miR-191靶向BDNF基因通過激活PI3K/AKT信號通路促進豬未成熟支持細胞增殖
2021-07-28 06:51:24唐湘薇楚丹顏賽娜尹艷飛卞橋翁波陳斌冉茂良
遺傳 2021年7期
唐湘薇,楚丹,顏賽娜,尹艷飛,卞橋,翁波,陳斌,冉茂良
研究報告
miR-191靶向基因通過激活PI3K/AKT信號通路促進豬未成熟支持細胞增殖
唐湘薇,楚丹,顏賽娜,尹艷飛,卞橋,翁波,陳斌,冉茂良
湖南農業大學動物科學技術學院,畜禽遺傳改良湖南省重點實驗室,長沙 410128
睪丸支持細胞數量是影響精子生成能力的主要因素之一,microRNA (miRNA)參與調控豬未成熟支持細胞的發育過程,然而,大多數被鑒定出的miRNA對支持細胞的作用及其機制尚不明確。基于本課題組前期高內涵篩選結果,本文進一步通過流式細胞術、蛋白免疫印跡和雙熒光素酶報告基因等方法,研究了調控豬未成熟支持細胞增殖和凋亡的作用機理。結果表明:過表達顯著促進細胞周期由G1期進入S期和G2期,細胞增殖能力顯著增強,細胞凋亡率顯著降低;而抑制表達則與之相反。雙熒光素酶報告基因系統驗證直接靶向基因3′-UTR。抑制表達基因促進細胞周期進入S期,并促進細胞增殖而抑制細胞凋亡,與過表達的作用一致。共轉染試驗結果顯示,基因可以拮抗對細胞增殖和凋亡的調控作用。此外,過表達和抑制表達基因均可顯著促進PI3K/AKT信號通路中關鍵蛋白PI3K和AKT的磷酸化水平,且基因同樣拮抗對PI3K和AKT蛋白的調控作用。本研究結果證實靶向基因,通過激活PI3K/AKT信號通路促進豬未成熟支持細胞增殖且抑制其凋亡,為進一步解析調控豬精子生成的生物學功能提供了理論基礎。
;基因;PI3K/AKT信號通路;增殖;豬睪丸支持細胞
睪丸支持細胞作為生精小管內一種體細胞,不僅通過細胞間的緊密連接形成血睪屏障,為精子細胞的生長發育提供物理支撐和穩定的微環境,還分泌多種細胞因子以保障和促進精子細胞的成熟。豬睪丸支持細胞增殖活性的高峰期在出生至1月齡與初情期前后兩個階段,隨后睪丸支持細胞增殖活性逐漸喪失,公豬性成熟后睪丸組織中支持細胞數量保持相對恒定[1]。然而,每個成熟的支持細胞僅能支撐30~50個精子細胞的生長發育,因此支持細胞數量與成年雄性動物睪丸組織大小、精子密度、精子活力、生精小管中生殖細胞數量和間質細胞數量息息相關[2,3]。這表明豬睪丸支持細胞的增殖活性決定著性成熟后睪丸支持細胞數量,進而影響公豬整個使用年限內的精子發育和成熟以及精液品質。
MicroRNA (miRNA)作為一類長約22 nt的非編碼RNA,通過靶向蛋白編碼基因3′-非翻譯區(3′-un-translated region, 3′-UTR)抑制其翻譯過程,從而廣泛參與調控細胞增殖、凋亡、分化等多種生物學過程。近年來,利用RNA-seq技術已從不同發育階段的豬睪丸組織中鑒定出300余個miRNA[4~6],且經細胞分離后,相比于生精細胞,18個miRNA高表達于支持細胞[7]。在此基礎上,多個miRNA已被證實在豬睪丸支持細胞的生長發育過程中具有重要調控作用。