Mdm2介導的正反饋環對p53基因網絡振蕩行為的影響*
2021-08-04 08:35:26楊紅麗劉楠楊聯貴
物理學報 2021年13期
關鍵詞:模型
楊紅麗 劉楠 楊聯貴
(內蒙古大學數學科學學院, 呼和浩特 010021)
轉錄因子p53是細胞應激網絡的核心, 以動態響應的方式控制基因毒性壓力下的細胞命運抉擇.Mdm2是種E3泛素連接酶, 既能破壞p53的穩定性又能提高p53的生成效率.Mdm2對p53的抑制性功能在p53-Mdm2振子中扮演著構建性角色, 而Mdm2對p53的促進性功能如何調控這個基因網絡的動力學仍缺少研究.因此, 本文利用數學模型, 全面探究了Mdm2上調p53的這條通路對p53動力學的影響.結果表明:Mdm2在Ser395位點的磷酸化作用對p53的振蕩必不可少; 之前報道的磷酸酶Wip1被p53振蕩所需要, 可能僅發生在Mdm2所介導的正反饋通路強度較高的情景下; Mdm2促進p53的失活以及泛素化降解也是p53反復振動動力學發生的關鍵因素, 與以往的結論一致.本文的結果可對今后p53動力學的相關實驗起到一定的指導作用.
1 引 言
癌癥是種細胞信號通路的疾病, 嚴重威脅著人類生命健康[1].抑癌因子p53作為細胞信號轉導網絡的樞紐, Puszyński等[2]斷言在人類癌癥中約有一半會出現p53自身的突變, 而另一半會出現p53調控子的突變.在靜息細胞中, E3泛素連接酶Mdm2 (murine double minute 2) 使得p53多泛素化, 進而被蛋白酶體識別并快速降解; 此時p53的濃度被抑制在一個基底水平[3].而在缺氧、紫外線照射、依托泊苷處理、電離輻射等細胞壓力下, p53被穩定并激活[4?7].活化的p53與靶基因的綁定親和力增強, 誘導多種下游蛋白的表達.這些下游蛋白參與到細胞周期停滯、DNA修復、細胞程序性死亡等生理過程中[8].據實驗報道, 細胞命運的信息被p53動力學編碼[9].因此, 還有一些問題亟需解決.例如, 解碼p53動力學所關聯的細胞命運抉擇的細致機制是個極具應用價值的問題;研究p53蛋白信號網絡中本身的動力學控制也是個很有意義的問題, 等等.本文主要關注的是有關p53蛋白信號網絡中的動力學控制問題, 即Mdm2在Ser395位點磷酸化這條信號通路對p53動力學調控的問題.
2000年, Lev Bar-Or等[10]做了一項關于p53動力學的實驗, 發現電離輻射下細胞群體中p53的衰減式脈沖, 也就是阻尼振蕩.后來的實驗在單細胞中觀測, 發現p53的振蕩是無阻尼的, 并且脈沖的個數與輻射劑量正相關[11,12].后續的很多理論模型圍繞p53的反復式脈沖動力學開展, 例如, 張麗娟等[13]用p53-Mdm2負反饋環附加時滯構建的振子模型揭示了群體細胞p53的阻尼振蕩可能是單細胞p53振蕩持續時間不一、脈沖數量差異的結果;Bottani和Grammaticos[14]研究了帶時滯的p53-Mdm2振子在p53, Mdm2半衰期參數控制下的分岔現象; Ma等[15]建立了復雜的混合模型 (隨機過程對接微分方程) 來模擬p53的數字式脈沖;Xia和Jia[16]重建了包含p53振子模塊和DNA修復模塊的簡化模型, 用更簡單的方式解釋了p53脈沖數正比于輻射劑量的可行性機制;等[17]探索了單細胞中Wip1 (wild type p53-induced phosphatase 1) 參與下p53發生振蕩的常微分模型和基于反應擴散的偏微分模型; 畢遠宏等[18]研究了PDCD5 (programmed cell death 5) 和Mdm2最大生成速率調控下的p53全局動力學與穩定性等問題, 展示了p53動力系統中存在的豐富動力學現象; Hat等[19]研究了p53基因網絡中常見的正負反饋通路及反饋背后的雙穩態、振蕩等分岔機制; 王道光等[20]提取了p53-Mdm2負反饋環和
p53-Wip1-ATM (ataxia telangiectasia-mutated)
負反饋環共存的模型, 分析了p53信號轉導網絡中的雙節率現象; Ochab和Puszynski[21]開發了一種分段線性方法, 并以p53振子為例對比了新方法與傳統的非線性方法; 劉楠等[22]重建了p53-Mdm2通路和p53-Wip1-ATM通路耦合的模型, 在Ser395位點磷酸化的Mdm2被考慮在內, 并探索了這種情景下Wip1管理的p53潛在動力學; 等等.雖然p53-Mdm2之間的正反饋被提及, 但據我們所知,尚未有細致的研究.
