茶樹CsMAPKK3基因克隆及表達(dá)分析
2021-08-02 19:19:32陸鯨冰楊潤梅張飛揚(yáng)曹紅利
南方農(nóng)業(yè)學(xué)報 2021年3期
陸鯨冰 楊潤梅 張飛揚(yáng) 曹紅利
摘要:【目的】克隆茶樹CsMAPKK3基因,并進(jìn)行表達(dá)分析,為探究CsMAPKK3基因的生物學(xué)功能及其抗逆分子機(jī)制提供理論支持。【方法】以鐵觀音為材料,采用RT-PCR克隆其CsMAPKK3基因的全長cDNA序列,對其進(jìn)行生物信息學(xué)分析,并采用實(shí)時熒光定量PCR(qRT-PCR)檢測其在不同組織及逆境脅迫下的表達(dá)情況。【結(jié)果】克隆獲得的CsMAPKK3基因(GenBank登錄號:AUD40506.1)cDNA序列全長1941 bp,包含完整開放閱讀框(ORF),長度為1557 bp,編碼518個氨基酸殘基,蛋白相對分子量為57.52 kD,理論等電點(diǎn)(pI)為5.39,為無跨膜螺旋結(jié)構(gòu)和信號肽的不穩(wěn)定蛋白,含有多個磷酸化位點(diǎn),定位于細(xì)胞質(zhì)和細(xì)胞核中,屬于PKc類型的MAPKK,且具有SPS1、S_TKc等催化結(jié)構(gòu)域,以及D(I/L/V)K激活域和S/T-X3-5-S/T保守域,與中華獼猴桃PSS35243.1親緣關(guān)系最近,且與擬南芥B亞家族聚為一類。CsMAPKK3基因起始密碼子上游1714 bp的啟動子區(qū)序列含有光響應(yīng)、逆境響應(yīng)、植物激素和厭氧誘導(dǎo)等相關(guān)的順式作用元件。CsMAPKK3基因在根、莖、葉、花和果中均有表達(dá),其中在莖和果的表達(dá)量較高,顯著高于在花中的表達(dá)量(P<0.05),但與根和葉中的表達(dá)量均無顯著差異(P>0.05)。低溫脅迫抑制CsMAPKK3基因表達(dá),而脫落酸(ABA)、高鹽和干旱脅迫均能誘導(dǎo)CsMAPKK3基因上調(diào)表達(dá)。【結(jié)論】CsMAPKK3基因表達(dá)具有組織表達(dá)特異性,且參與茶樹ABA、高鹽、低溫和干旱脅迫響應(yīng)等逆境脅迫的分子調(diào)控。
關(guān)鍵詞: 茶樹;CsMAPKK3基因;基因克隆;生物信息學(xué)分析;表達(dá)分析;逆境脅迫
中圖分類號: S571.103.53? ? ? ? ? ? ? ? ? ? ? ? ? ? ?文獻(xiàn)標(biāo)志碼: A 文章編號:2095-1191(2021)03-0651-09
Cloning and expression analysis of CsMAPKK3 gene in tea plant
LU Jing-bing, YANG Run-mei, ZHANG Fei-yang, CAO Hong-li*
(College of Horticulture,F(xiàn)ujian Agriculture and Forestry University/Key Laboratory of Tea Science in
Universities of Fujian Province, Fuzhou? 350002, China)
Abstract:【Objective】The CsMAPKK3 gene of tea tree was cloned and its expression was analyzed to provide theoretical support for clarifying the biological function of tea plant CsMAPKK3 gene and revealing its role in tea plants stress resistance. 【Method】In this study,the full-length cDNA of MAPKK3 gene were cloned using RT-PCR from Tieguanyin tea plant. Then,the bioinformatics characteristics and functions of CsMAPKK3 gene were predicted,and its expression patterns in various tissues and under different stress treatments were investigated using real-time fluorescence quantitative PCR. 【Result】The full-length cDNA of CsMAPKK3 was 1941 bp,with a 1557 bp open reading frame(ORF),encoding 518 amino acids,and submitted to the GenBank with accession number of? AUD40506.1. The molecular weight of CsMAPKK3 protein was 57.52 kD,and the theoretical isoelectric point(pI) was 5.39. The amino acid sequence prediction showed that it was an unstable protein without transmembrane helix structure and signal peptide,and contained multiple phosphorylation sites. CsMAPKK3 protein,located in cytoplasm and nucleus,belonged to PKC type MAPKK,and had catalytic domains such as SPS1,S_TKC,D(I/L/V)K activation domain and S/T-X3-5-S/T conserved domain. It had close relationship with Chinese kiwifruit(PSS35243.1),and was clustered in a class with Arabidopsis B subfamily in phylogenetic tree.? Additionally,promoter sequence analysis showed that the 1714 bp promoter region in the? initiation codon upstream of CsMAPKK3 contained several cis-acting elements related to light response,stress response,plant hormones,and anaerobic induction. Real time quantitative PCR analysis showed that CsMAPKK3 was expressed in roots,stems,leaves,flowers and fruits,and the expression level was higher in stems and fruits,which was significantly higher than that in flowers(P<0.05),but had no significant difference with that in roots and leaves(P>0.05). The expression of CsMAPKK3 was down-regulated by cold stress,but up-regulated by abscisic acid(ABA),salt and drought treatments. 【Conclusion】The expression of CsMAPKK3 is tissue-specific,and it is involved in the tea plant response to ABA,high salt,low temperature and drought stresses.
