到光合酶中即可獲得最大光合速率,將其余氮素分配到根莖部以獲得較高生長速率[12]。相比禾本科植物,豆科植物將更多葉氮分配于光合系統,但禾本科植物在光合系統內部的氮分配機制更為有效[15]。由此可見,環境變化過程中,物種屬性在決定其光合氮、磷分配及利用效率上發揮著重要作用。
退耕草地在保持水土、涵養水源、調節氣候及防風固沙等方面具有重要生態保護作用[16]。對土壤養分貧瘠的退耕草地而言,合理均衡的氮磷添加可提升土壤養分含量,改變植株葉片氮磷含量,提高葉片光合能力,促進植被生長,加快草地恢復進程[7,9]。研究表明,內蒙古溫帶草原在適宜氮添加后羊草(Leymuschinensis)光合能力顯著增強,地上生物量顯著增加[7];適宜氮添加后,青海高寒草甸區禾本科植物凈光合速率顯著提升,生長促進明顯[9],而隨施氮量增加豆科植物葉片氮含量未發生變化但生物量逐漸下降[10]。因此,氮磷添加會對植物群落組成及各植物光合生理過程產生影響。
優勢種在群落中占據著重要生態位,對群落結構和功能植被生態系統穩定起著至關重要的作用,旱區植被對環境條件變化的響應,主要取決于其優勢種的響應和適應能力[17-18]。因此,明確主要優勢種對氮磷添加的生理生態響應與變化特征,對揭示氮磷添加對退耕草地植被生產力和物種多樣性的影響具有一定意義[18]。近年來,關于氮磷添加對退耕草地的效應研究主要集中在植物生長方面,而關于優勢種在氮磷添加下光合生理生態研究較少[13,19]。白羊草(Bothriochloaischaemum)、長芒草(Stipabungeana)和達烏里胡枝子(Lespedezadavurica)為陜北黃土丘陵區草地群落中常見種,三者在該區退耕草地群落中均具有較廣的分布幅度和占比[20],其中白羊草和長芒草分別為禾本科多年生C4和C3植物,達烏里胡枝子為豆科多年生C3半灌木。因此,本研究以這3種常見優勢種為對象,通過測定分析3個優勢種在不同配比氮磷添加下的光合生理參數、葉綠素熒光參數以及葉片氮磷含量等,探討不同優勢種光合生理對氮磷添加的響應特征與差異,以期為黃土丘陵區退耕草地恢復過程中的施肥管理與調控提供科學依據。
1 材料與方法
1.1 研究區概況
試驗區位于陜西省延安市安塞區紙坊溝小流域,地處黃土丘陵溝壑區腹地,屬暖溫帶半干旱大陸性季風氣候。地理位置為東經109°13′46″—109°16′03″,北緯36°42′42″—36°46′28″,海拔為1041.5—1425.7 m。年均降雨量為507.2 mm(1951—2018),且季節分配不均,從全年降水量來看,4—10月占85%—95%,7—9月占60%—80%。年均氣溫8.8°C,輻射總量為493 kJ/cm2。土壤類型主要為黃綿土,植被區劃上屬暖溫帶半干旱森林草原區,處于落葉闊葉林區和草原區的過渡帶。天然植被中以旱生草本植物群落為主,包括白羊草、長芒草、達烏里胡枝子、鐵桿蒿(Artemisiasacrorum)、茭蒿(Artemisiagiraldii)等;有少量天然喬、灌木群落分布,主要由遼東櫟(Quercuswutaishanica)、黃刺玫(Rosaxanthina)等組成。
1.2 試驗設計