基于上述考慮, 本文重新審視了Mdm2促進p53的作用對p53動態的影響.首先給出這個基因網絡模型對應的動力學方程; 然后利用分岔分析的辦法, 找出了關鍵的分岔點, 系統的動力學特征由分岔點劃分的參數區間所決定; 最后根據分岔曲線, 在二維參數平面上找出了振蕩的區域.數值研究結果表明: p53-Mdm2之間正反饋強度的差異可能是p53面對兩種都能激活ATM的細胞壓力時出現不同動力學響應方式的關鍵原因; Wip1對這個基因網絡的振蕩會時而有利時而不利, 取決于Mdm2對p53的正調控強度; 在這個模型中, p53-Mdm2的正負雙重反饋, 是振蕩出現的根基, 也就是說p53-Mdm2之間的正負反饋對p53振蕩而言是同等重要的.總之, 振蕩出現需要正負兩種反饋強度接近、相互偶聯.
2 模型與方程
事實上, p53的動力學行為是壓力類型依賴的[23].振蕩的p53動力學出現在那些能引起DNA雙鏈斷鏈 (DSB) 的細胞壓力中, 例如電離輻射和依托泊苷刺激[12,24].ATM作為DSB的信號傳感器, 接收到DNA損傷信號并迅速傳導到p53-Mdm2模塊[25].一方面激活p53的轉錄活性, 另一方面通過催化Mdm2在Ser395位點的磷酸化促使Mdm2自我泛素化并被快速分解來增強p53的穩定性[26].在Ser395位點磷酸化的Mdm2能結合p53的信使RNA, 使其翻譯效率被大幅提升[27].此外, Mdm2還有一種在Ser166或Ser168位點的磷酸化方式, 這類Mdm2的作用是促進p53失活和降解[28].Mdm2是p53的靶蛋白之一, 實驗表明另一種p53的靶蛋白Wip1也對維持形狀均勻的p53脈沖至關重要[29].作為一種蛋白激酶, Wip1能促進ATM的去磷酸化, 從而抑制DNA損傷誘導下的ATM激活.因此, p53-Mdm2之間既有正反饋環又有負反饋環, 而p53-Wip1-ATM僅形成了負反饋環, 如圖1所示, 其中紅色線形成的回路為負反饋環, 綠色線形成的回路為正反饋環.
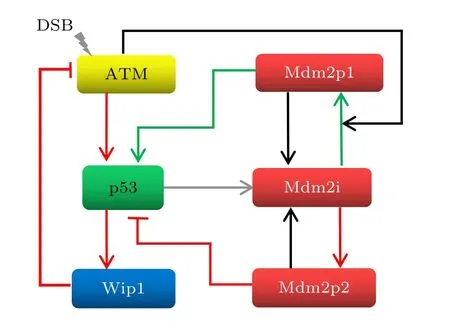
圖1 模型示意圖.箭頭線表示促進或狀態轉移; 杠頭線表示抑制Fig.1.Schematic diagrams of the model.Arrow-headed lines indicate promotion or state transition; bar-head lines indicate inhibition.
要構建基因網絡的動力學模型, 首先要明確網絡中的節點.我們的模型中共有6個節點: 活性的ATM, 活性的p53, Wip1, 在Ser395位點磷酸化的Mdm2 (Mdm2p1), 未修飾的Mdm2 (Mdm2i), 在Ser166或Ser168位點磷酸化的Mdm2 (Mdm2p2).根據模型所考慮的生化反應 (也就是ATM被DSB激活, 被Wip1滅活; p53的激活需要ATM,合成受到Mdm2p1影響; Wip1基因的轉錄依賴于p53; Mdm2基因的轉錄也依賴p53; 不考慮能終止p53振蕩的Akt通路[30], Mdm2可自由地在Ser166或Ser168位點被磷酸化以及去磷酸化, 導致Mdm2i與Mdm2p2相互轉化; Mdm2被ATM催化轉變為Mdm2p1; Mdm2p1又能自發地去磷酸化, 變回Mdm2i), 可給出如圖1 所示的模型框架.網絡中的每個節點對應著一個方程.用方括號“[]” 表示無量綱的蛋白質濃度, 在方括號里寫了蛋白質的種類, 用一類 “S” 形函數 (廣義的希爾函數) 來刻畫蛋白質之間的相互調控, 則由這個基因網絡“翻譯”出的動力學方程組如下:

其中濃度單位為“C”, 時間單位為分鐘(min), 希爾系數s0=s1=s2=s3=s4=s6=4 ,s5=2 , 其他的參數值及描述如表1[22,30,31]所列.在標準參數下p53的振蕩周期在4—7個小時之間 (圖2), 與實驗數據一致.