Key words: Camellia sinensis; CsMAPKK3 gene; gene cloning; bioinformatics analysis; expression analysis; stress
Foundation item: National Natural Science Foundation of China(31800587); Education Research Project for Young and Middle-aged Teachers of Education Department of Fujian(JAT170160)
0 引言
【研究意義】在逆境脅迫下,植物可通過促分裂原活化蛋白激酶(Mitogen-activated protein kinases,MAPKs)級聯(lián)途徑響應(yīng),經(jīng)過MAPKKK(MAPK kinase kinase)-MAPKK(MAPK kinase)-MAPK的依次磷酸化作用,將外界逆境信號級聯(lián)放大并傳遞至下游,調(diào)控相應(yīng)的生理生化反應(yīng)(Hamel et al.,2006)。目前,國內(nèi)外學(xué)者對MAPKs級聯(lián)途徑的研究越來越多,結(jié)果發(fā)現(xiàn)MAPKs級聯(lián)途徑在植物生長發(fā)育、抗病及非生物脅迫響應(yīng)、植物激素信號轉(zhuǎn)導(dǎo)通路和乙烯信號轉(zhuǎn)導(dǎo)過程中均發(fā)揮重要作用(趙琳琳等,2008)。茶樹喜好溫暖濕潤的生長環(huán)境,且偏好酸性土壤,低溫、干旱和高鹽等逆境條件下會導(dǎo)致茶樹生長發(fā)育不良甚至死亡,對成茶品質(zhì)產(chǎn)生嚴(yán)重影響。因此,研究茶樹MAPKs級聯(lián)途徑相關(guān)基因?qū)ζ淇鼓婢趁{迫的分子機(jī)制研究及抗逆育種具有重要意義。【前人研究進(jìn)展】MAPKK是雙特異性蛋白酶,在MAPKs級聯(lián)途徑中,上游被激活的MAPKKK能磷酸化MAPKK中保守的S/T-X3-5-S/T(S為絲氨酸,T為蘇氨酸,X為任意氨基酸)基序(Wu et al.,2014),從而激活MAPKK,然后MAPKK通過對MAPK活化環(huán)中T-X-Y(T為蘇氨酸,Y為酪氨酸)基序的磷酸化而激活MAPK,最終通過激活的MAPK來激活下游應(yīng)答因子將胞外逆境信號傳遞到細(xì)胞內(nèi)進(jìn)行響應(yīng)。植物MAPKK在MAPKs級聯(lián)途徑中的成員數(shù)量最少。據(jù)報道,擬南芥基因組中至少存在60個AtMAPKKK基因和20個AtMAPK基因,但僅10個AtMAPKK基因被鑒定出(張振才等,2014)。根據(jù)S/T-X3-5-S/T保守結(jié)構(gòu)和D激活位點(diǎn),AtMAPKK基因家族可區(qū)分為A、B、C和D 4個亞族,其中AtMAPKK1、AtMAPKK2和AtMAPKK6屬于A亞族,AtMAPKK3屬于B亞族,AtMAPKK4和AtMAPKK5屬于C亞族,AtMAPKK7、AtMAPKK8、AtMAPKK9和AtMAPKK10屬于D亞族(Hamel et al.,2006)。此外,MAPKK是上游逆境信號的聚集點(diǎn),也是下游MAPK的分支點(diǎn),因此,對MAPKK功能的揭示是明確MAPKs級聯(lián)反應(yīng)途徑作用機(jī)制的關(guān)鍵(Xu et al.,2010)。在逆境脅迫下,植物不同組織中的MAPKK基因表達(dá)量會發(fā)生上調(diào)或下調(diào)變化,然后通過MAPKs級聯(lián)途徑傳導(dǎo)脅迫信號,最終通過激素調(diào)控、滲透調(diào)節(jié)、氧化還原和信號轉(zhuǎn)導(dǎo)等對非生物脅迫作出應(yīng)答。擬南芥AtMKK1基因參與多種擬南芥響應(yīng)生物和非生物脅迫過程中的信號轉(zhuǎn)導(dǎo),包括對脫落酸(ABA)的響應(yīng)調(diào)控(蔡國華等,2013);將玉米ZmMKK1基因轉(zhuǎn)入擬南芥中,可提高轉(zhuǎn)基因植株的耐鹽性和抗旱性(Cai et al.,2014);棉花GhMKK3基因在響應(yīng)干旱脅迫下被誘導(dǎo)上調(diào)表達(dá),表明該基因參與干旱脅迫響應(yīng)調(diào)控(Wang et al.,2016)。