水稻苗期根部性狀的遺傳分析和最長根長QTL qLRL4的精細定位
2021-08-05 11:29:28丁仕林劉朝雷阮班普姜洪真董國軍胡光蓮郭龍彪高振宇
作物學報 2021年10期
關鍵詞:水稻
田 彪 丁仕林 劉朝雷 阮班普 姜洪真 郭 銳 董國軍 胡光蓮 郭龍彪 錢 前 高振宇
水稻苗期根部性狀的遺傳分析和最長根長QTL的精細定位
田 彪 丁仕林 劉朝雷 阮班普 姜洪真 郭 銳 董國軍 胡光蓮 郭龍彪 錢 前 高振宇*
中國水稻研究所, 浙江杭州 310006
為了解析水培苗期根系相關性狀的遺傳調控, 以秈稻9311和粳稻日本晴(Nipponbare, NPB)為親本的148個株系構成的重組自交系群體為材料, 對水稻幼苗根系相關性狀開展QTL分析。在2次重復中共檢測到26個控制最長根長、總根系長、根表面積、根體積和根直徑的QTL, 分布在水稻第1、2、4、7、9、10、11號共7條染色體上, 發現了水稻第2、4、7和10號染色體上的4個QTL簇, 包括第4號染色體上控制最長根長的QTL。為了精細定位該QTL, 我們構建了以9311為背景、插入缺失標記IND4-1和IND4-4間來自NPB的近等系NIL-。利用NIL-和9311構建的F2群體, 最終將精細定位在標記IND4-1和IND4-3之間約68.23 kb的區間內并預測了候選基因。此根長QTL的精細定位將有助于水稻根長遺傳機理的研究, 為探究水稻根系形態建成的分子機制奠定了基礎。
水稻; 根部性狀; 最長根長;; 精細定位
水稻是人類最主要的糧食作物之一[1], 而根系作為水稻主要的器官, 具有固定水分、吸收養分、合成與分泌激素、酶和有機酸等功能[2]。雖然根系研究與其他性狀相比相對薄弱[3], 但是植物學家對根系相關性狀已開展了大量遺傳研究。徐吉臣等[4]以窄葉青8號和京系17的F1代花培加倍的127個雙單倍體(double haploid, DH)群體為材料, 測定了營養液培養條件下親本及DH株系的最大根長、根干重和根莖干重比, 檢測到4個最大根長QTL, 分別位于第2、4、9、10號染色體; 檢測到水稻第2號染色體的1個根干重QTL; 根莖干重比共檢測到3個QTL, 分別位于第3、5、6號染色體上。滕勝等[5]和胡興明等[6]也用相同群體進行了根部性狀的QTL分析。前者在抽穗期對根系活力進行QTL分析, 在第4號染色體的分子標記RG449和RG809間檢測到1個QTL ()。后者對水稻苗期發根力進行了QTL和上位性分析, 在水稻第3號染色體的C63~CT125之間檢測到1個發根力主效QTL; 同時檢測到影響水稻發根力的5對上位性效應基因座, 分別位于第2、3、5、6、7、12號染色體上, 其中影響根長和根數的上位性效應各有2對QTL, 有一對QTL同時影響根長和根數。迄今, 國內外科學家已定位了多個水稻根長相關QTL。Mitsuhiro等[7]用粳稻品種Koshihikari和秈稻品種Kasalath雜交和回交得到的38個染色體片段替代系(CSSL)檢測生長在5mmmol L–1或500mmmol L–1NH4+中的水稻幼苗根長QTL, 共檢測到8個QTL, 分別位于第1、2、4、6、8、11和12號染色體, 并將6號染色體的根長主效QTL精細定位在長臂337 kb區間內。王汝慈等[8]利用協青早B與中恢9308雜交構建的RIL群體在水稻第4號染色體長臂的分子標記RM307與RM1205之間定位到一QTL簇, 控制不同時期的相對根長、相對根冠比、相對根干質量等性狀, 其中包括控制水稻根長的主效QTL——。Obara等[9]以水稻WAB56-104和NERICA7品系的雜交F2群體為材料, 在高NH4+環境下于水稻第1號染色體檢測到2個水稻根長相關QTL, 分別命名為和, 其中的定位區間進一步縮小至0.7 Mb。Kitomi等[10]為了鑒定與水稻最大根長相關的基因組區間, 用來自IR64和KinandangPatong間雜交回交得到的26個CSSL進行QTL分析, 得到2個最大根長QTL: 2號染色體的和6號染色體的。將精細定位在SSR標記RM5651和RM6107之間的1.7 Mb區間, 將定位在SSR標記RM20495和RM3430-1之間884 kb的區間。到目前為止, 科學家已成功克隆了一些控制水稻根系性狀的基因[11-12], 如[13]和[14]基因調控水稻根長,[15]基因影響冠狀根的發生和不定根的形成,[16]基因影響不定根的形成,[17]基因促進冠根的起始和伸長,[18]和[19]基因影響不定根的的數目和長度,[20]基因抑制初生根的發育,[21]基因控制側根發生。為了深入了解水稻根系性狀的遺傳基礎, 我們以9311和日本晴(Nipponbare, NPB)構建的重組自交系(recombinant inbred lines, RIL)群體為材料, 利用水培技術和數量遺傳學手段對水稻根系性狀相關QTL進行了分析和鑒定。
1 材料與方法
1.1 試驗材料
研究采用秈稻品種9311和粳稻品種NPB雜交獲得的148個株系構成的RIL群體。我們選擇了區域具有NPB基因型的RIL與親本9311回交, 并在插入缺失(InDel)標記IND4-1和IND4-4之間選擇目標QTL區域攜帶NPB純合等位基因的近等基因系(near isogenic lines, NIL) NIL-。
1.2 試驗方法
在黑暗條件下, 水稻種子于37°C去離子水中浸泡2 d, 然后轉移至漂浮在去離子水上的網格, 培養3 d。將幼苗培養在正常濃度的Kimura B營養液中(pH 5.4), 每3 d更換一次營養液。然后, 將幼苗轉移至30°C溫室中培養21 d, 用尺子測量5株幼苗的主根長, 然后用i800掃描儀掃描總根長, 用LA-S根分析系統(萬深, 中國杭州)進行分析。用亞甲基藍比色法(Leagene, 中國北京)測定根系活性(總吸收面積和比表面積), 每3株幼苗為1次重復, 3個重復, 總體進行2次重復實驗, 記為重復I (Repeat I)和重復II (Repeat II)。