在紙坊溝流域,選擇以白羊草、長芒草和達烏里胡枝子為優勢種的退耕草地群落為試驗樣地,坡度為7°,坡向為東偏西46°。采用裂區試驗設計,分別設置3個4 m×4 m 主區,區間由1 m寬未施肥緩沖區隔開,每個主區再分為4個2 m×2 m 的副區。主區為施氮處理,副區為施磷處理。根據黃土丘陵區年均氮沉降量(21.76 kg N hm-2a-1)[2],設置2倍和4倍的年均模擬氮沉降水平;參照劉海威等[21]的施磷標準(32.55 kg P2O5hm-2a-1),設置1倍和2倍的年施磷量,3個N添加處理分別為N0(0 kg N hm-2a-1)、N50(50 kg N hm-2a-1)和N100(100 kg N hm-2a-1),3個P添加處理分別為P0(0 kg P2O5hm-2a-1)、P40(40 kg P2O5hm-2a-1)和P80(80 kg P2O5hm-2a-1)。氮肥為硝酸銨鈣[5Ca(NO3)2NH4NO310H2O],磷肥為重過磷酸鈣[Ca(H2PO4)2H2O]。于2019年2月將上年地表所有枯枝落葉移除,在雨季來臨前的6月6日,將提前稱好的顆粒狀氮肥和磷肥均勻撒入對應小區。
1.3 測定指標及方法
1.3.1光合生理特征
采用CIRAS- 2光合儀(美國PP System 公司),在無風晴朗天氣里連續3天依次測定白羊草(抽穗期)、長芒草(開花期)和達烏里胡枝子(開花前期)的光合生理特征(每天完成一個優勢種的光合生理指標測定)。本次測定于2019年7月19—21日09:00—11:30進行。每種植物測定時選取生長狀況一致的植株,相同葉位且完全展開的成熟葉片,3個生物學重復。測定項目包括:凈光合速率(Pn,μmol m-2s-1)、蒸騰速率(Tr,mmol m-2s-1)、氣孔導度(Gs,mmol m-2s-1)、胞間CO2濃度(Ci,μmol/m)等。測定時使用自帶紅藍光源(光強設置為1200 μmol m-2s-1),葉溫設為25℃,相對濕度為60%,大氣CO2濃度為380 μmol/mol。3種植物單葉面積均小于光合儀葉室面積,因此在測定時,使用佳能D7500相機對葉室內測定葉片拍照,后期在室內用Adobe Photoshop將圖片處理后,再用Image-J計算出實際光合葉面積,將求得葉面積值輸入光合軟件(CIRAS- 2 remote control software)進行換算。葉片瞬時水分利用效率(WUEi,μmol/mmol)=Pn/Tr[22];氣孔限制值(Ls)=1-Ci/Ca[23]。
1.3.2葉綠素熒光參數
采用連續激發式植物效率分析儀Handy PEA(英國 Hansatech 公司),在測定光合氣體交換參數后,選擇同一植株測定葉綠素熒光參數。測定前用暗適應夾夾住葉片暗適應30min(夾葉片時避開葉脈,當葉片太小時,將兩葉片并排以填滿暗適應夾圓形孔隙)。測定指標包括葉綠素初始熒光(Fo)、最大熒光(Fm)、PSⅡ最大光化學效率(Fv/Fm)和PSⅡ潛在活性(Fv/Fo)[24],每處理3次重復。
1.3.3葉片氮、磷含量