【本研究切入點(diǎn)】目前鮮見有關(guān)茶樹MAPKK基因及MAPK信號通路在茶樹抗逆過程中作用機(jī)制的研究報道。【擬解決的關(guān)鍵問題】克隆茶樹CsMAPKK3基因cDNA全長序列,對其進(jìn)行生物信息學(xué)分析,并檢測該基因在外源ABA、鹽脅迫、干旱脅迫和低溫脅迫下的表達(dá)模式,并從茶樹(舒茶早)基因組數(shù)據(jù)庫中檢索該基因的啟動子序列,分析其順式作用元件,為深入研究茶樹MAPKK基因響應(yīng)逆境脅迫中的分子機(jī)制和調(diào)控機(jī)理提供理論參考。
1 材料與方法
1. 1 試驗材料
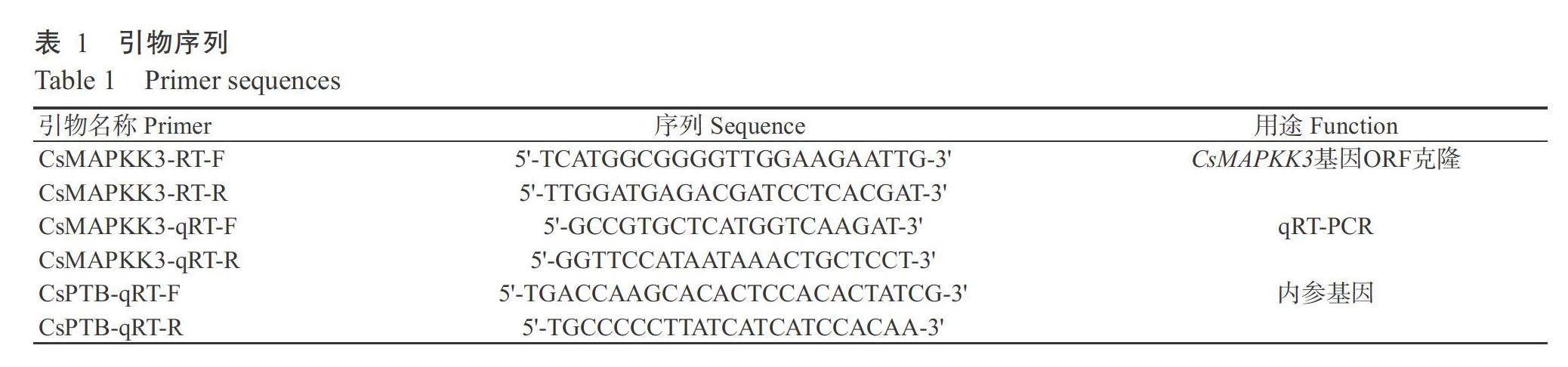
供試材料為3年生盆栽茶樹品種鐵觀音的茶苗。多糖多酚植物總RNA提取試劑盒購自杭州開泰生物有限公司;Easy Script @One-Step gDNA Removal and cDNA Synthesis SuperMix試劑盒和Transstart Tip Green qPCR SuperMix試劑盒購自廈門泰京生物技術(shù)有限公司。主要儀器設(shè)備:GeIDoc It TS3 Imager凝膠成像儀(Bio-Rad,美國)、CFX96 Touch熒光定量PCR儀(Bio-Rad,美國)、T100TM Thermal Cycler PCR儀(Bio-Rad,美國)、瓊脂糖水平電泳儀和電泳槽(北京六一儀器有限公司)、AIRTECH紫外超凈工作臺(蘇州安泰科技有限公司)和NanoDrop ND2000超微量分光光度計(Thermo Scientific,賽默飛世爾科技公司)。
1. 2 樣品處理及采集
取供試材料的根、莖、葉、花和果實(shí),每組織設(shè)3次生物學(xué)重復(fù)。參照曹紅利等(2017)的方法對材料進(jìn)行低溫(4 ℃)、ABA(100 μmol/L ABA)、高鹽(150 mmol/L NaCl)和干旱(100 g/L PEG)4種脅迫處理,分別在處理后3、9和24 h進(jìn)行取樣,以0 h為對照,每處理設(shè)3次生物學(xué)重復(fù)。所取樣品經(jīng)液氮速凍后放-80 ℃冰箱保存?zhèn)溆谩?/p>
1. 3 總RNA提取及cDNA合成
根據(jù)多糖多酚植物總RNA提取試劑盒說明提取總RNA,利用NanoDrop ND2000超微量分光光度計測定其濃度,并用1%瓊脂糖凝膠電泳檢測其完整性,置于-80 ℃冰箱保存?zhèn)溆谩0凑誆asy Script@One-Step gDNA Removal and cDNA Synthesis SuperMix試劑盒說明將所提取的總RNA反轉(zhuǎn)錄合成cDNA第一鏈,用于逆轉(zhuǎn)錄PCR(RT-PCR)和實(shí)時熒光定量PCR(qRT-PCR)檢測。
1. 4 基因克隆及啟動子序列分析
根據(jù)本課題組前期試驗獲得的茶樹低溫轉(zhuǎn)錄組數(shù)據(jù)庫中的基因注釋,篩選出1個MAPKK3基因序列,將該參考序列在NCBI中進(jìn)行比對后顯示其含有完整的開放閱讀框(ORF),因此設(shè)計ORF上、下游擴(kuò)增引物(表1),按照PrimeSTAR HS試劑盒的說明配制50.0 μL體系進(jìn)行PCR擴(kuò)增。擴(kuò)增程序:94 ℃預(yù)變性30 s,98 ℃ 10 s,55 ℃ 5 s,72 ℃ 90 s,共進(jìn)行30個循環(huán)。PCR產(chǎn)物回收測序后與原序列拼接,最終獲得該基因的cDNA序列。并根據(jù)cDNA序列在舒茶早茶樹基因組數(shù)據(jù)庫中檢索,獲得其密碼子上游序列2000 bp的序列,最終得到1714 bp啟動子序列。
1. 5 生物信息學(xué)分析