用均勻分布于水稻12條染色體的166個在9311和NPB之間存在多態的SSR和STS標記構建了RIL群體的遺傳連鎖圖譜。使用MultiQTL軟件(www. multiqtl.com)對RIL群體采用最大似然區間映射方法進行QTL分析, 根據每個數據集的排列測試(1000個排列,=0.05)獲得LOD閾值。根據McCouch等[22]的方法命名QTL。利用Microsoft Excel軟件分析根部形狀的表型值與頻數分布, 并繪制相關圖表; 利用IBM SPSS Statistics 24軟件進行顯著性差異檢驗和相關性分析。
通過近等系NIL-與9311的雜交和自交, 構建了用于精細定位的830個植株構成的F2群體, 測量了單株根長表型, 在已定位的區間內開發了4個InDel標記(表1)檢測單株基因型, 用于的精細定位。
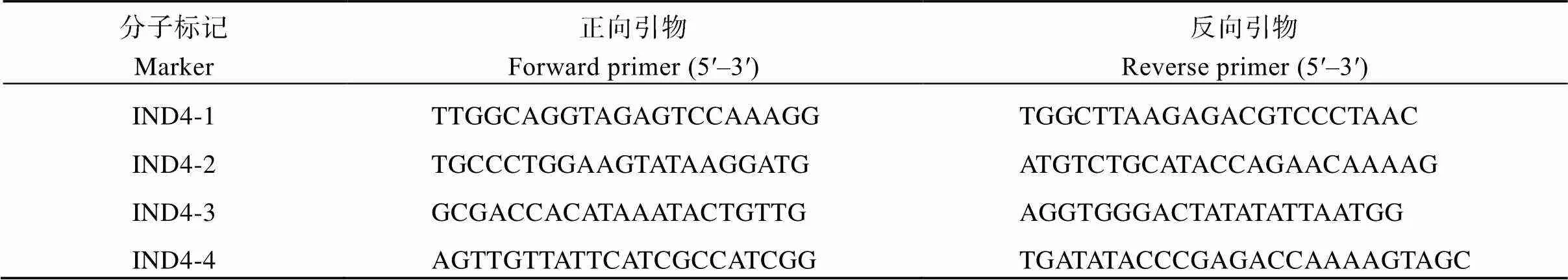
表1 qLRL4精細定位的InDel標記引物
采用Axygen公司的AxyPrep總RNA小量制備試劑盒(AP-MD-MS-RNA-50)提取9311、NPB和NIL-的根部RNA并使用TOYOBO公司的cDNA逆轉錄試劑盒(fsq-101)反轉錄得到cDNA后, 用候選基因引物和內參引物(表2)在ABI7900HT Real-time PCR儀上對樣品的候選基因表達量進行相對定量檢測。

表2 候選基因實時定量PCR引物
2 結果與分析
2.1 9311、NPB及其RIL群體的根系性狀
親本9311和NPB之間的根系形態存在顯著差異(表3和圖1)。9311的最長根長(the longest root length, LRL)、總根系長(total root length, TRL)、根表面積(root surface area, RSA)、根體積(root volume, RV)和根直徑(root diameter, RD)均顯著大于NPB。考察9311和NPB構建的RIL群體的最長根長、總根系長、根表面積、根體積和根直徑這5個性狀, 統計2次重復的平均值, 最長根長為14.8 cm和14.3 cm, 總根系長為296.35 cm和331.61 cm, 根表面積為25.26 cm2和25.62 cm2, 根體積為0.22 cm3和0.26 cm3, 根直徑為0.25 mm和0.23 mm。RIL的5個性狀與兩親本相比, 最長根長和平均直徑均小于兩親本。RIL的平均總根系長、根表面積和根體積介于NPB和9311之間(表4)。在2次重復中, 5個根系性狀在RIL群體中均呈連續正態分布(圖2), 且大部分呈現出明顯的超親分離, 表明這些性狀由多基因控制, 適合進行QTL分析。2次重復之間比較后發現, 重復I和重復II的最長根長、根表面積和根體積的分布范圍基本一致。然而, 重復I的總根長和平均直徑的變化范圍大于重復II。
2.2 RIL群體各根系性狀間的相關性分析
將RIL群體各根系性狀進行相關性分析發現(表5), 在重復I中, 最長根長與總根系長、根表面積、根體積均呈顯著正相關, 相關系數為0.359~0.399, 與根直徑沒有顯著相關性。總根系長與根表面積和根體積的相關系數為0.946和0.859, 遠大于與最長根長和根直徑的0.359和0.408。同樣, 根表面積與根體積間具有顯著相關性。總的來說, 這5個性狀, 最長根長與根直徑沒有顯著的相關性, 其他性狀之間都存在顯著正相關。而在重復II中, 最長根長與其他4個性狀間均無明顯相關性, 其他性狀相互之間都存在顯著正相關。
NPB: 日本晴; 標尺為3 cm。NPB: Nipponbare; Bar: 3 cm.
2.3 RIL群體根系性狀的QTL分析
利用一些SSR和STS標記引物進行QTL定位(表6), 然后對9311和NPB構建的RIL群體的最長根長、總根系長、根表面積、根體積和根直徑這5個性狀進行QTL分析(圖3和表7)。重復I共檢測到15個QTL, 分布在水稻第1、2、4、7、10和11染色體上。控制最長根長的QTL有3個, 分布在第1、4和10號染色體, 變異貢獻率幅度為4.8%~10.3%。其中, 貢獻率超過5%的位點有2個:和, 貢獻率分別為10.3%和6.3%, 前者效應來自NPB, 后者效應來自9311。我們檢測到控制總根系長的QTL 4個, 分布在水稻第1、2、7和11號染色體上, LOD值為2.92~8.47。這些QTL的表型變異貢獻率均超過5%, 幅度為7.8%~42.5%。的效應來自NPB, 其余3個QTL的效應來自9311。控制根表面積的QTL有3個, 分布于水稻第2、7和11號染色體, 3個QTL的表型變異貢獻率均超過5%。控制根體積的QTL有4個, 分布于水稻第2、7、10和11號染色體, 貢獻率(除外)都超過5%。控制根直徑的QTL有2個, 分布在水稻第1和4號染色體, 這2個QTL的貢獻率均超過5%。的效應來自9311,的效應來自NPB。
NPB: 日本晴; 倒三角指示親本的平均值; LRL: 最長根長; TRL: 總根系長; RSV: 根表面積; RV: 根體積; RD: 根直徑。
NPB: Nipponbare; the inverted triangle indicates the average value of the parent. LRL: the longest root length; TRL: total root length; RSV: root surface area; RV: root volume; RD: root diameter.