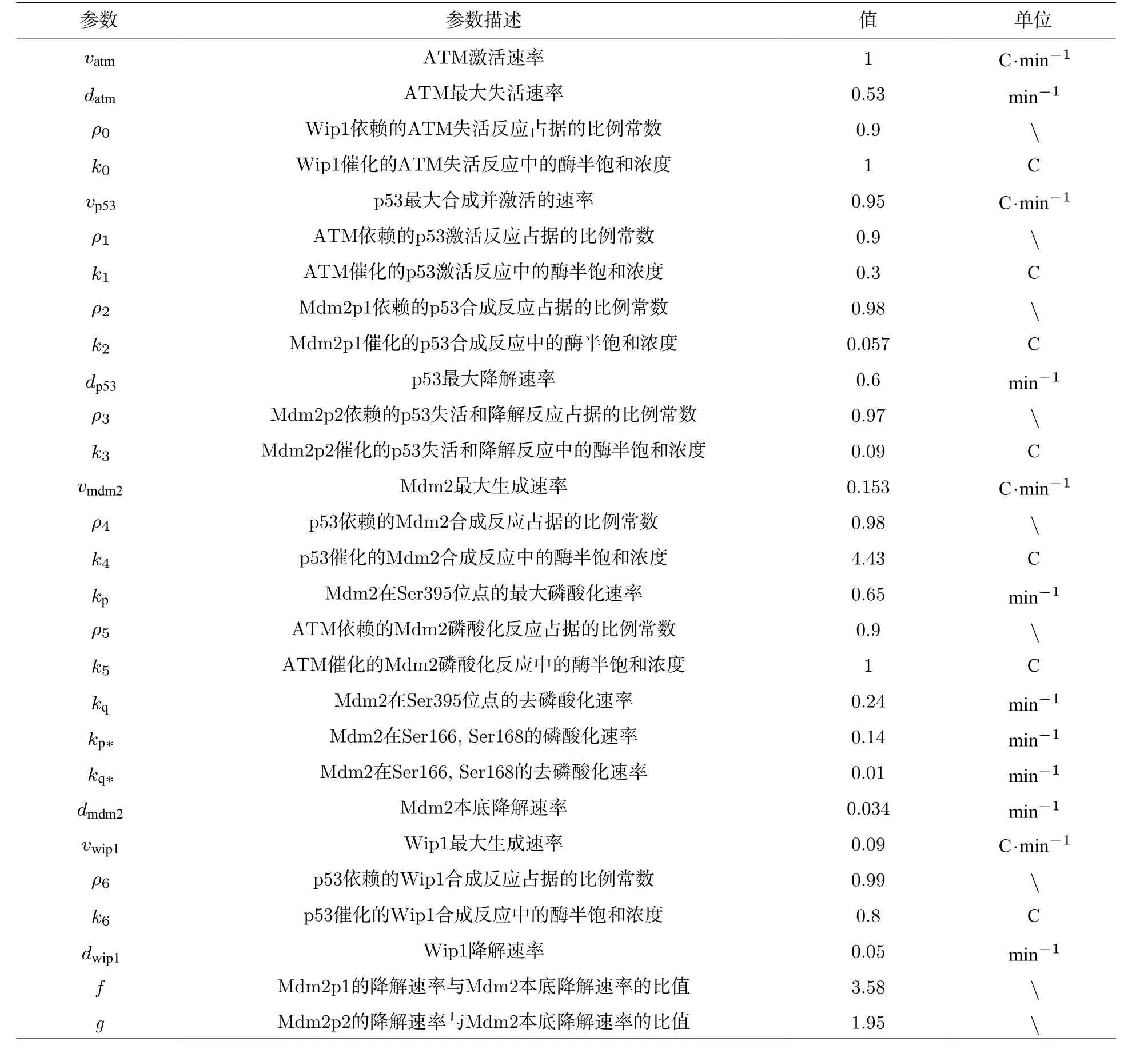
表1 模型中的參數Table 1.Parameters of the model.
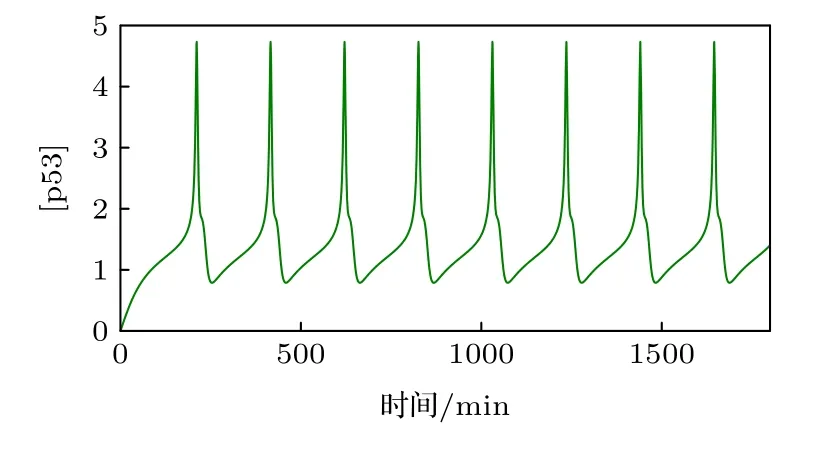
圖2 p53的時間歷程圖Fig.2.Time occurs of p53.