例如:[8]、[9]和[10]等調控豬睪丸支持細胞增殖;[11]、[12]和[13]等調控豬睪丸支持細胞氧化應激水平;調控豬睪丸支持細胞自噬活性[14]。本課題組前期采用高內涵篩選技術對60個miRNA調控豬未成熟支持細胞增殖的作用進行了篩選,鑒定出包括在內的27個miRNA對豬未成熟支持細胞具有較強的促增殖效應[15],但的作用機制尚不清楚。研究表明,可以促進多種細胞的增殖,例如肝癌細胞[16]、食管鱗狀細胞[17]、成纖維細胞[18]等。基于miRNA序列在物種間的高度保守性,我們推測對豬未成熟支持細胞增殖具有重要的調控作用。因此,本研究利用流式細胞術、CCK-8 (cell counting kit-8)、EdU (5-ethynyl-2′-deoxyuridine)、實時熒光定量PCR (quantitative real time PCR, qRT-PCR)、Western blotting和雙熒光素酶報告基因等技術解析了調控豬未成熟支持細胞增殖和凋亡的靶基因及信號機制,為進一步闡明調控豬睪丸發育和精子生成的機制奠定理論基礎。
1 材料與方法
1.1 細胞培養
豬睪丸細胞系(swine testicular, ST) ATCC?CRL- 1746TM和293T工具細胞購自上海安為生物科技有限公司,其中ST已被證實為豬未成熟支持細胞[19],且廣泛應用于相關研究[8,15]。細胞培養基為DMEM高糖培養基(美國Gibco公司):胎牛血清(美國Gibco公司):雙抗(Penicillin-Streptomycin,美國Gibco公司)= 10∶1∶0.11,并將細胞置于37℃、5%CO2的培養箱中培養。
1.2 細胞轉染
設計合成模擬物(mimic)和抑制劑(inhibitor)序列過表達和抑制表達,設計合成基因siRNA (5′-GCCAACTGAAGCAGTACTT- 3′)抑制表達基因。接種細胞于6孔板,在融合度達到60%~70%時,吸棄培養基,每孔加入1 mL PBS (美國Gibco公司)洗滌,重復操作2次,吸棄PBS。向DMEM高糖基礎培養基(每孔250 μL)中加入Lipofectamine 2000試劑(每孔5 μL,美國Invitrogen公司),充分混勻成試劑①,室溫靜置5 min;再向DMEM 高糖基礎培養基(每孔250 μL)中分別加入mimic、mimic NC、inhibitor、inhibitor NC、siRNA、siRNA NC、inhibitor +siRNA、inhibitor + siRNA NC和inhibitor NC + siRNA NC (每孔5 μL,廣州銳博生物科技有限公司),充分混勻成試劑②,室溫靜置5 min;將試劑①與試劑②混勻,靜置30 min后加入細胞液中。每組至少設置3個重復,轉染6~8 h后更換為完全培養基繼續培養,以備后續實驗使用。
1.3 流式細胞周期檢測