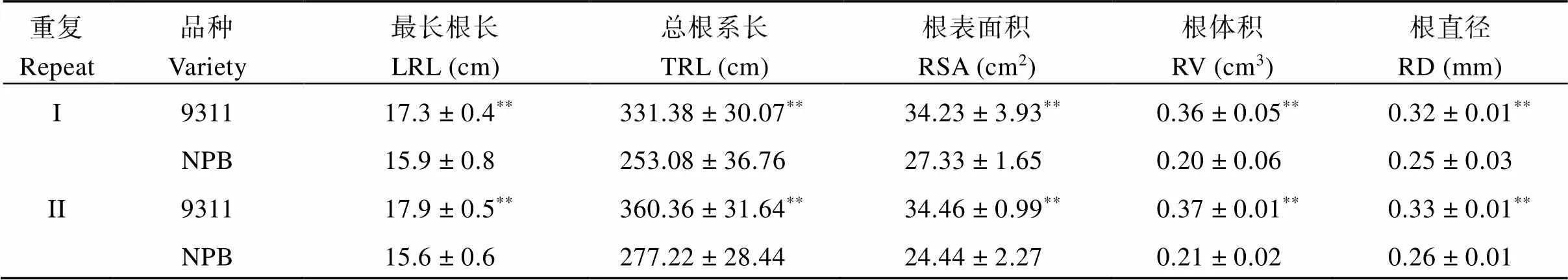
表3 9311和NPB的根系性狀
NPB: 日本晴; 值表示均值± SD;**表示達0.01顯著水平差異。LRL: 最長根長; TRL: 總根系長; RSV: 根表面積; RV: 根體積; RD: 根直徑。
NPB: Nipponbare; values represent means ± SD;**indicates significant difference at the 0.01 probability level. LRL: the longest root length; TRL: total root length; RSV: root surface area; RV: root volume; RD: root diameter.
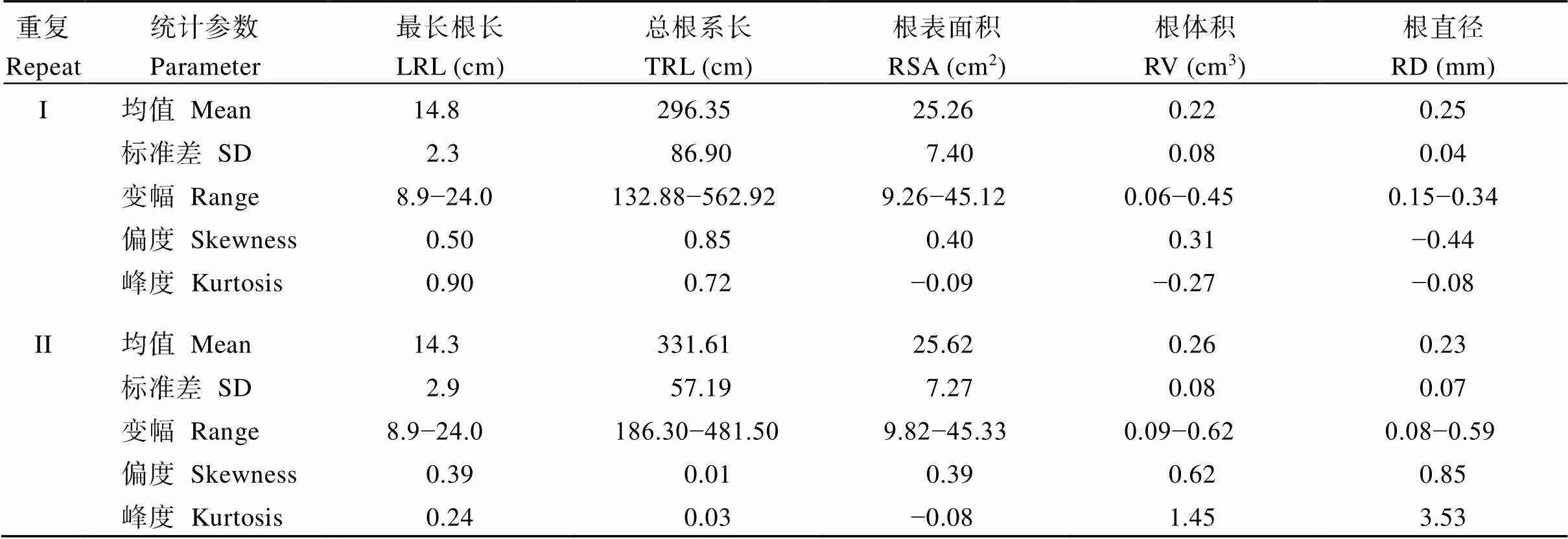
表4 RIL群體的根系性狀
LRL: 最長根長; TRL: 總根系長; RSV: 根表面積; RV: 根體積; RD: 根直徑。
LRL: the longest root length; TRL: total root length; RSV: root surface area; RV: root volume; RD: root diameter.