在葉片光合和熒光參數測定結束后,迅速采集同一株植物相同葉位5—6枚葉片裝于自封袋中,帶回實驗室后將葉片平鋪于A4紙上,用佳能750D相機垂直拍照并用Image-J計算葉片面積后,將葉片置于105℃烘箱殺青15min,75℃烘至恒重(0.0001g)。葉片采用MM400高通量組織研磨儀(Retsch,德國)研磨,經H2SO4-HClO4消煮后使用全自動凱氏定氮儀(FOSS- 8400,丹麥)測定葉片氮含量(Nmass)。采用鉬藍比色法(UV- 2600 紫外可見分光光度計,日本)測定葉片磷含量(Pmass)。葉片單位面積氮含量[Narea,g(N) m-2]和單位面積磷含量[Parea,g(P) m-2]分別為單位質量葉片氮含量(Nmass,g/kg)和葉片磷含量(Pmass,g/kg)與比葉面積(SLA)的比值,其中SLA為葉面積與葉片干重之比。光合氮利用效率[PNUE,μmol(CO2) g(N)-1s-1]和光合磷利用效率[PPUE,μmol(CO2) g(P)-1s-1]分別為凈光合速率(Pn)與Narea和Parea的比值[11]。計算公式分別為:
Narea=Nmass/SLA
(1)
Parea=Pmass/SLA
(2)
PNUE=Pn/Narea
(3)
PPUE=Pn/Parea
(4)
1.4 數據分析
利用單因素方差分析(One-way ANOVA)分別檢驗各指標(光合氣體交換參數、熒光參數、光合氮和磷利用效率)在不同氮磷添加處理下的差異顯著性(P=0.05)。利用雙因素方差分析(Two-way ANOVA)分別檢驗氮磷添加的交互作用對各優勢種指標影響的差異顯著性。采用線性模型擬合凈光合速率與PSⅡ最大光化學效率和PSⅡ潛在活性以及凈光合速率與葉片氮含量和葉片磷含量間的關系。采用SPSS 20.0處理數據,OriginPro 2016繪圖。
2 結果與分析
2.1 光合生理特征
由圖1可知,單獨氮添加下,白羊草和達烏里胡枝子的Pn相比未施肥顯著升高(P<0.05)。白羊草的Pn在N50和N100處理間差異不顯著,達烏里胡枝子的Pn在N100下顯著高于N50處理(P<0.05),長芒草的Pn僅在N100下顯著高于未施肥處理。單獨磷添加下,僅達烏里胡枝子的Pn較未施肥顯著增加,且P40和P80處理間差異顯著。在N50和N100處理下,磷添加對白羊草和長芒草的Pn無顯著影響。施磷顯著提高了達烏里胡枝子的Pn,N50處理下P40和P80處理間差異顯著,而N100下P40和P80處理間差異不顯著。氮磷添加對白羊草的Pn交互作用顯著,而對長芒草和達烏里胡枝子無顯著影響。
單獨氮添加下,達烏里胡枝子的Tr相比于未施肥顯著增加,且N100顯著高于N50處理(P<0.05);白羊草和長芒草的Tr僅在N100下顯著高于未施肥處理(P<0.05),分別為后者的2.13和1.54倍。單獨磷添加下,白羊草的Tr無顯著變化,長芒草和達烏里胡枝子的Tr在P80和P40處理下均顯著降低(P<0.05)。N50下,白羊草、長芒草和達烏里胡枝子的Tr均隨施磷量增加顯著增加,其中白羊草和長芒草的Tr在P40和P80處理間差異不顯著,而達烏里胡枝子的Tr在各處理間差異顯著(P<0.05);N100下,僅達烏里胡枝子的Tr施磷后顯著增加,且P40和P80處理間差異顯著(P<0.05)。氮磷添加對3個優勢種的Tr交互作用均顯著(P<0.05)(圖1)。