3 結果與分析
3.1 關于正反饋強度參數 kp 的分岔分析
Mdm2對p53正調控的必要途徑是Mdm2在Ser395位點的磷酸化, 與參數kp對應.換言之,kp的值反應了p53-Mdm2正反饋的強度.首先在圖3給出了p53平衡態與kp的函數關系, 即余維1分岔圖.在不動點分支上出現了三個分岔點: 鞍結分岔點 (SN)、鞍結同宿分岔點 (SNIC)、霍普夫分岔點 (HB).在SN和SNIC之間的藍線是不穩定的鞍點, 在SN和HB之間的紅色線是不穩定的結點和焦點, 在SNIC和HB之外的黑色線代表p53穩定的穩態.在SNIC和HB之間, 這個動力系統中僅存在穩定的極限環, 也就是p53動態行為僅有穩定振蕩.這里極限環的分支之所以沒有被延拓是因為極限環上的折疊分岔點靠近HB, 因此系統的動力學由不動點分支上的分岔點基本確定.當kp在(0, SNIC) 參數區間中時, p53最終會穩定在低穩態; 當kp在(SNIC, HB) 參數區間中時, p53動力學呈現反復的、非衰減的脈沖; 當kp在(HB, +∞)參數區間中, p53最終會達到一個較高的穩態.
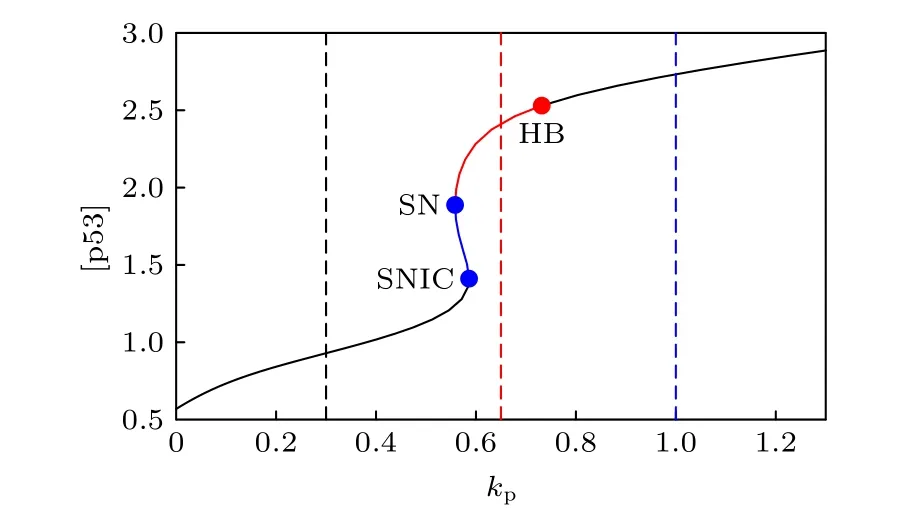
圖3 關于參數 kp 的一維分岔圖Fig.3.One-dimensional bifurcation graph on parameter kp.
為了驗證一維分岔分析的結果, 取了3個kp值在([p53], [Mdm2])相平面畫出軌線(圖4), 這里的[Mdm2]為三種形式Mdm2的濃度之和.當kp=0.3時(圖3中的黑色虛線), 分岔圖顯示p53有穩定的低穩態, 在圖4中對應著黑色軌道, 從原點開始最終停在了不動點.當kp=0.65 時(圖3中的紅色虛線), 不動點是不穩定的焦點, 在圖4中對應著紅色軌道, 始于原點的軌跡最終收斂到極限環.當kp=1 時(圖3中的藍色虛線), p53最終會穩定在較高的濃度, 在圖4中對應著藍色軌道,從原點出發沿著漸進的環狀軌跡最終到達不動點,并不再移動.相平面分析(圖4)的結果與分岔分析(圖3)的結果保持一致, 證實了分岔分析的正確性.在正反饋強度很弱的時候, 極限環消失, 說明了Mdm2介導的正反饋對p53振蕩發生的必要性.此外, 正反饋強度過強時候振蕩也會消失.實際上,正反饋過強時可通過調節一些負反饋的強度來抵消正反饋, 進而使得振蕩復現, 詳情見3.2和3.3小節.