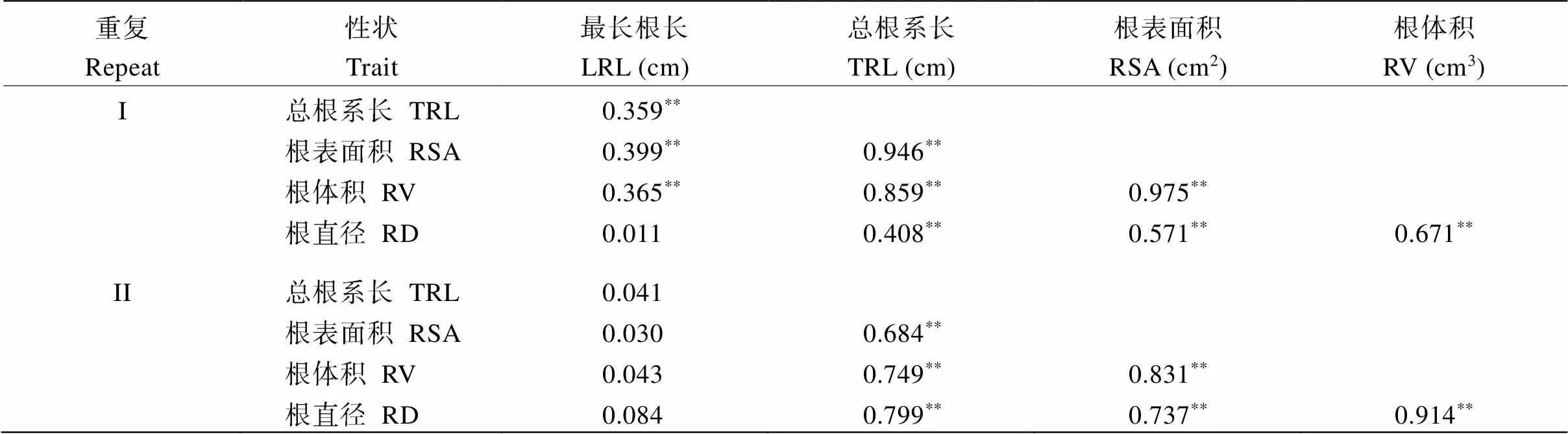
表5 RIL群體根系性狀間的相關系數
**表示0.01顯著水平相關。LRL: 最長根長; TRL: 總根系長; RSV: 根表面積; RV: 根體積; RD: 根直徑。
**indicates significant difference at the 0.01 probability level. LRL: the longest root length; TRL: total root length; RSV: root surface area; RV: root volume; RD: root diameter.

表6 QTL定位的SSR和STS標記引物