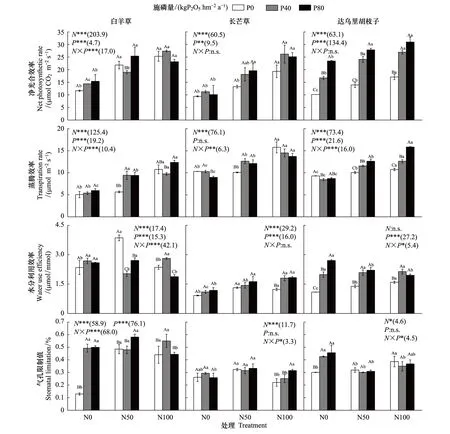
圖1 不同氮磷添加處理下三種優勢植物葉片凈光合速率、蒸騰速率、水分利用效率和氣孔限制值Fig.1 Leaf net photosynthetic rate (Pn), transpiration rate (Tr), water use efficiency (WUE) and stomatal limiting value (Ls) of the three dominant species under different N and P addition treatmentsN0表示0 kg N hm-2 a-1;N50表示50 kg N hm-2 a-1;N100表示100 kg N hm-2 a-1; 柱上不同大寫字母表示同一施氮處理下施磷處理間差異顯著,不同小寫字母表示同一施磷處理下施氮處理間差異顯著(P<0.05);括弧內數字為不同氮磷處理下的F值;*,**和***分別表示P≤0.05,P≤0.01和P≤0.001
單獨氮添加下,三個優勢種的WUEi相比未施肥處理顯著升高(P<0.05)。白羊草和長芒草的WUEi在N50和N100處理間差異不顯著,達烏里胡枝子的WUEi在N100處理下顯著高于N50處理(P<0.05)。單獨磷添加下,僅達烏里胡枝子的WUEi較未施肥處理顯著提升,且P80和P40處理間差異顯著(P<0.05)。N50處理下,磷添加對長芒草的WUEi無顯著影響,顯著降低了白羊草的WUEi,顯著提升了達烏里胡枝子WUEi。N100處理下,磷添加顯著提升了長芒草和達烏里胡枝子的WUEi,但P40和P80處理間差異不顯著;白羊草的WUEi在P40處理下顯著提升,但P80處理下顯著降低。氮磷添加對白羊草和達烏里胡枝子的WUEi交互作用顯著,對長芒草的WUEi無顯著影響(圖1)。
單獨氮添加下,白羊草的Ls相比未施肥處理顯著增加(P<0.05)。白羊草的Ls在N50和N100處理間差異不顯著,長芒草的Ls在N50處理下顯著高于N100處理(P<0.05),達烏里胡枝子的Ls僅在N100處理下顯著增加。單獨磷添加下,僅白羊草的達烏里胡枝子的Ls相比未施肥顯著增加(P<0.05),且均在N50和N100處理間差異不顯著。N50處理下,磷添加對長芒草和達烏里胡枝子的Ls無顯著影響,白羊草的Ls僅在P80處理下顯著增加;N100處理下,磷添加對達烏里胡枝子的Ls無顯著影響,白羊草的Ls僅在P40處理下顯著增加,長芒草的Ls僅在P80處理下顯著增加(P<0.05)。氮磷添加對三個優勢種的Ls交互作用顯著(圖1)。三個優勢種的Pn與Nmass均呈顯著正相關關系,僅長芒草和達烏里胡枝子的Pn與Pmass呈顯著正相關關系(圖2)。

圖2 不同氮磷添加處理下三種植物葉片凈光合速率與葉片氮磷含量關系Fig.2 Relationships between net photosynthetic rate with Leaf N and P contents of the three dominant species