利用NCBI數(shù)據(jù)庫中的BLASTn和BLASTp對CsMAPKK3基因的核苷酸序列及其編碼的氨基酸序列進(jìn)行同源性分析,并利用DNAMAN 6.0將CsMAPKK3與同源蛋白序列進(jìn)行多重比對。使用ExPASy在線工具中的ProtParam預(yù)測CsMAPKK3蛋白的理化性質(zhì);使用SingalP 3.0預(yù)測其信號肽;利用TMHMM 2.0預(yù)測其跨膜螺旋結(jié)構(gòu)域;采用NetPhos 3.1 Server預(yù)測其磷酸化位點(diǎn);利用Plant-mPLoc和WOLF PSORT預(yù)測其亞細(xì)胞定位。將獲得的CsMAPKK3基因啟動子序列提交至PlantCARE中對其逆境相關(guān)順式作用元件進(jìn)行預(yù)測。利用MEGA 7.0的鄰接法(Neighbor-joining method)構(gòu)建系統(tǒng)發(fā)育進(jìn)化樹。
1. 6 qRT-PCR檢測
通過qRT-PCR檢測CsMAPKK3基因在茶樹不同組織及4種脅迫處理下的表達(dá)情況,以CsPTB為內(nèi)參基因,定量PCR引物見表1。參照Transstart Tip Green qPCR SuperMix試劑盒說明配制反應(yīng)體系(20.0 μL),包括2×TransTaq HiFiPCR SuperMix 10.0 μL,cDNA模板1.0 μL,10 μmol/L上、下游引物各0.5 μL,ddH2O補(bǔ)足至20.0 μL。擴(kuò)增程序:94 ℃預(yù)變性30 s;94 ℃ 5 s;60 ℃ 30 s,共進(jìn)行40個循環(huán)。采用2-ΔCt算法計算目的基因在組織中的相對表達(dá)量,2-ΔΔCt算法計算其在非生物脅迫下的相對表達(dá)量。
1. 7 統(tǒng)計分析
使用SPSS 22.0對數(shù)據(jù)采用LSD法和Dunnett法進(jìn)行顯著性分析(P<0.05),使用Excel 2020繪制柱形圖。
2 結(jié)果與分析
2. 1 CsMAPKK3基因克隆及序列分析結(jié)果
如圖1所示,CsMAPKK3基因ORF的PCR產(chǎn)物約1500 bp,與預(yù)期結(jié)果相符,其測序結(jié)果顯示,該序列為1557 bp,與參考序列拼接后,獲得cDNA全長為1941 bp。CsMAPKK3基因編碼518個氨基酸殘基,且氨基酸序列中含有D(I/L/V)K激活域以及S/T-X3-5-S/T保守域(圖2)。該基因序列已提交至GenBank,登錄號為AUD40506.1。
2. 2 生物信息學(xué)分析結(jié)果
ProtParam預(yù)測結(jié)果顯示,CsMAPKK3蛋白分子式為C2566H3996N682O768S26,相對分子量為57.52 kD,含酸性氨基酸(Asp+Glu)63個、堿性氨基酸(Arg+Lys)48個,理論等電點(diǎn)(pI)為5.39,不穩(wěn)定系數(shù)為42.07,屬于不穩(wěn)定蛋白。TMHMM 2.0預(yù)測結(jié)果(圖3)顯示,CsMAPKK3蛋白無跨膜螺旋結(jié)構(gòu)和信號肽。SignalP 3.0信號肽預(yù)測結(jié)果與TMHMM預(yù)測結(jié)果相同,即該蛋白無信號肽。
NetPhos 3.1 Server預(yù)測結(jié)果(圖4)顯示,CsMAPKK3蛋白含有52個能被磷酸化的位點(diǎn)(超過閾值線),其中包括27個Ser位點(diǎn)、16個Thr和9個Tyr位點(diǎn),主要被unsp、cdc2、CKII和INSR等蛋白激酶磷酸化。Plant-mPLoc預(yù)測結(jié)果顯示,CsMAPKK3蛋白定位于細(xì)胞核;WOLF PSORT預(yù)測結(jié)果顯示,其定位于細(xì)胞質(zhì)和細(xì)胞核(K值分別為11和2),故推測CsMAPKK3蛋白的亞細(xì)胞定位可能定位于細(xì)胞質(zhì)和細(xì)胞核中。利用NCBI中BLASTp對CsMAPKK3蛋白功能結(jié)構(gòu)域進(jìn)行分析,結(jié)果(圖5)發(fā)現(xiàn),CsMAPKK3蛋白屬于PKc類型的MAPKK,且具有SPS1、S_TKc等催化結(jié)構(gòu)域。