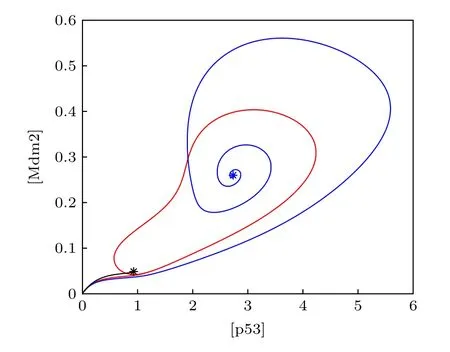
圖4 相平面分析Fig.4.Phase plane analysis.
上述的分析表明, 要獲取二維參數平面上的振蕩區域, 只需延拓SNIC和HB兩個分岔點.由于ATM的激活參數vatm是這個基因網絡接收刺激信號的開端, 受到DSB的調控, 所以我們把vatm作為第二參數張成(kp,vatm)參數平面(圖5).通過延拓SNIC分岔點(圖5中的藍色線)和HB分岔點(圖5中的紅色線), 得到二維參數平面上的兩條分岔曲線, 由這兩條分岔線圍成的灰色區域為振蕩區域.圖5表明, 當正反饋強度較弱, 即kp較小時, 例如kp=0.6 , p53的振蕩行為對DNA損傷更魯棒,也就是說只要DNA損傷輸入的信號超過某個閾值就能觸發p53振蕩.這與Lahav等[12]的實驗結果一致, 即電離輻射強度(DNA損傷程度)不能改變p53脈沖式動力學的應答模式.而正反饋強度較強, 例如kp=1 時, 過多或過少的DNA損傷都不能使p53振蕩, 與Chen等[24]的實驗一致, 即依托泊苷的劑量(DNA損傷程度)需要適中才能出現p53振蕩.因此我們推斷, 在引起DSB的兩類刺激下, p53出現對DSB相異的魯棒性有可能是p53-Mdm2之間正反饋強度的差異引起的, 這與Sun和Cui[32]的觀點(ATM誘導的Mdm2磷酸化通路有利于雙模式的p53動力學)有相似之處.
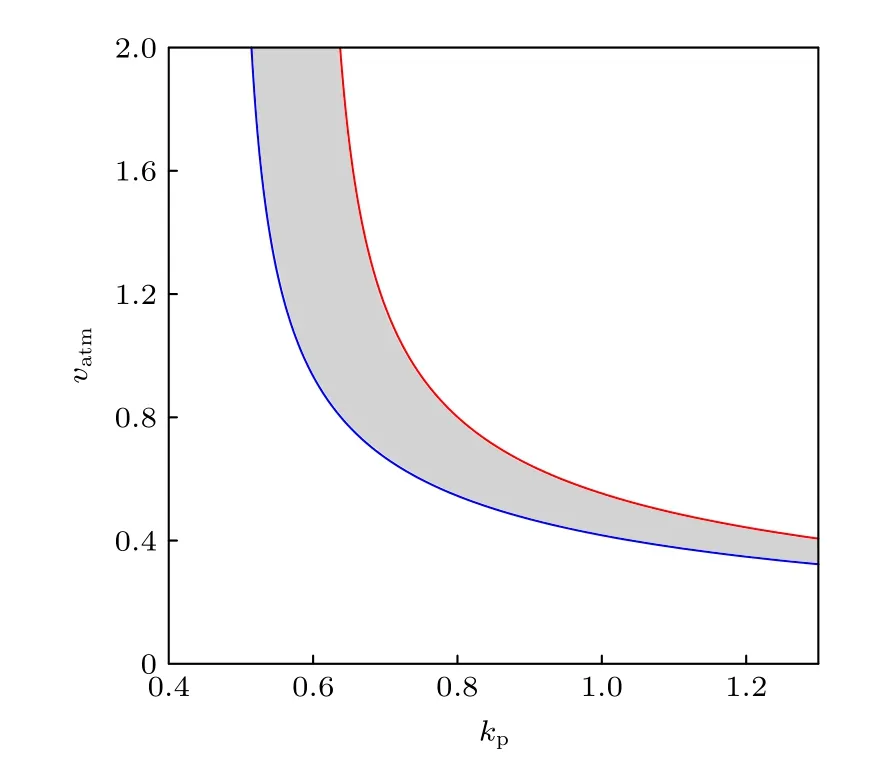
圖5 二維參數平面( kp , v atm )上的振蕩區域Fig.5.Oscillation area on the ( kp , v atm ) two-dimensional parameter plane.