2.2 葉綠素熒光特征
單獨氮添加下,白羊草和達烏里胡枝子的Fo相比未施肥處理顯著降低(P<0.05),長芒草的Fo相比未施肥處理顯著增加(P<0.05);白羊草的Fv/Fm和Fv/Fo僅N100下顯著高于未施肥處理(P<0.05),而長芒草和達烏里胡枝子的Fv/Fm和Fv/Fo在各處理間差異不顯著。單獨磷添加下,白羊草和達烏里胡枝子的Fo相比未施肥處理顯著降低(P<0.05),其中白羊草的Fo在P80處理下顯著低于P40處理,達烏里胡枝子的Fo在P40和P80處理間差異不顯著(P<0.05);白羊草的Fv/Fo相比未施肥處理顯著增加,且P40和P80處理間差異不顯著,而Fv/Fm僅在P40處理下顯著高于未施肥處理(圖3)。
N50下,磷添加顯著降低了白羊草的Fo,顯著增加了達烏里胡枝子的Fv/Fo;白羊草的Fo在P40和P80處理間差異顯著,長芒草和達烏里胡枝子的Fo僅在P80處理下顯著降低;白羊草的Fv/Fm和Fv/Fo僅在P40處理下顯著增加,長芒草的Fv/Fm僅在P80處理下顯著增加,達烏里胡枝子的Fv/Fo在P40和P80處理間差異不顯著(P<0.05)。N100下,磷添加顯著降低了白羊草和達烏里胡枝子的Fo,顯著增加了長芒草和達烏里胡枝子的Fv/Fm和Fv/Fo;白羊草和達烏里胡枝子的Fo在P40和P80處理間差異不顯著,長芒草的Fo僅在P80處理下顯著增加;在P40和P80處理間,長芒草和白羊草的Fv/Fm和Fv/Fo和達烏里胡枝子的Fv/Fm差異不顯著,達烏里胡枝子的Fv/Fo差異顯著。氮磷添加僅對三個優勢種的Fo和白羊草的Fv/Fm、達烏里胡枝子的Fv/Fo交互作用顯著(圖3)。
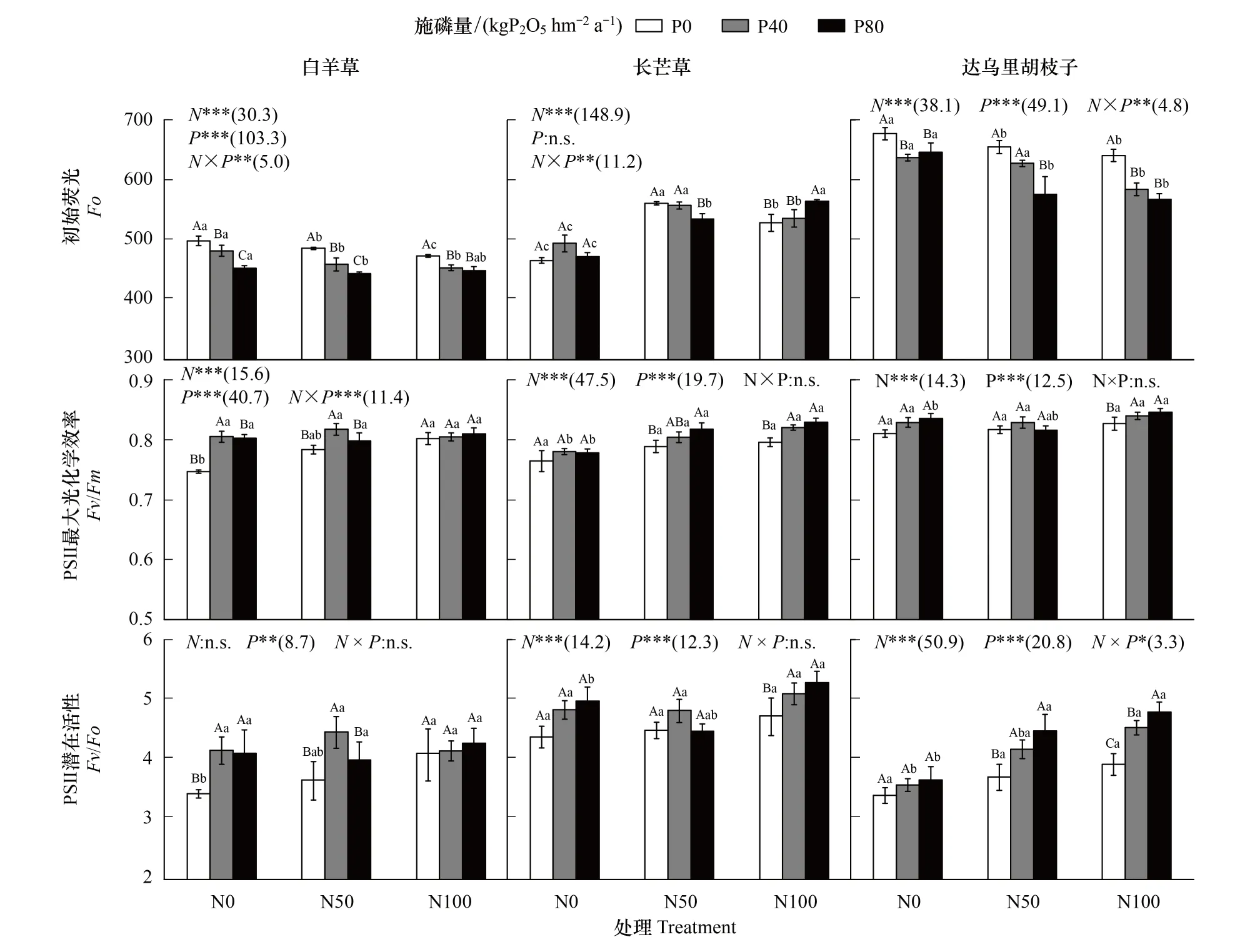
圖3 不同氮磷添加處理下三種優勢植物初始熒光、PSⅡ最大光化學效率和PSⅡ潛在活性Fig.3 Initial fluorescence, PSⅡ maximum photochemical efficiency and PSⅡ potential activity of the three dominant species under different N and P addition treatments
相關性分析表明,三個優勢種的Pn與Fv/Fm呈顯著正相關(白羊草:R2=0.31,P<0.01;長芒草:R2=0.35,P<0.01;達烏里胡枝子:R2=0.63,P<0.01),其中長芒草和達烏里胡枝子的Pn—Fv/Fm斜率明顯高于白羊草。三個優勢種僅長芒草和達烏里胡枝子的Pn與Fv/Fo呈顯著正相關(長芒草:R2=0.33,P<0.01;達烏里胡枝子:R2=0.70,P<0.01),其中達烏里胡枝子的Pn—Fv/Fo斜率高于長芒草(圖4)。
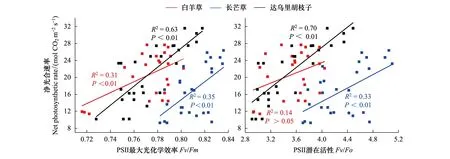