用NCBI數(shù)據(jù)庫中BLASTn對CsMAPKK3基因進(jìn)行同源比對,結(jié)果顯示該基因與葡萄(Vitis vini-fera)MAPKK3基因(GenBank登錄號MT154083.1)的相似性最高,達(dá)85%,與栓皮櫟(Quercus suber)MAPKK3基因(GenBank登錄號XM_024032986.1)和巴西橡膠樹(Hevea brasiliensis)MAPKK3基因(GenBank登錄號XM_021788394.1)的相似性均達(dá)84%。從NCBI數(shù)據(jù)庫下載CsMAPKK3的同源蛋白氨基酸序列,并利用DNAMAN 6.0進(jìn)行多重序列比對,結(jié)果(圖6)顯示,CsMAPKK3、擬南芥AtMKK3、玉米ZmMKK3和中華獼猴桃(Actinidia chinensis var. chinensis)PSS35243.1序列中均含D(I/L/V)K激活域和S/T-X3-5-S/T保守域。以CsMAPKK3蛋白與擬南芥的10個AtMAPKK蛋白構(gòu)建系統(tǒng)發(fā)育進(jìn)化樹,結(jié)果(圖7)顯示,CsMAPKK3蛋白與擬南芥B亞家族聚為一類,其中,與AtMKK3蛋白關(guān)系最近。以CsMAPKK3蛋白與其他17種植物的MAPKK蛋白構(gòu)建系統(tǒng)發(fā)育進(jìn)化樹,結(jié)果(圖8)表明,CsMAPKK3蛋白與中華獼猴桃PSS35243.1親緣關(guān)系最近。
2. 3 CsMAPKK3基因啟動子順式作用元件分析結(jié)果
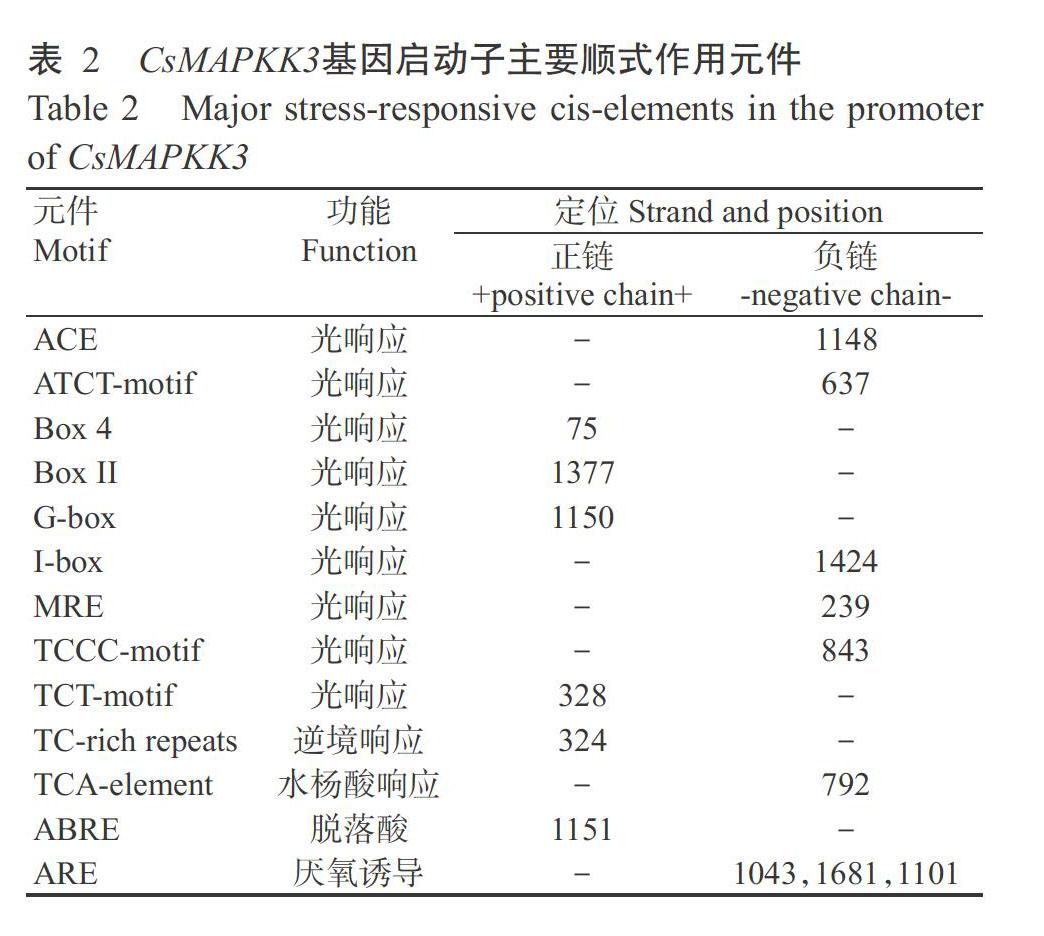
在PlantCARE中對茶樹CsMAPKK3基因啟動子序列進(jìn)行順式作用元件預(yù)測分析,結(jié)果(表2)顯示,啟動子區(qū)含有與光響應(yīng)相關(guān)元件如ACE、ATCT-motif、Box 4、MRE和TCCC-motif等,以及多種逆境響應(yīng)相關(guān)元件如TC-rich repeats等。此外,還含有與植物激素響應(yīng)相關(guān)元件如脫落酸響應(yīng)元件ABRE、水楊酸響應(yīng)元件TCA-element等,以及與厭氧誘導(dǎo)相關(guān)元件如ARE等。
2. 4 CsMAPKK3基因在不同組織及逆境脅迫處理下表達(dá)模式分析結(jié)果
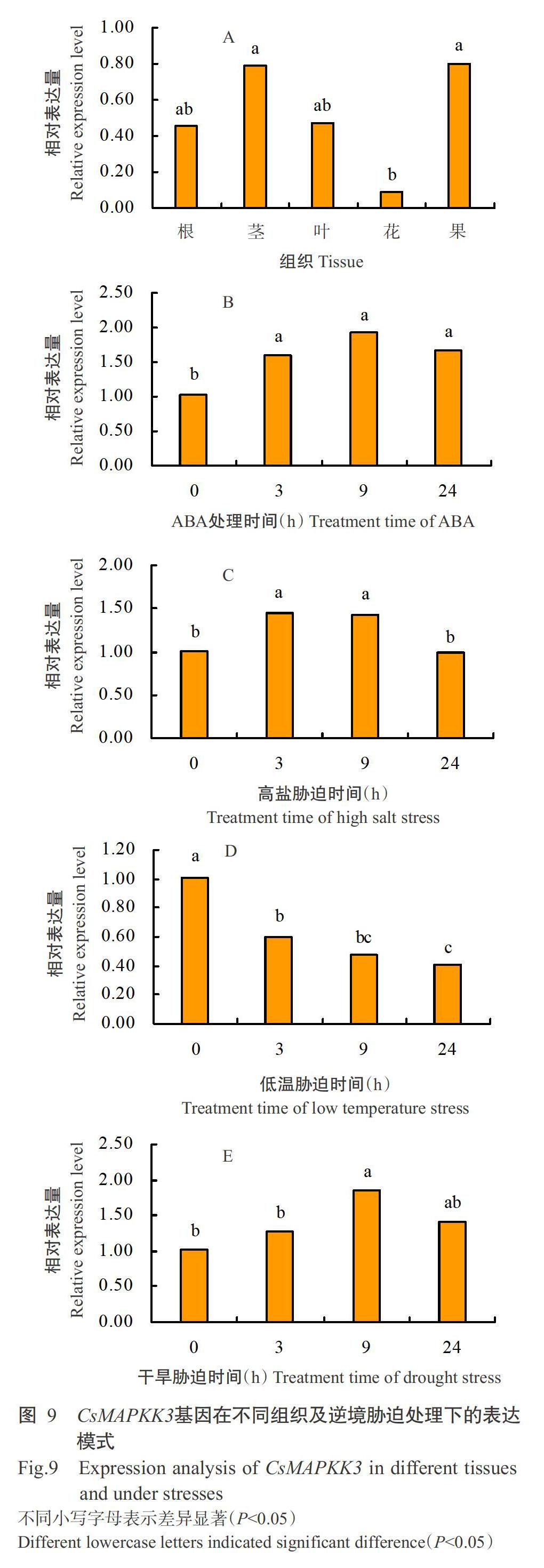
CsMAPKK3基因在根、莖、葉、花和果中均有表達(dá),其中在莖和果的表達(dá)量較高,顯著高于花中的表達(dá)量,但與根和葉中的表達(dá)量均無顯著差異(P>0.05)(圖9-A)。CsMAPKK3基因在4種逆境脅迫處理(ABA、高鹽、低溫和干旱)下的表達(dá)量檢測結(jié)果如圖9-B~圖9-E所示。ABA處理后CsMAPKK3基因較處理0時顯著上調(diào)表達(dá)(P<0.05,下同),在9 h時達(dá)最大值(圖9-B);高鹽脅迫處理也能顯著誘導(dǎo)CsMAPKK3基因顯著上調(diào)表達(dá),在3 h時達(dá)最大值(圖9-C);低溫脅迫處理下CsMAPKK3基因的表達(dá)模式與其他3個處理完全相反,其表達(dá)量呈顯著下調(diào)趨勢,在24 h時達(dá)最小值(圖9-D);干旱脅迫處理下CsMAPKK3基因的表達(dá)量與ABA處理呈相似的變化趨勢,在9 h時顯著上調(diào)表達(dá),表達(dá)量達(dá)最大值(圖9-E)。推測CsMAPKK3基因參與茶樹ABA、高鹽、低溫和干旱脅迫響應(yīng)調(diào)控。
3 討論
自Ichimura等(1998)首次從模式植物擬南芥中鑒定出1條完整的MAPKs級聯(lián)途徑AtMEKK1-AtMKK1-AtMPK4,并證明其能傳遞干旱和機(jī)械損傷信號后,Teige等(2004)也在擬南芥中發(fā)現(xiàn)另一條MAPKs級聯(lián)途徑AtMEKK1-AtMAPKK2-AtMAPK4/AtMAPK6,并證實(shí)其參與抵御鹽脅迫和低溫脅迫;Schrnidt等(2013)研究發(fā)現(xiàn)MAPKs級聯(lián)途徑OsMEKK6-OsMAPK5參與水稻根部對鹽脅迫的信號轉(zhuǎn)導(dǎo)。