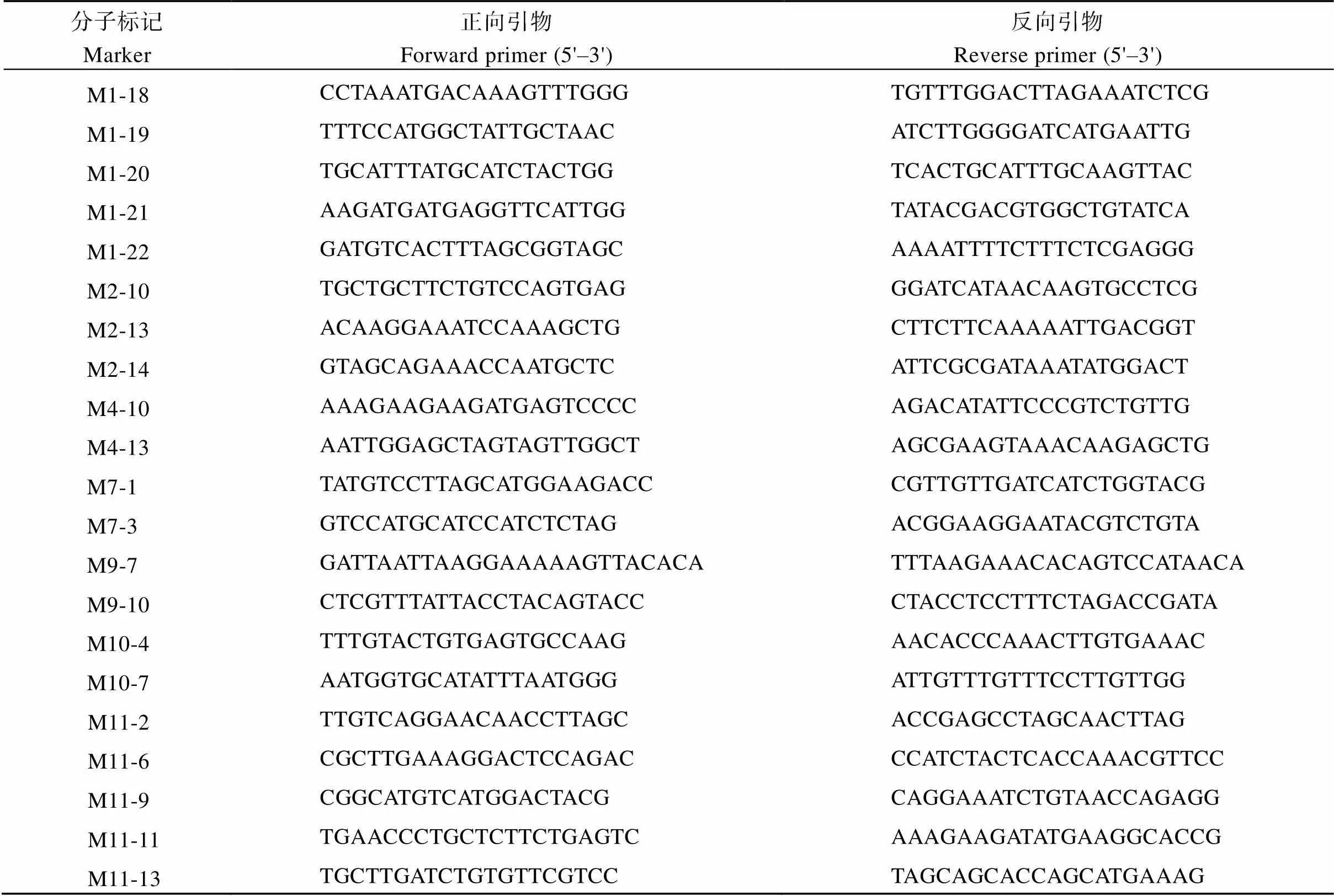
(續表6)
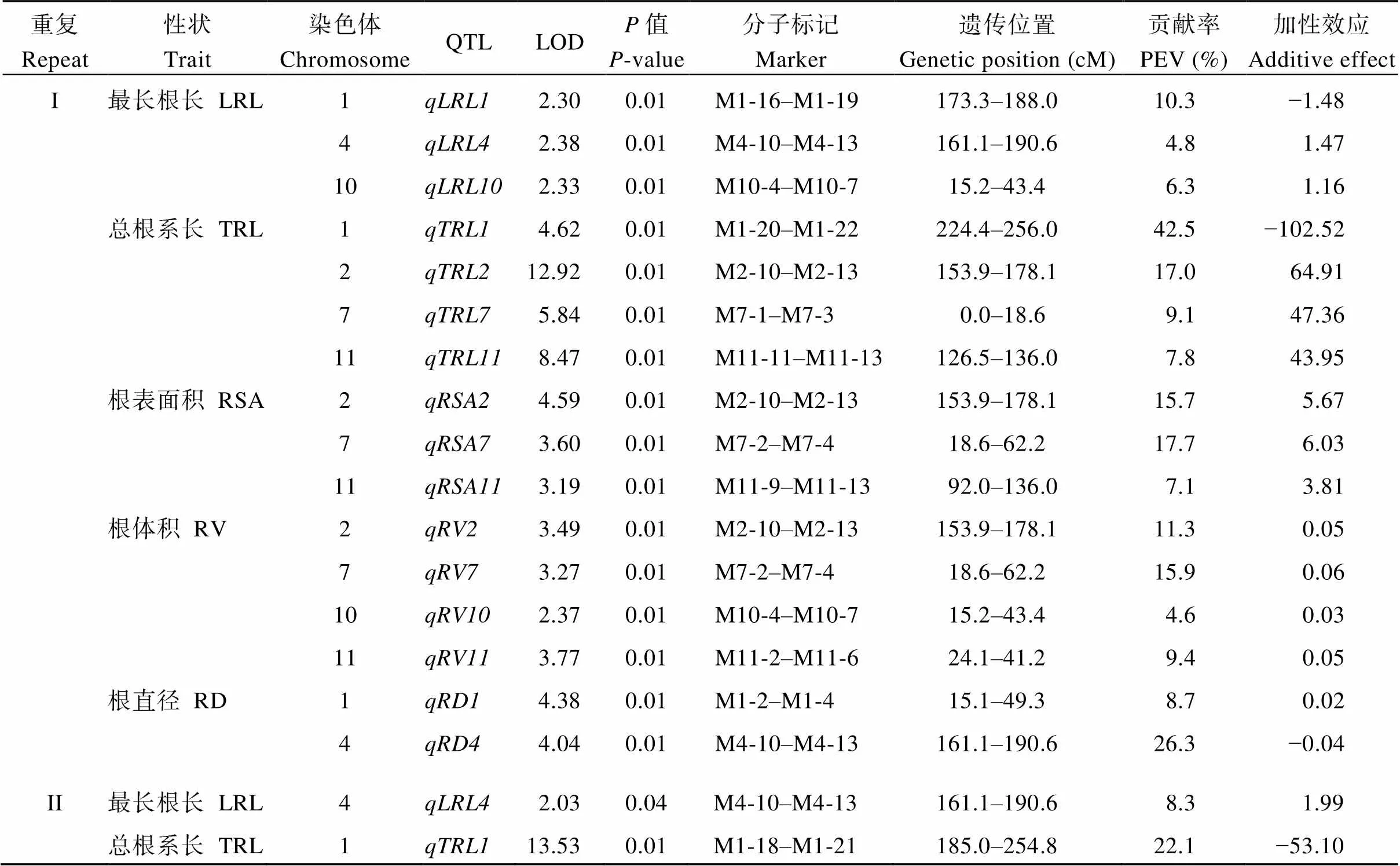
表7 RIL群體的根系性狀QTL
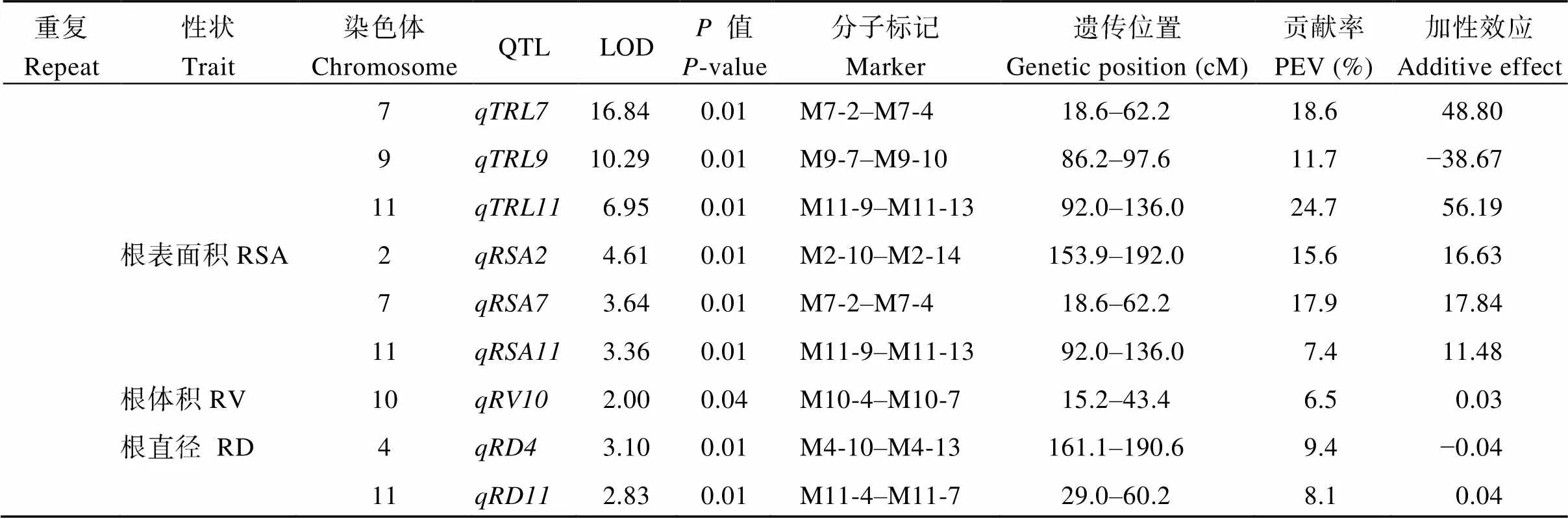
(續表7)
LRL: 最長根長; TRL: 總根系長; RSV: 根表面積; RV: 根體積; RD: 根直徑。
LRL: the longest root length; TRL: total root length; RSV: root surface area; RV: root volume; RD: root diameter.