圖4 不同氮磷添加處理下三種植物葉片凈光合速率與PSⅡ最大光化學效率和PSⅡ潛在活性關系Fig.4 Relationships between maximum net photosynthetic rate with PSⅡ maximum photochemical efficiency and PSⅡ potential activity of the three dominant speciesFv/Fm:光系統Ⅱ最大光化學效率 PSⅡ maximum photochemical efficiency;Fv/Fo:光系統Ⅱ潛在光化學活性 PSⅡ potential activity
2.3 PNUE和PPUE
單獨氮添加下,白羊草和達烏里胡枝子的PNUE、白羊草和長芒草的PPUE相比未施肥處理顯著提升,長芒草的PNUE相比未施肥處理顯著降低(P<0.05)。白羊草在N100下的PNUE顯著高于N50(P<0.05),長芒草和達烏里胡枝子的PNUE在N50和N100間無差異;白羊草和長芒草的PPUE在N50和N100間差異顯著,達烏里胡枝子的PPUE僅在N50下顯著增加。單獨磷添加下,長芒草的PNUE和白羊草的PPUE相比未施肥處理顯著降低(P<0.05)。長芒草的PNUE僅在P80處理下顯著降低,而達烏里胡枝子的PNUE僅在P80處理顯著增加;白羊草的PPUE在P40和P80間無差異,長芒草的PPUE在P40下顯著增加而在P80下顯著降低,達烏里胡枝子的PPUE僅在P80處理顯著增加(圖5)。
N50下,磷添加后長芒草的PNUE顯著增加,而白羊草的PPUE顯著降低。長芒草的PNUE在P40和P80間無差異,白羊草和達烏里胡枝子的PNUE僅在P80下顯著增加;白羊草的PPUE在P40和P80間無差異,達烏里胡枝子的PPUE僅在P40下顯著增加。N100下,磷添加后僅達烏里胡枝子的PNUE顯著增加。達烏里胡枝子的PNUE在P80下顯著高于P40,長芒草的PNUE僅在P40處理下顯著增加;白羊草的PPUE僅在P40下顯著增加,長芒草的PPUE僅在P80下顯著降低。氮磷添加對三優勢種的PNUE和白羊草的PPUE交互作用顯著,對長芒草和達烏里胡枝子的PPUE無影響(圖5)。
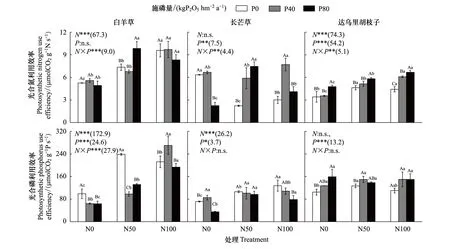
圖5 不同氮磷添加處理下三個優勢植物葉片光合氮和光合磷利用效率 Fig.5 Leaf photosynthetic N and P utilization efficiency of the three dominant species under different N and P addition treatments
3 討論
氣體交換參數反映植物光合速率變化的內在特征[6],其中Pn是表征植物光合作用大小的直接指標,是植物形成生物量的關鍵。植物種生理屬性影響Pn對氮磷添加的響應[6,25-26]。本研究中,單獨的氮添加下,白羊草和達烏里胡枝子在N50和N100處理下的Pn均顯著高于未施肥對照,且白羊草的Pn在兩個氮添加處理間差異不顯著,而長芒草的Pn僅在N100處理下顯著高于未施肥對照,表明禾本科C4植物白羊草在較低氮添加下即可達到較高的Pn,這可能是白羊草的CO2富集機制使其具有較高的Rubisco活性所致[12]。