可見,MAPKs級聯(lián)途徑在響應(yīng)非生物脅迫中發(fā)揮重要作用,而MAPKK作為連接上游MAPKKK和下游MAPK的信號樞紐,是MAPKs級聯(lián)途徑中的關(guān)鍵點(diǎn)(單鴻軒和付暢,2017)。本研究克隆獲得CsMAPKK3基因,并進(jìn)行組織表達(dá)特性及非生物脅迫下表達(dá)模式分析,以期為研究MAPKs級聯(lián)途徑在茶樹中的抗逆作用機(jī)制提供參考。CsMAPKK3亞細(xì)胞定位結(jié)果顯示其可能定位于細(xì)胞質(zhì)和細(xì)胞核中,與玉米ZmMKK3蛋白定位在細(xì)胞質(zhì)和細(xì)胞核中的預(yù)測結(jié)果一致(Zhang et al.,2012)。而在此前與MAPKK定位相關(guān)的研究中,發(fā)現(xiàn)大多MAPKK蛋白定位于細(xì)胞質(zhì)(Song et al.,2015),但也有定位于細(xì)胞核(Song et al.,2015)的報道,可能是由于MAPKK在細(xì)胞質(zhì)中被上游MAPKKK激活,隨后轉(zhuǎn)移至細(xì)胞核磷酸化其下游的MAPK所導(dǎo)致。
本研究發(fā)現(xiàn),CsMAPKK3基因啟動子序列中含有與響應(yīng)ABA脅迫和水楊酸等激素信號分子相關(guān)的多種順式作用元件,推測該基因參與激素信號分子調(diào)控,從而影響植物生長發(fā)育。且通過qRT-PCR檢測發(fā)現(xiàn),CsMAPKK3基因在茶樹莖和果中的表達(dá)量較高,與黃瓜CsMAPK4-2基因在果實(shí)中表達(dá)量最高,其旁系同源基因CsMAPK4-1在莖中的表達(dá)量較高的結(jié)果相似(Wang et al.,2015);CsMAPKK3基因受高鹽和干旱脅迫誘導(dǎo)表達(dá)顯著上調(diào),推測該基因可能參與茶樹耐鹽和抗旱響應(yīng)。李粲等(2012)克隆獲得甘蔗SoMAPK4基因,并證實(shí)該基因可能參與多種生物和非生物脅迫響應(yīng)的信號傳導(dǎo)。Song等(2015)研究發(fā)現(xiàn),西瓜ClMKK3基因受非生物(干旱、高鹽、高溫和ABA)脅迫不同程度誘導(dǎo)上調(diào)表達(dá),但低溫脅迫抑制其顯著下調(diào)表達(dá),表明該基因在逆境脅迫中發(fā)揮重要調(diào)控功能;ClMKK5基因在西瓜枯萎菌侵染后第9 d時的表達(dá)量是對照的6倍,表明該基因在西瓜抗枯萎病過程中發(fā)揮重要調(diào)控功能(Song et al.,2015)。張演義等(2015)對葡萄MAPKK基因家族進(jìn)行鑒定分析,結(jié)果發(fā)現(xiàn)MAPKK基因家族對植物生長發(fā)育細(xì)胞周期調(diào)控以及響應(yīng)環(huán)境脅迫方面有重要作用。
基于前人對植物抗旱耐鹽基因的研究結(jié)果,發(fā)現(xiàn)這些基因主要包括滲透調(diào)節(jié)基因、蛋白類基因(如信號傳導(dǎo)中的蛋白激酶基因)及轉(zhuǎn)錄因子基因(陳麗萍和何道一,2010)。CsMAPKK3基因則屬于蛋白類基因。蛋白磷酸化及去磷酸化過程在細(xì)胞的信號識別和轉(zhuǎn)導(dǎo)中發(fā)揮重要作用,而細(xì)胞信號識別和轉(zhuǎn)導(dǎo)又直接影響植物體對環(huán)境變化的感應(yīng)和對逆境信息的傳遞,故在這個過程中蛋白激酶發(fā)揮重要的作用。通過蛋白質(zhì)組學(xué)研究發(fā)現(xiàn),在逆境條件下許多與逆境相關(guān)的蛋白表達(dá)量會發(fā)生明顯變化(付晨熙等,2016)。因此,后續(xù)應(yīng)重點(diǎn)研究CsMAPKK3蛋白在逆境中的調(diào)控機(jī)制及CsMAPKK3基因在MAPKs級聯(lián)途徑中的作用分子機(jī)制。
4 結(jié)論
CsMAPKK3基因表達(dá)具有組織表達(dá)特異性,且參與茶樹ABA、高鹽、低溫和干旱脅迫響應(yīng)等逆境脅迫的分子調(diào)控。
參考文獻(xiàn):