每條染色體的左邊列出為分子標記。
Molecular markers for QTLs are listed on the left along each chromosome.
重復II共檢測到11個QTL, 分布在第1、2、4、7、9、10和11號染色體上。我們只在4號染色體上發現了一個控制最長根長的QTL, 貢獻率為8.3%, 加性效應來自于9311。控制總根長的QTL有4個, 分布在水稻第1、7、9和11號染色體上, LOD值為6.95~16.84。這些QTL的表型變異貢獻率均超過5%, 幅度為11.7%~24.7%。和的效應來自NPB, 其余2個QTL的效應來自9311。控制根表面積的QTL有3個, 分布于水稻第2、7和11號染色體, 3個QTL的表型變異貢獻率均超過5%。控制根體積的QTL有1個, 在10號染色體上。控制根直徑的QTL有2個, 分布在水稻第4和11號染色體, 這2個QTL的貢獻率均超過5%。其中,的效應來自NPB,的效應來自9311。
對2個重復進行分析發現, 有5個控制相同性狀的QTL在同一位置被重復檢測到。包括4號染色體上的和, 7號染色體上的, 10號染色體上的和11號染色體上的。、、和這4個QTL的效應在2個重復中均來自9311, 說明RIL的最長根長、根表面積和根體積的正效應來自9311等位型。的效應在2個重復中來自NPB, 說明RIL根直徑受NPB等位型的正調控。綜合2個重復鑒定的QTL來看, 在7條染色體上共分布有4個QTL簇, 分別位于水稻第2、4、7和10號染色體, 只有4號染色體上檢測到的最長根長和根直徑相關QTL, 在2次重復中定位的染色體相同位置。因此, 我們對QTL開展精細定位。
2.4 最長根長QTL qLRL4的精細定位和候選基因確定
位于水稻第4號染色體的QTL是控制最長根長的QTL。9311背景下InDel標記IND4-1和IND4-4間約158 kb來自NPB的近等系NIL與9311相比, 最長根長縮短, 根直徑略微變大, 總吸收面積和活躍吸收面積減少, 其他性狀基本無變化(圖4和表8)。利用近等系NIL-和9311構建的大規模F2群體, 結合個體的InDel標記基因型和根長表型, 我們進一步精細定位這一最長根長QTL, 最終將其定位于標記IND4-1和IND4-3之間約68.23 kb的物理距離內(圖5和表1)。依據水稻基因組注釋數據庫(http://rice.plantbiology.msu.edu/ cgi-bin/gbrowse/rice/), 該區間包含了8個候選基因, 其中有7個基因存在引起氨基酸改變的單核苷酸多態(SNP)或InDel (表9)。qRT-PCR檢測8個候選基因相對表達量(圖6), 在NPB和NIL-中, 基因、和的相對表達量較9311極顯著上升; 而基因的相對表達量極顯著下降。其他4個基因除的相對表達量在NPB中極顯著上升外,、和的相對表達量在NPB 或NIL-中與9311間無顯著差異。
標尺為3 cm。Bar: 3 cm.
3 討論
在QTL分析中, 同一染色體區間存在多個QTL控制多個根系性狀, 這可能是一因多效的作用。姜樹坤等[23]以笹錦和北陸129雜交衍生的重組自交系群體為材料, 對水稻移栽后的新生根相關性狀進行研究, 共檢測到10個QTL, 分別控制根平均直徑、總根長、平均根表面積、平均根長和根數等性狀。并且對第11號染色體的分子標記C477和G320B之間的新主效QTL——同時控制根長、根表面積和根數的QTL、和進行了驗證。這些QTL簇體現了QTL的一因多效作用。由于一因多效QTL/基因的存在, 在開展水稻分子育種時, 我們可以考慮選擇單個QTL/基因同時改良多個性狀。徐曉明等[24]以超級稻協優9308衍生的重組自交系與輪回親本中恢9308回交多代群體為材料, 采用瓊脂無土栽培技術開展了水稻根長主效QTL的精細定位, 最終將其定位在水稻第4號染色體分子標記RM5687與InDel149之間約624.6 kb的范圍內。我們定位的是控制最長根長的新QTL, 其位置與報道的在4號染色體上的位置并不相同, 這可能是所用親本和群體不同所致。幼苗期根的生長為作物獲取水和營養物質提供了保障[25]。近等系NIL-在發芽后根的生長受到抑制, 根的總吸收面積和活躍吸收面積相對9311也顯著減小(圖4和表8)。Raffaele等[26-27]在擬南芥中研究發現, 具有較大根分生組織的突變體往往根較長, 而根分生組織的大小受生長素和細胞分裂素的共同調節。水稻中已克隆了一些與根發育相關的基因, 例如基因調控不定根的數目[15],基因調控不定根的數目與長短[18],基因調控側根和不定根的數量[28],通過根系與乙烯的特異性反應來影響根的生長發育[29]。調控水稻根長的基因也有報道,基因影響水稻主根長短[30];基因通過外源性蔗糖、葡萄糖和果糖的調控影響水稻根系的長短[13];基因通過影響根細胞的長短與數目影響水稻根系的長短[14];基因影響水稻苗期根長與直徑的作用可能與ABA的感知或信號傳導有關[20]。我們的研究最終將最長根長QTL精細定位到約68.23 kb的區間內, 依據基因注釋發現該區間包含8個候選基因, 其中有7個基因存在引起氨基酸改變的SNP或InDel, 包括編碼水解酶、受體蛋白激酶和染色質結構調節因子的基因。根部轉錄水平的表達分析顯示, 4個候選基因的相對表達量在NIL-或NPB與9311之間存在顯著差異。因此, 這4個基因作為候選基因的可能性更大。當然, 候選基因的最終確定還有待進一步基因克隆和功能驗證。

表8 9311和NIL-qLRL4的根系性狀和活性
**表示達0.01顯著水平差異。
**indicates significant difference at the 0.01 probability level.
被縮小到標記IND4-1和IND4-3之間的68.23 kb區間內。圖例的黑色條塊代表9311基因型, 白色條塊代表NPB基因型。NPB: 日本晴。最長根長表示為均值±標準差。*和**分別表示0.05和0.01顯著水平差異。LRL: 最長根長。
was narrowed down to a 68.23 kb interval defined by markers IND4-2 and IND4-3. Black box represents 9311 genotype, and white box represents NPB genotype. NPB: Nipponbare. Values represent means ± SD.*and**indicate significant difference at the 0.05 and 0.01 probability levels compared with NIL-, respectively. LRL: the longest root length.