N50和N100處理下,達烏里胡枝子的Pn顯著增加且各處理間差異顯著,這可能是氮添加后改變了其光合氮與非光合氮分配比例[25](圖5)。單獨磷添加下,僅達烏里胡枝子的Pn顯著增加,說明具有固氮能力的達烏里胡枝子對磷素添加更為敏感[10]。氮添加提高了禾本科和豆科植物的Pn,磷添加僅顯著提升了豆科植物的Pn,這與兩種功能型物種對土壤氮磷有效性變化的敏感度有關[7,10]。不同物種的Pn與葉片氮磷含量之間的相關性存在差異[27-28]。本研究結果表明,白羊草的Pn與Nmass呈正相關關系但與Pmass不相關,達烏里胡枝子和長芒草Pn與Nmass和Pmass均呈顯著正相關關系(圖2),說明單獨的氮或磷添加,通過提高優勢種葉片氮或葉片磷含量增加了Pn[6-7]。白羊草的Pn與Pmass無相關性,這可能是由于作為C4植物白羊草具有更高的Rubisco酶效率因而擁有較高的PPUE,分配較少的磷于光合組分中即可擁有較大的Pn[11,29]。相比于長芒草,達烏里胡枝子的Pn與Pmass的相關性更強,這可能是由于磷素能夠促進達烏里胡枝子結瘤和提高其根瘤固氮酶活性,增加了葉片氮和光合能力[8]。
合理的氮磷添加能調節植物光合和蒸騰速率,進而影響植物WUEi[6,25]。本研究中,單獨氮添加下長芒草和達烏里胡枝子的Tr相比于Pn增幅較小,引起WUEi顯著提升;白羊草WUEi僅在N50處理下顯著提升,N100處理下白羊草WUEi降低源于蒸騰速率增幅較大。氮添加下,白羊草較高的WUEi是由于其相對較高的Pn與相對較低的Tr所致,其較低Tr與C4植物較高的葉肉阻力和氣孔阻力有關[30]。單獨磷添加下達烏里胡枝子WUEi的顯著提升歸因于Pn的增幅較高(圖1)。氮添加下,三個優勢種的Ls增加;磷添加下僅白羊草和達烏里胡枝子Ls顯著增加;氮磷添加下三個優勢種Pn和Tr無統一變化規律,表明氮磷添加通過影響蒸騰速率和氣孔限制因素改變植物光合能力[31-32]。
氮磷添加在影響植物光合作用的同時,對植物熒光特征產生影響[6]。研究表明,適宜氮磷添加常伴隨PSⅡ反應中心光化學效率和潛在活性的提高,即Fo降低和Fv/Fm與Fv/Fo的增加[6]。本研究中,單獨氮或磷添加下,僅白羊草的Fo顯著降低而Fv/Fm和Fv/Fo值均顯著增加,長芒草和達烏里胡枝子的Fv/Fm和Fv/Fo雖增加但均不顯著。說明C4植物白羊草的PSⅡ反應中心對氮磷添加的敏感性高于C3植物長芒草和達烏里胡枝子,這可能是由于C3和C4植物類囊體膜的光化學和非光化學過程存在差異[33-34]。白羊草在P40處理下的Fv/Fm和Fv/Fo與P0的差異顯著,但與P80差異不顯著,說明施磷過高可能對白羊草光能轉化和傳遞能力產生抑制作用[27]。Pn與Fv/Fm和Fv/Fo的擬合結果表明(圖3),適宜氮或磷添加增強了各優勢種PSⅡ反應中心光化學效率和潛在活性,提高了光合能力[6]。本研究中,氮磷共同添加下三個優勢種Pn、PSⅡ活性及長芒草的WUEi高于單獨氮或磷添加,雙因素分析也顯示氮磷共同添加對光合生理特征參數有交互作用(圖1),說明氮磷共同添加在提高三種植物光合生理特征參數方面優于單獨氮或磷添加[35-37]。