蔡國華,王麗,潘教文,孔祥培,李德全. 2013. 擬南芥AtMKK1結(jié)構(gòu)特征和信號轉(zhuǎn)導(dǎo)功能研究進(jìn)展[J]. 植物生理學(xué)報,49(3):225-233. doi:10.13592/j.cnki.ppj.2013.03.012. [Cai G H,Wang L,Pan J W,Kong X P,Li D Q. 2013. Research progress of structure characteristics and signal transduction functions of AtMKK1 in Arabidopsis[J]. Plant Physiology Journal,49(3):225-233.]
曹紅利,陳丹,葉乃興,郭雅玲,岳川. 2017. 茶樹CsMAPK3的全長克隆及其逆境表達(dá)分析[J]. 園藝學(xué)報,44(11):2203-2214. doi:10.16420/j.issn.0513-353x.2017-0367. [Cao H L,Chen D,Ye N X,Guo Y L,Yue C. 2017. Clo-ning and abiotic stress expression analysis of CsMAPK3 gene in tea plant[J]. Acta Horticulturae Sinica,44(11):2203-2214.]
陳麗萍,何道一. 2010. 植物抗旱耐鹽基因的研究進(jìn)展[J]. 基因組學(xué)與應(yīng)用生物學(xué),29(3):542-549. doi:10.3969/gab. 029.000542. [Chen L P,He D Y. 2010. Research advance on drought and salt resistant genes in transgenic plants[J]. Genomics and Applied Biology,29(3):542-549.]
付晨熙,肖自華,高飛,周宜君. 2016. 植物應(yīng)答非生物脅迫的蛋白質(zhì)組學(xué)研究進(jìn)展[J]. 基因組學(xué)與應(yīng)用生物學(xué),35(12):3569-3582. doi:10.13417/j.gab.035.003569. [Fu C X,Xiao Z H,Gao F,Zhou Y J. 2016. Progress on proteomics to reveal abiotic stress response in plant[J]. Genomics and Applied Biology,35(12):3569-3582.]
李粲,滕崢,劉開雨,桑洪玉,盧雙楠,方位寬,梁朝旭,劉曉靜,何珊珊,劉芳,邱永福,李鳴,李容柏. 2012. 甘蔗MAP激酶基因SoMAPK4克隆與生物信息學(xué)分析[J]. 南方農(nóng)業(yè)學(xué)報,43(6):727-732. doi:10.3969/j:issn.2095-1191. 2012.06.727. [Li C,Teng Z,Liu K Y,Sang H Y,Lu S N,F(xiàn)ang W K,Liang C X,Liu X J,He S S,Liu F,Qiu Y F,Li M,Li R B. 2012. Cloning and bioinformatic analysis of mitogen-activated protein kinase gene SoMAPK4 in sugarcane[J]. Journal of Southern Agriculture,43(6):727-732.]
單鴻軒,付暢. 2017. 逆境脅迫下植物MAPK級聯(lián)反應(yīng)途徑研究新進(jìn)展[J]. 核農(nóng)學(xué)報,31(4):680-688. doi:10.11869/j.issn.100-8551.2017.04.0680. [Shan H X,F(xiàn)u C. 2017. The recent advances on plant MAPK cascade pathway under adverse stress conditions[J]. Journal of Nuclear Agriculture Sciences,31(4):680-688.]