表9 qLRL4精細定位區間的候選基因和候選基因中引起氨基酸改變的序列變化
SNP和InDel為引起氨基酸改變的SNP數目和InDel數目。字母為改變的堿基, 數字為CDS上的位點。
SNP and InDel represent the number of SNPs and InDels that cause amino acid changes.The letters are changed bases and the numbers are positions on the CDS.
相對表達量數據為平均數±標準差,= 3。**表示與9311比較達到0.01的差異顯著水平。
Data represent means ± SD of three biological replicates.**indicates significant difference at the 0.01 probability level compared with 9311.
4 結論
利用秈稻9311和粳稻日本晴為親本的RIL群體對水稻苗期根系相關性狀開展QTL分析, 共檢測到26個控制最長根長、總根系長、根表面積、根體積和根直徑的QTL, 發現水稻第2、4、7和10號染色體上的4個QTL簇, 包括第4號染色體上控制最長根長的QTL。精細定位將定位于InDel標記IND4-1和IND4-3之間約68.23 kb的區間, 包含了8個候選基因。其中7個基因在親本間有引起氨基酸改變的SNP或InDel, 4個基因在近等基因系NIL-和9311之間的轉錄水平相對表達量上存在顯著差異。此最長根長QTL的精細定位為水稻根系發育相關基因的克隆奠定了基礎。
[1] 范楚玉. 西周農事詩中反映的糧食作物選種及其發展. 自然科學史研究, 1982, 3: 267–272.
Fan C Y. Selection and development of grain crops reflected in the poetry of agriculture in the Western Zhou Dynasty, 1982, 3: 267–272 (in Chinese with English abstract).
[2] 陶榮榮, 蔡晗, 朱慶權,周益雷, 王康平, 余超, 侯丹平, 劉海浪, 張耗. 水稻高產高效的根-冠互作機制研究進展. 中國農學通報, 2018, 34(5): 1–4.
Tao R R, Cai H, Zhu Q Q, Zhou Y L, Wang K P, Yu C, Hou D P, Zhang H. Research progress on root-crown interaction mechanism of high-yield and high-efficiency rice., 2018, 34(5): 1–4 (in Chinese with English abstract).
[3] 丁仕林, 劉朝雷, 錢前. 水稻根系遺傳研究進展. 中國稻米, 2019, 25(5): 24–29.
Ding S L, Liu C L, Qian Q. Advances in rice root genetics., 2019, 25(5): 24–29 (in Chinese with English abstract).
[4] 徐吉臣, 李晶昭, 鄭先武, 鄒亮星, 朱立煌. 苗期水稻根部性狀的QTL定位. 遺傳學報, 2001, 28: 433–438.
Xu J C, Li J Z, Zheng X W, Zou L X, Zhu L H. QTL mapping of rice root traits at seedling stage., 2001, 28: 433–438 (in Chinese with English abstract).
[5] 滕勝, 曾大力, 錢前, 國廣泰史, 藤本寬, 黃大年, 朱立煌. 水稻根系活力的遺傳分析. 中國水稻科學, 2002, 16: 119–123.
Teng S, Zeng D L, Qian Q, Kunihiro Y, Fujimoto K, Huang D N, Zhu L H. Genetic analysis of root vigor in rice., 2002, 16: 119–123 (in Chinese with English abstract).
[6] 胡興明, 郭龍彪, 曾大力, 高振宇, 滕勝, 李浩戈, 朱立煌, 錢前. 水稻苗期發根力的QTL和上位性分析. 中國水稻科學, 2004, 18: 396–400.
Hu X M, Guo L B, Zeng D L, Gao Z Y, Teng S, Li H G, Zhu L H, Qian Q. QTL mapping and epistasis analysis of rice root growth ability at seedling stage., 2004, 18: 396–400 (in Chinese with English abstract).
[7] Mitsuhiro O, Wataru T, Takeshi E. Fine-mapping of, a major QTL for root length of rice seedlings grown under a wide range of NH4+concentrations in hydroponic conditions., 2010, 121: 535–547.
[8] 王汝慈. 兩個生育時期水稻耐低磷脅迫相關性狀的QTL定位. 中國農業科學院碩士學位論文, 北京, 2009.
Wang R C. QTL Mapping of Low Phosphorus Stress-related Traits in Rice during Two Growth Periods. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2015 (in Chinese with English abstract).
[9] Obara M, Fukuta Y, Yanagihara S. Genetic variation and QTLs related to root development in upland new rice for(NERICA) varieties., 2019, 69: 94–103.
[10] Kitomi Y, Nakao E, Sawako K. Fine mapping of quick rooting 1 and 2, quantitative trait loci increasing root length in rice., 2018, 8: 727–735.
[11] 章怡蘭, 林雪, 吳儀, 李夢佳, 張晟婕, 路梅, 饒玉春, 王躍星. 水稻根系遺傳育種研究進展. 植物學報, 2020, 55: 382–393.
Zhang Y L, Lin X, Wu Y, Li M J, Zhang S J, Lu M, Rao Y C, Wang Y X. Research progress on rice root genetics and breeding., 2020, 55: 382–393 (in Chinese with English abstract).
[12] 梁永書, 周軍杰, 南文斌, 段東東, 張漢馬. 水稻根系研究進展. 植物學報, 2016, 51: 98–106.
Liang Y S, Zhou J J, Nan W B, Duan D D, Zhang H M. Research progress of rice root system., 2016, 51: 98–106 (in Chinese with English abstract).