PNUE和PPUE是表征植物葉片生理特征和氮磷利用及分配的重要參數[14-15]。合理氮磷添加可優化氮磷分配能力,從而提高植物對環境的適應性[6,25]。本研究中,N50和N100處理下白羊草和達烏里胡枝子的PNUE和PPUE增加,且白羊草的PNUE和PPUE在兩個氮添加處理下分別為達烏里胡枝子的1.58、2.17和1.88、1.92倍,說明適當氮添加提高了兩個優勢種光合氮利用效率[38],相比達烏里胡枝子,白羊草具有更高效的氮磷分配機制[15]。N50和N100處理下長芒草的PNUE顯著降低,是由于其Pn相比于Narea增幅較小,說明禾本科C3和C4植物對氮磷分配策略存在差異,即白羊草分配更多氮于植物光合系統中,而長芒草則將更多的氮分配于光合系統以外部分(如細胞壁)[9,15]。磷添加下僅達烏里胡枝子的PNUE和PPUE在P80處理下相比于未施肥處理顯著增加,這歸因于磷添加后達烏里胡枝子的Pn顯著增加[15]。氮或磷添加下各優勢種PNUE和PPUE差異表明,白羊草和達烏里胡枝子將更多氮素分配到光合系統內部,而長芒草將更多氮素分配于光合系統以外部分,這與物種對資源利用策略有關[15,32]。
氮磷配施下,N50處理下,隨著施磷量增加三個優勢種PNUE逐漸增加,且兩個禾本科植物增加幅度大于豆科植物達烏里胡枝子,其中白羊草和長芒草的PNUE在N50P80處理下已達最大值;N100處理下,三個物種的光合能力相比于N50處理下未顯著增加,且白羊草和長芒草的PNUE在P80處理下出現下降(圖5)。從光合能力和養分利用效率來看,N50P80是提高優勢種光合能力和養分利用效率的最佳組合。氮磷施肥后植物光合氮分配策略的改變與三個優勢種對氮磷敏感性差異有關[15],這種差異對增強草地群落適應外部環境條件變化的穩定性具有重要作用。從群落多樣性和穩定性方面考慮,氮磷共同添加更有利于退耕草地植被恢復。
4 結論
氮或磷添加下,3個優勢種通過增加葉片氮或磷含量,活化了氣孔調節能力和PSⅡ活性,提高了光合能力,其中白羊草的光合氣體交換參數和葉綠素熒光參數對氮添加的敏感程度高于長芒草和達烏里胡枝子,達烏里胡枝子光合氣體交換參數對磷添加的敏感程度高于其余兩個優勢種。葉片氮含量增加是白羊草凈光合速率上升的主要原因,而長芒草和達烏里胡枝子的凈光合速率的提升則由葉片氮和磷含量增加所致。適當氮添加顯著提高了白羊草和達烏里胡枝子的光合氮利用效率以及三個優勢種的光合磷利用效率,三種植物光合生理特征對氮磷添加的響應與物種屬性和施肥水平有關。N50P80是提高優勢種光合能力和養分利用效率的最佳組合。上述結果表明,適當氮磷添加有利于提高半干旱區草地群落優勢種的光合能力,這是施肥提高草地生物量的生理基礎;草地群落優勢種光合生理特征對氮磷添加響應差異是其對環境變化不同步性的生理表現,不同優勢種在光合生理和養分利用方面的差異性表現對退耕草地恢復過程中維持群落多樣性和穩定性具有重要意義。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
趣味(語文)(2020年3期)2020-07-27 01:42:46
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