細胞轉染24 h后,胰酶(美國HyClone公司)消化并收集細胞于1.5 mL EP管,每管加入250 μL PBS重懸,將細胞懸液逐滴加入750 μL預冷乙醇中,4℃過夜保存,1000 r/min離心5 min,棄上清,每管加入1 mL預冷的PBS,重懸細胞,1000 r/min離心5 min,棄PBS,每管加入150 μL濃度為100 μg/mL的碘化丙啶(propidium iodide,PI)染色液(長沙維爾生物科技有限公司),在4℃避光條件下染色30 min。使用流式細胞儀(FACSCalibur,美國BD公司)檢測細胞周期分布情況。
1.4 細胞增殖檢測
接種細胞于96孔板,采用CCK-8(東仁化學科技(上海)有限公司)和EdU試劑盒(廣州銳博生物科技有限公司)檢測細胞增殖情況。CCK-8檢測方法:分別轉染24 h和48 h后,每孔加入10 μL CCK-8試劑,繼續置于37℃、5%CO2條件下孵育1 h,使用酶標儀于450 nm波長下檢測每孔細胞的吸光度值,將24 h作為對照,計算48 h的相對增殖率。EdU檢測方法:轉染24 h后,每孔加入100 μL EdU培養基,繼續置于37℃、5%CO2條件下孵育2 h,并嚴格按照EdU試劑盒使用說明書進行細胞固定、Apollo染色和DNA染色,置于熒光顯微鏡(德國ZEISS公司)下拍照,并使用Image J軟件對圖片中的所有活細胞和有絲分裂細胞計數。
1.5 流式細胞凋亡檢測
接種細胞于6孔板,采用Annexin V-FITC/PI染色法(南京凱基生物科技發展有限公司)于流式細胞儀上檢測細胞凋亡情況。流式細胞凋亡檢測:轉染24 h后,胰酶消化并收集細胞,每管加100 μL 1× Binding Buffer (上海碧云天生物技術有限公司)重懸洗滌,加入5 μL Annexin V-APC試劑孵育15 min,加5 μL PI染色液重懸細胞,置于4℃避光,使用流式細胞儀檢測細胞凋亡情況。
1.6 細胞ATP水平檢測
接種細胞于6孔板,轉染24 h后,棄去培養基,PBS清洗2次,每孔加入200 μL ATP裂解液(上海碧云天生物技術有限公司),靜置裂解5 min,收集細胞于1.5 mL EP管,4℃、15,000 r/min離心5 min,使用ATP試劑盒(上海碧云天生物技術有限公司)按照使用說明檢測細胞ATP水平。
1.7 Western blotting檢測
收集細胞于1.5 mL EP管,采用RIPA lysis buffer (上海碧云天生物技術有限公司)提取細胞總蛋白,采用BCA蛋白定量試劑盒(上海碧云天生物技術有限公司)在酶標儀上對總蛋白進行定量。每孔加10 μL煮沸的蛋白樣品于10%SDS-PAGE的分離膠和5%SDS-PAGE的濃縮膠(上海碧云天生物技術有限公司)中,先60~80 V、30 min,然后調整為100~ 120 V、60 min進行電泳,觀察溴酚藍至膠板底部即可停止。使用電流300 mA、40 min條件進行轉膜,隨后加入含5%脫脂奶粉的TBST封閉液(北京索萊寶科技有限公司),搖床室溫封閉條帶1.5 h,加入一抗于4℃孵育過夜。