[13] Yao S G, Mushika J, Taketa S, Ichii M. The short root mutationdefines a sugar-mediated root growth in rice (L.)., 2004, 167: 49–54.
[14] Jia L, Zhang B, Mao C. OsCYT-INV1 for alkaline/neutral invertase is involved in root cell development and reproductivity in rice (L.)., 2008, 228: 51–59.
[15] Inukai Y, Sakamoto T, Ueguchitanka M. Crown rootless1, which is essential for crown root formation in rice, is a target of an auxin response factor in auxin signaling., 2005, 17: 1387–1396.
[16] Liu H, Wang S, Yu X, Yu J, He X, Zhang S, Shou H, Wu P. ARL1, a LOB-domain protein required for adventitious root formation in rice., 2005, 43: 47–56.
[17] Zhao Y, Hu Y F, Dai M G, Huang L M, Zhou D Y. The WUSCHEL-related homeobox geneis required to activate shoot-borne crown root development in rice., 2009, 21: 736–748.
[18] Yang S Q, Li W Q, Miao H, Gan P F, Qiao L, Chang Y L, Shi C H, Chen K M., a gene encoding an unknown function protein which contains DUF630 and DUF632 domains controls leaf rolling in rice., 2016, 9: 1–14.
[19] Dai X Y, Wang Y Y, Zhang W H. OsWRKY74, a WRKY transcription factor, modulates tolerance to phosphate starvation in rice., 2016, 67: 947–960.
[20] Ao S G, Shin T, Masahiko I. Isolation and characterization of an abscisic acid-insensitive mutation that affects specifically primary root elongation in rice (L.)., 2003, 164: 971–978.
[21] Jing H W, Yang X L, Zhang J, Liu X H, Zheng H K, Dong G J, Nian J Q, Feng J, Xia B, Qian Q, Li J Y, Zuo J R. Peptidyl-prolyl isomerization targets rice Aux/IAAs for proteasomal degradation during auxin signaling., 2015, 6: 7395.
[22] McCouch S, Cho Y, Yano M, Paul E, Blinstrub M, Morishima H, Kinoshita T. Report on QTL nomenclature., 1997, 14: 11–131.
[23] 姜樹坤, 張鳳鳴, 白良明, 孫世臣, 王彤彤, 丁國華, 姜輝, 張喜娟. 水稻移栽后新生根系相關性狀的QTL分析. 中國水稻科學, 2014, 6: 598–604.
Jiang S K, Zhang F M, Bai L M, Sun S C, Wang T T, Ding G H, Jiang H, Zhang X J. QTL analysis of the related traits of new roots after transplanting rice., 2014, 6: 598–604 (in Chinese with English abstract).
[24] 徐曉明, 張迎信, 王會民, 任翠, 王汝慈, 沈希宏, 占小登, 吳瑋勛, 程式華, 曹立勇. 一個水稻根長QTL的分離鑒定. 中國水稻科學, 2016, 30: 363–370.
Xu X M, Zhang Y X, Wang H M, Ren C, Wang R C, Shen X H, Zhan X D, Wu W X, Cheng S H, Cao L Y. Isolation and identification of a QTLof rice root length., 2016, 30: 363–370 (in Chinese with English abstract).
[25] Rogers E D, Benfey P N. Regulation of plant root system architecture: implications for crop advancement., 2015, 32: 93–98.
[26] Raffaele D I, Francisco S L, Emanuele S. Cytokinins determineroot-meristem size by controlling cell differentiation., 2007, 17: 678–682.
[27] Raffaele D I, Kinu N, Laila M. A genetic framework for the control of cell division and differentiation in the root meristem., 2008, 322: 1380–1384.
[28] Liu W, Xu Z H, Luo D. Roles of OsCKI1, a rice casein kinase I, in root development and plant hormone sensitivity., 2003, 36: 189–202.
[29] Chen H, Ma B, Zhou Y, He S J, Tang S Y, Lu X, Xie Q, Chen S Y, Zhang J S. E3 ubiquitin ligase SOR1 regulates ethylene response in rice root by modulating stability of Aux/IAA protein., 2018, 115: 4513–4518.
[30] Zhang H G, Zhang L J, Si H, Ge Y S, Liang G H, Gu M H, Tang S Z. Rf5 is able to partially restore fertility to Honglian-type cytoplasmic male sterilerice () lines., 2016, 36: 1–10.
Genetic analysis of seedling root traits and fine mapping of the QTLfor the longest root length in rice
TIAN Biao, DING Shi-Lin, LIU Chao-Lei, RUAN Ban-Pu, JIANG Hong-Zhen, GUO Rui, DONG Guo-Jun, HU Guang-Lian, GUO Long-Biao, QIAN Qian, and GAO Zhen-Yu*
China National Rice Research Institute, Hangzhou 310006, Zhejiang, China
In order to analyze the genetic basis of root traits at seedling stage, we performed QTL analysis of root morphology with 148 recombinant inbred lines derived fromvariety 9311 andvariety Nipponbare (NPB). In two repetitions, a total of 26 QTLs were detected for the longest root length, total root length, root surface area, root volume, and root diameter, distributed on chromosomes 1, 2, 4, 7, 9, 10, and 11 in rice. Four QTL clusters on chromosomes 2, 4, 7, and 10 were found, including a major QTLcontrolling the longest root length. To fine mapping of the major QTL, we constructed a near isogenic line NIL-with a segment from NPB between markers IND4-1 and IND4-4 with 9311 background. With a F2population derived from the NIL-and 9311, we fine mapped thewithin ~68.23 kb region between markers HIND4-1 and IND4-3, where eight candidate genes located. Fine mapping of this QTL for root length will help explore genetic mechanism of root elongation and morphogenesis in rice.
rice; root trait; the longest root length;; fine mapping
10.3724/SP.J.1006.2021.02088
本研究由國家自然科學基金項目(32061143039, 31671761)資助。
This study was supported by the National Natural Science Foundation of China (32061143039, 31671761).
高振宇, E-mail: gaozhenyu@caas.cn, Tel: 0571-63370211
E-mail: 82101186057@caas.cn, Tel: 0571-63370483
2020-12-13;
2021-03-19;
2021-04-13.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20210413.1521.006.html
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