張演義,呂福堂,張全軍,房經(jīng)貴. 2015. 葡萄MAPKK基因家族的識別與分析[J]. 西南農(nóng)業(yè)學(xué)報,28(4):1791-1797. doi:10.16213/j.cnki.scjas.2015.04.070. [Zhang Y Y,Lü F T,Zhang Q J,F(xiàn)ang J G. 2015. Identification and analysis of MAPKK gene family in Vitis vinifera[J]. Southwest China Journal of Agricultural Sciences,28(4):1791-1797.]
張振才,梁燕,李翠. 2014. 植物MAPK級聯(lián)途徑及其功能研究進(jìn)展[J]. 西北農(nóng)林科技大學(xué)學(xué)報(自然科學(xué)版),42(4):207-214,doi:10.13207/j.cnki.jnwafu.2014.04.013. [Zhang Z C,Liang Y,Li C. 2014. Review on plant MAPK cascades and their functions[J]. Journal of Northwest A & F University(Natural Science Edition),42(4):207-214.]
趙琳琳,徐啟江,李玉花. 2008. 生物和非生物脅迫下的植物細(xì)胞中絲裂原活化蛋白激酶(MAPK)信號轉(zhuǎn)導(dǎo)[J]. 植物生理學(xué)通訊,44(1):169-174. doi:10.13592/j.cnki.ppj. 2008.01.017. [Zhao L L,Xu Q J,Li Y H. 2008. The mitogen-activated protein kinase signal transduction in plant cell under biotic and abiotic stress conditions[J]. Plant Physiology Communications,44(1):169-174.]
Cai G H,Wang G D,Wang L,Liu Y,Pan J W,Li D Q. 2014. A maize mitogen-activated protein kinase kinase,Zm-MKK1,positively regulated the salt and drought tolerance in transgenic Arabidopsis[J]. Journal of Plant Physiology,171(12):1003-1016. doi:10.1016/j.jplph.2014.02.012.
Hamel L P,Nicole M C,Sritubtim S,Morency M J,Ellis M,Ehlting J,Beaudoin N,Barbazuk B,Klessig D,Lee J,Martin G,Mundy J,Ohashi Y,Scheel D,Sheen J,Xing T,Zhang S Q,Seguin A,Ellis B E. 2006. Ancient signals:Comparative genomics of plant MAPK and MAPKK gene families[J]. Trends in Plant Science,11(4):192-198. doi:10.1016/j.tplants.2006.02.007.
Ichimura K,Mizpguchi T,Irie K,Morris P,Giraudat J,Matsumoto K,Shinozaki K. 1998. Isolation of ATMEKK1(a MAP kinase kinase kinase):Interacting proteins and analy-sis of a MAP kinase cascade in Arabidopsis[J]. Biochemi-cal & Biophysical Research Communications,253(2):532-543. doi:10.1006/bbrc.1998.9796.
Schrnidt R,Mieulet D,Hubberten H M,Obata T,Hoefgen R,Alisdair R,F(xiàn)ernie,F(xiàn)isahn J,Segundo B S,Guiderdoni E,Schippers J,Bernd M R 2013. Salt-responsive ERF1 regu-lates reactive oxygen species-dependent signaling during the initial response to salt stress in rice[J]. The Plant Cell,25(6):2115-2131. doi:10.1105/TPC.113.113068.
Song Q M,Li D Y,Dai Y,Liu S X,Huang L,Hong Y B,Zhang H J,Song F M. 2015. Characterization,expression patterns and functional analisis of the MAPK and MAPKK genes in watermelon(Citrullus lanatus)[J]. BMC Plant Biology,15(1):298-321. doi:10.1186/s12870-015-0681-4.
Teige M,Scheikl E,Eulgem T,Doczi R,Ichimura K,Shinozaki K,Shinozaki K,Jeffery L,Dangl,Hirt H. 2004. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis[J]. Molecular Cell,15(1):141-152. doi:10. 1016/j.molcel.2004.06.023.
Wang C,Lu W J,He X W,Wang F,Zhou Y L,Guo X L,Guo X Q. 2016. The cotton Mitogen-Activated Protein Kinase Kinase 3 functions in drought tolerance by regulating stomatal responses and root growth[J]. Plant Cell Physiology,57(8):1629-1642. doi:10.1093/pcp/pcw090.
Wang J,Pan C T,Wang Y,Ye L,Wu J,Chen L F,Zou T,Lu G. 2015. Genome-wide identification of MAPK,MAPKK,and MAPKKK gene families and transcriptional profiling analysis during development and stress response in cucumber[J]. BMC Genomics,16(1):1-22. doi:10.1186/s12864-015-1621-2.
Wu J,Wang J,Pan C T,Guan X Y,Wang Y,Liu S Y,He Y J,Chen J L,Chen L F,Lu G. 2014. Genome-wide identification of MAPKK and MAPKKK gene families in tomato and transcriptional profiling analysis during development and stress response[J]. PLoS One,9(7):103032. doi:10. 1371/journal.pone.0103032.
Xu H N,Li K Z,Yang F J,Shi Q H,Wang X F. 2010. Overexpression of CsNMAPK in tobacco enhanced seed germination under salt and osmotic stresses[J]. Molecular Biology Reports,37(7):3157-3163. doi:10.1007/s11033-009-9895-6.
Zhang M Y,Pan J W,Kong X P,Zhou Y,Liu Y,Sun L P,Li D Q. 2012. ZmMKK3,a novel maize group B mitogen-activated protein kinase kinase gene,mediates osmotic stress and ABA signal responses[J]. Journal of Plant Phy-siology,169(15):1501-151. doi:10.1016/j.jplph.2012.06. 008.
(責(zé)任編輯 陳 燕)