一抗包括:Bcl2 (1∶1000,美國Protein Tech Group公司)、BAX (1∶2000,美國Protein Tech Group公司)、Caspase-3 (1∶100,英國Abcam公司)、p-PI3K (1∶1000,phospho-Tyr458,美國Cell Signaling Technology公司)、p-AKT (1∶1000,phosphoSer473,美國Affinity Biosciences公司)和β-actin (1∶2000,美國Protein Tech Group公司)。最后,加入適量二抗稀釋液,室溫搖床2 h。二抗包括:辣根過氧化物酶標記山羊抗兔IgG (1∶1000,上海碧云天生物技術有限公司)和辣根過氧化物酶標記山羊抗小鼠IgG (1∶1000,上海碧云天生物技術有限公司)。二抗洗滌后,使用ECL化學發光液(上海翊圣生物科技有限公司)與膜孵育1 min,采用保鮮膜完全包裹印跡膜,在暗盒內與X膠片曝光10 min,顯影沖洗。
1.8 實時熒光定量PCR
采用TRIzol試劑盒(美國Thermo公司)提取細胞總RNA,采用核酸/蛋白濃度測定儀(NanDrop ND- 2000,美國Thermo Scientific公司)檢測總RNA質量,合格的總RNA使用PrimeScriptTMRT試劑盒(日本TaKaRa公司)進行cDNA逆轉錄。采用Oligo 6.0軟件設計實時熒光定量PCR (qRT-PCR)引物,由上海生工生物工程股份有限公司合成,引物信息見表1。構建25 μL qRT-PCR反應體系,包括12.5 μL SYBR Premix Ex(2×,日本TaKaRa公司)、1 μL上游引物、1 μL下游引物、2 μL cDNA模板和8.5 μL ddH2O。反應體系置于IQ-5熒光定量PCR儀(美國Bio-Rad公司)上采用如下程序進行qRT-PCR反應:95℃ 30 s;95℃ 5 s,60℃ 30 s,40個循環;55℃~95℃溶解30 s,81個循環。基因作為內參基因,每組設置3個重復。采用2–ΔΔCt公式計算各基因的相對表達水平。
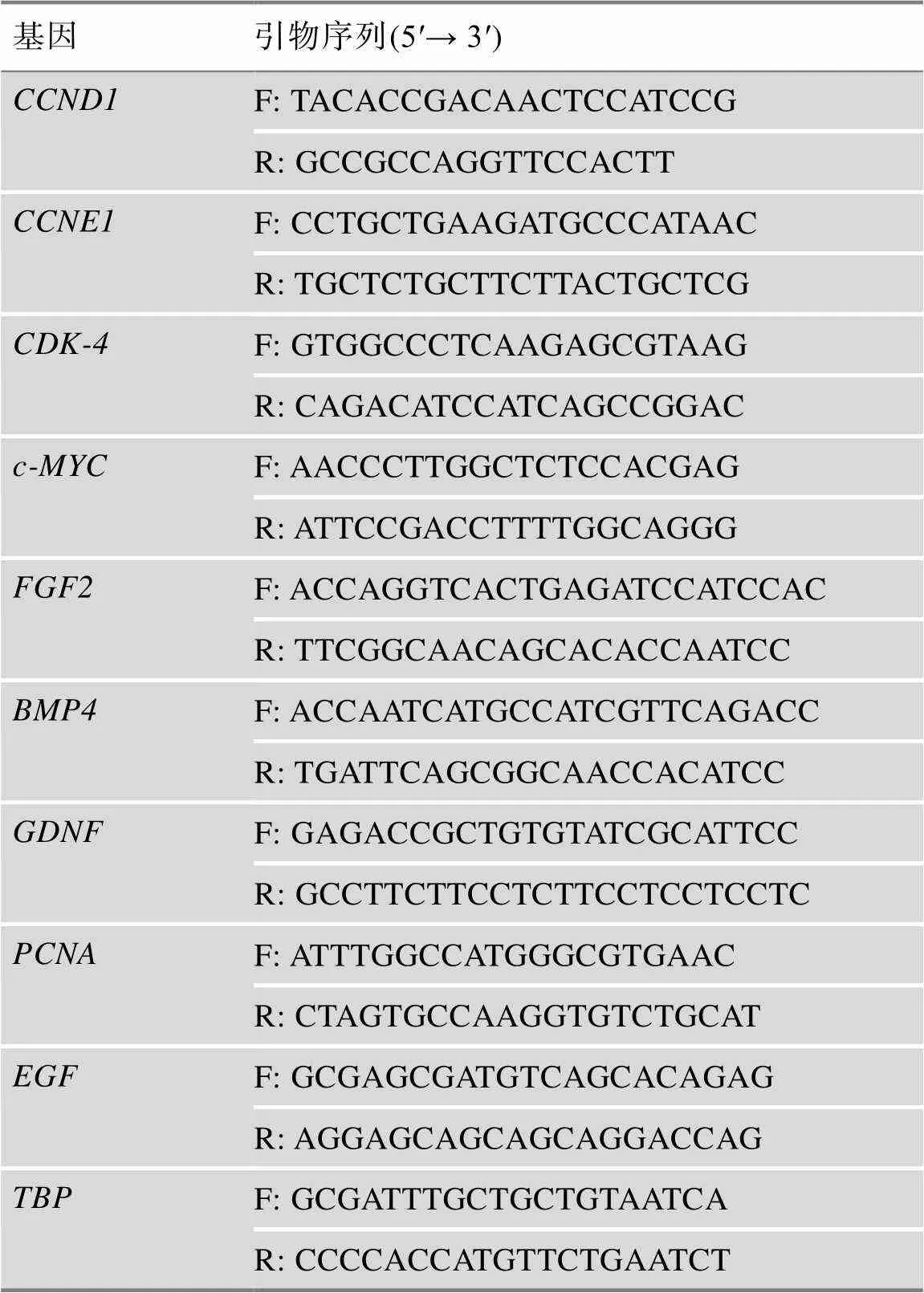
表1 本研究所用的引物序列
1.9 雙熒光素酶活性檢測
將合成的(brain-derived neurotrophic factor)基因3′-UTR-WT和3′-UTR-MUT序列(廣州市銳博生物科技有限公司)連接至psiCHECK2載體,篩選陽性克隆并測序確認后,與mimic和mimic NC在DMEM high glucose基礎培養基條件下兩兩組合共轉染至293 T細胞,48 h后收集細胞,每孔加35 μL PBS、35 μL luciferase reagent (美國Promega公司),震蕩10 min,移至96孔白色細胞培養板中,采用多功能酶標儀(Spectra max m5e,美國Molecular Devices公司)測定熒光值;每孔加30 μL Stop reagent (美國Promega公司),震蕩10 min后,測定熒光值。
1.10 統計分析