3.2 p53-Wip1-ATM負反饋環與p53-Mdm2p正反饋環的聯合影響
Batchelor等[29]的實驗發現, 在細胞中加入Wip1信使RNA的抑制劑后, p53振蕩消失.此時的p53出現單個大幅度脈沖, 類似于在紫外線照射實驗中觀測到的p53脈沖.因此, Wip1通路作用強度的差異很可能是兩種刺激下出現不同的p53動力學應答的根本原因[23].為了探究Wip1和振蕩發生的關系, 在(kp,vwip1)參數平面上延拓出SNIC分岔線和HB分岔線; 同樣, 灰色區域為振蕩區域(圖6).當vwip1增大到一定程度時, 兩條分岔曲線與x軸接近垂直, 此時Wip1的作用飽和,也就是Wip1含量雖然充足, 但是反應底物活性ATM的含量較低, 使得ATM失活反應的最大速率被限制.這點在模型中體現在關于Wip1的希爾函數中, 在Wip1的濃度很高的情形下, 繼續增加Wip1濃度不再對這個基因網絡的其他節點有影響.顯然, 在標準參數下(kp=0.65 ), Wip1是振蕩出現的重要條件, 與文獻[22]結論一致.
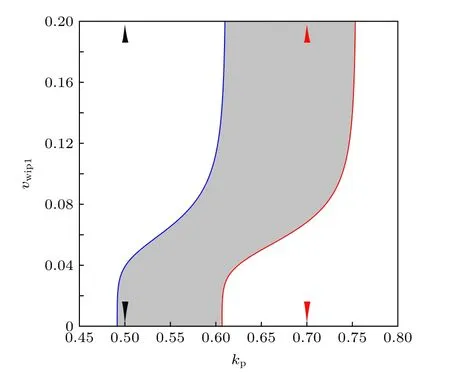
圖6 二維參數平面( kp , v wip1 )上的振蕩區域Fig.6.Oscillation area on the ( kp , v wip1 ) two-dimensional parameter plane.
有趣的是, 當kp較小時,vwip1=0 也會發生振蕩.這意味著Wip1被抑制的情況下, 通過削弱p53-Mdm2之間的正反饋也能使得p53出現振蕩,在生物實驗中應注意這一點.為了進一步說明這些情況, 在兩種kp值下做了[p53] 關于參數vwip1的余維1分岔圖(圖7).當kp較小時, 如圖6黑色小三角所指(kp=0.5 ), 此時Wip1可能不利于p53振蕩; 在圖7(a)中, 隨著vwip1增大, 到超過SNIC分岔點時, 不動點為穩定的結點, 極限環消失.而當kp較大時, 如圖6紅色小三角所 (kp=0.7 ),此時Wip1有利于p53振蕩; 在圖7(b)中, 隨著vwip1增大, 到超過HB分岔點時, 不動點為不穩定的焦點, 極限環出現.因此, 在這個模型中Wip1對系統振蕩的影響是由p53-Mdm2之間的正反饋強度決定的.非時滯模型的振蕩需要恰當的正反饋強度, 默認參數下(kp=0.65 )正反饋較強, 附加的p53-Wip1-ATM負反饋回路與p53-Mdm2p1正反饋回路拮抗, 所以能出現振蕩.
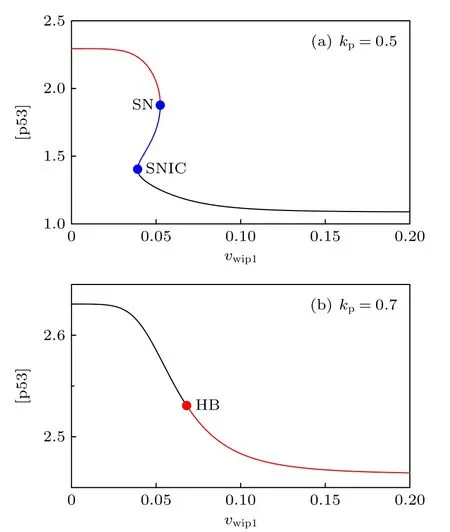
圖7 關于參數 k wip1 的一維分岔圖Fig.7.One-dimensional bifurcation graph on parameter k wip1.
3.3 p53-Mdm2正負反饋環的聯合影響
最后探究p53-Mdm2之間的正負反饋協作對系統振蕩出現的影響.模型中假設Mdm2要實現對p53的抑制作用, 即破壞p53的穩定性和轉錄活性, 首先要經過Akt的磷酸化.事實上, 在文獻[30]的模型中, Akt的活性也受到p53的間接調控, 并且這種作用很可能引起p53的雙穩態.在本文考慮的模型中,kp?被視為定常量, 是控制Mdm2抑制p53這條通路的關鍵參數.如圖8所示, 在(kp,kp?)參數平面中的振蕩區域(灰色部分)與x軸和y軸都不相交, 這代表p53-Mdm2p1通路和p53-Mdm2p2通路都是振蕩的要素.負反饋回路是振蕩發生的網絡結構基礎, 在很多p53振子的理論模型中, p53-Mdm2的負反饋都起到了重要作用[13?16], 本文的模型中起主導作用的負反饋回路同樣是p53-Mdm2.參數kp的振蕩區間(SNIC,HB)隨著kp?的增大而擴寬, 進一步說明了p53-Mdm2p2負反饋環有利于p53的振蕩.隨著負反饋強度的增加, 振蕩所需要的正反饋強度也隨之增加, 再次說明了振蕩的出現需要兩者同時參與、相互制約.事實上, Zhang等[33]提出, 用正負反饋回路耦合構建的p53振子模型能產生更具魯棒性的振蕩.
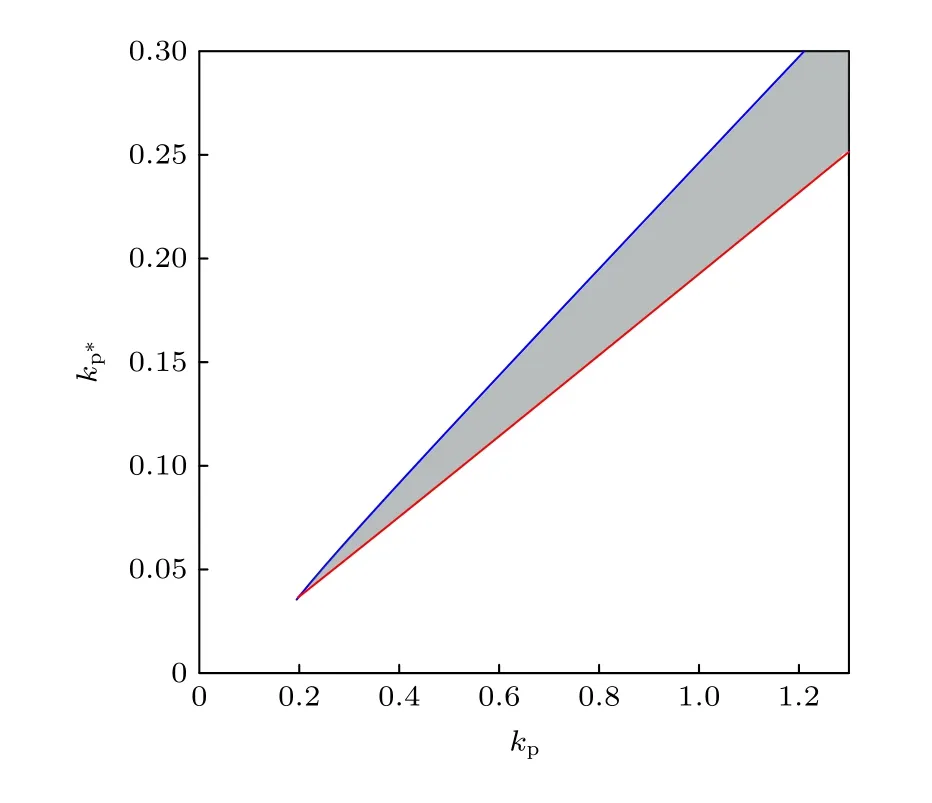
圖8 二維參數平面( kp , k p? )上的振蕩區域Fig.8.Oscillation area on the ( kp , k p? ) two-dimensional parameter plane.
4 結論與討論
總而言之, 本文全面探討了Mdm2對p53的正調控(即Mdm2p1促進p53的翻譯)和p53振蕩發生之間的關聯.首先利用生物事實給出一個p53基因網絡的數學模型; 然后利用數值分岔分析的方法, 研究了刻畫p53-Mdm2正反饋強度的參數kp: 1)kp與DSB的協作關系, 提出了實驗發現的p53對兩類能引起DSB的刺激(電離輻射、依托泊苷處理)出現不同動力學響應模式的原因可能是kp的不同; 2)kp與Wip1的協作關系, 指出了Wip1一方面在高強度正反饋下能促進振蕩, 另一方面在低強度正反饋下能抑制振蕩; 3)kp與Mdm2p2的協作關系, 再次證實了p53-Mdm2的負反饋回路是振蕩發生的結構基礎.本文強調的是正負反饋對p53基因網絡發生振蕩的同等重要性.值得注意的是, 希爾函數和微分方程刻畫的是宏觀層次的模型, 今后的研究還需建立更為細致的多態模型[34].p53的振蕩與細胞周期停滯或細胞凋亡等過程密不可分[35], 希望本研究能為今后的實驗設計和醫學應用提供思路.
此外, 本文的結果是在非線性程度(協同性)很高的調節關系上得出的, 這里的非線性程度很高主要體現在數值較大的希爾系數上.為了使結論更加可靠, 檢測希爾系數較小時振蕩發生的條件很有必要.把s0調到1;s1,s2和s3下調到2;s4,s6的
值不變 (因為p53以4聚體的形式綁定靶基因[36],其希爾系數理論上應取為4).為了使p53出現振蕩,k1的值改為0.09,k3的值改為0.05, 其他參數保持默認值, 在圖9中作出關于參數kp的分岔圖組.從圖9可以看出, 不同于強非線性的分岔現象是分岔點SN以及SNIC消失, 振蕩區間變為兩個HB之間的參數區間(HB1, HB2); 其他的振蕩控制現象未發生變化.也就是(kp,vatm)參數平面上的振蕩區域仍由兩條形分岔曲線所圍成, 當kp較小時p53能在ATM過度激活的情況下保持振蕩; (kp,vwip1) 參數平面上的振蕩區域由兩條形分岔曲線所圍成, 存在一部分kp的值使得Wip1抑制振蕩; (kp,kp?)參數平面上的振蕩區域由兩條“ / ”形直線所圍成, p53-Mdm2的正負反饋偶聯依然是這個基因網絡振蕩的“引擎”.因此, 本文的定性結論可能僅取決于這個基因網絡的結構,對希爾函數的非線性程度并不敏感.結論的可靠性得到了進一步的驗證.
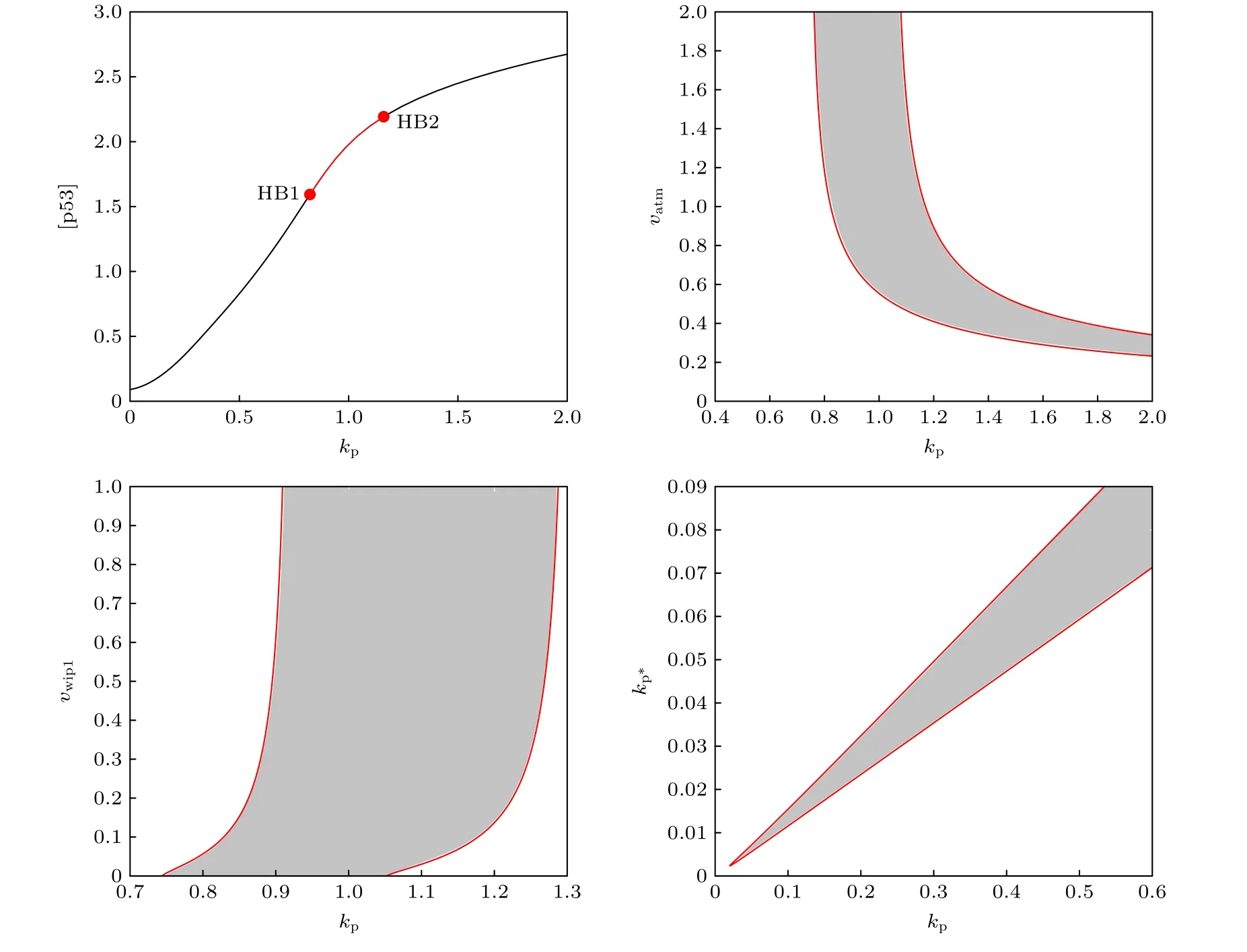
圖9 分岔圖組Fig.9.Bifurcation graph group.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