采用IBM SPSS 22.0軟件進行統計分析,使用One-Way ANOVA進行單因素方差分析,并采用Duncan氏法進行多重比較以評估各組之間的差異。數據均以平均值±標準差的形式表示,*<0.05,**<0.01。
2 結果與分析
2.1 miR-191促進豬未成熟支持細胞增殖
為明確對豬未成熟支持細胞增殖的作用,分別轉染mimic、mimic NC、inhibitor和inhibitor NC。流式細胞周期結果表明,過表達后,處于G1期的細胞比例極顯著降低(<0.01),S期和G2期的細胞比例顯著增加(< 0.05) (圖1A),抑制表達后,處于S期和G2期的細胞比例則顯著降低(<0.05) (圖1C)。采用qRT-PCR技術檢測細胞周期相關基因的表達,結果表明,過表達極顯著促進、、和基因的表達水平(<0.01) (圖1B),而抑制表達則顯著抑制以上基因的表達水平(<0.05) (圖1D)。以上結果表明,促進豬未成熟支持細胞周期進程。
采用qRT-PCR技術檢測細胞增殖相關基因的表達水平。結果表明,過表達顯著增加、、、和基因的表達水平(< 0.05) (圖1E),而抑制表達則顯著降低細胞增殖相關基因的表達(<0.05) (圖1F)。CCK-8試劑盒檢測細胞增殖情況,結果表明過表達極顯著提高細胞增殖能力(<0.01),而抑制表達則極顯著降低細胞增殖能力(<0.01) (圖1G)。此外,EdU染色結果表明,過表達顯著促進細胞增殖活性(<0.05),而抑制表達則極顯著降低細胞增殖活性(<0.01) (圖1:H和I)。
2.2 miR-191抑制豬未成熟支持細胞凋亡
為進一步明確對豬未成熟支持細胞凋亡的影響,本研究采用Annexin V-FIT/PI方法檢測細胞凋亡情況。結果表明,過表達顯著降低細胞凋亡率(<0.05) (圖2:A和B),而抑制表達則顯著增加細胞凋亡率(<0.05) (圖2:A和D)。細胞ATP水平檢測結果表明,過表達極顯著增加細胞ATP水平(<0.01) (圖2C),而抑制表達則顯著降低細胞ATP水平(<0.05) (圖2E)。采用Western blotting技術檢測細胞凋亡相關基因蛋白水平的表達,結果顯示,過表達增加Bcl2蛋白表達水平而抑制BAX和Caspase-3蛋白表達水平(圖2F),而抑制表達則與之相反(圖2G)。綜上所述,抑制豬未成熟支持細胞凋亡。
2.3 miR-191靶向BDNF基因3′-UTR
本研究利用miRwalk、TargetScan和miRanda在線軟件預測了不同物種中的靶基因,根據結合位點保守性分析,初步篩選出基因作為候選靶基因(圖3A)。為進一步確定與基因之間的靶向關系,本研究構建了-WT和-MUT雙熒光素酶報告基因載體,并與mimic和mimic NC兩兩組合共轉染至293T細胞。雙熒光素酶活性檢測結果表明,-WT +mimic共轉染組的雙熒光素酶活性顯著低于其他3組(<0.05) (圖3B)。采用qRT-PCR技術檢測對基因mRNA表達的影響,結果表明,過表達極顯著降低基因mRNA表達水平(<0.01),而抑制表達極顯著增加基因mRNA表達水平(<0.01)。綜上所述,靶向基因3′-UTR并抑制其表達水平。
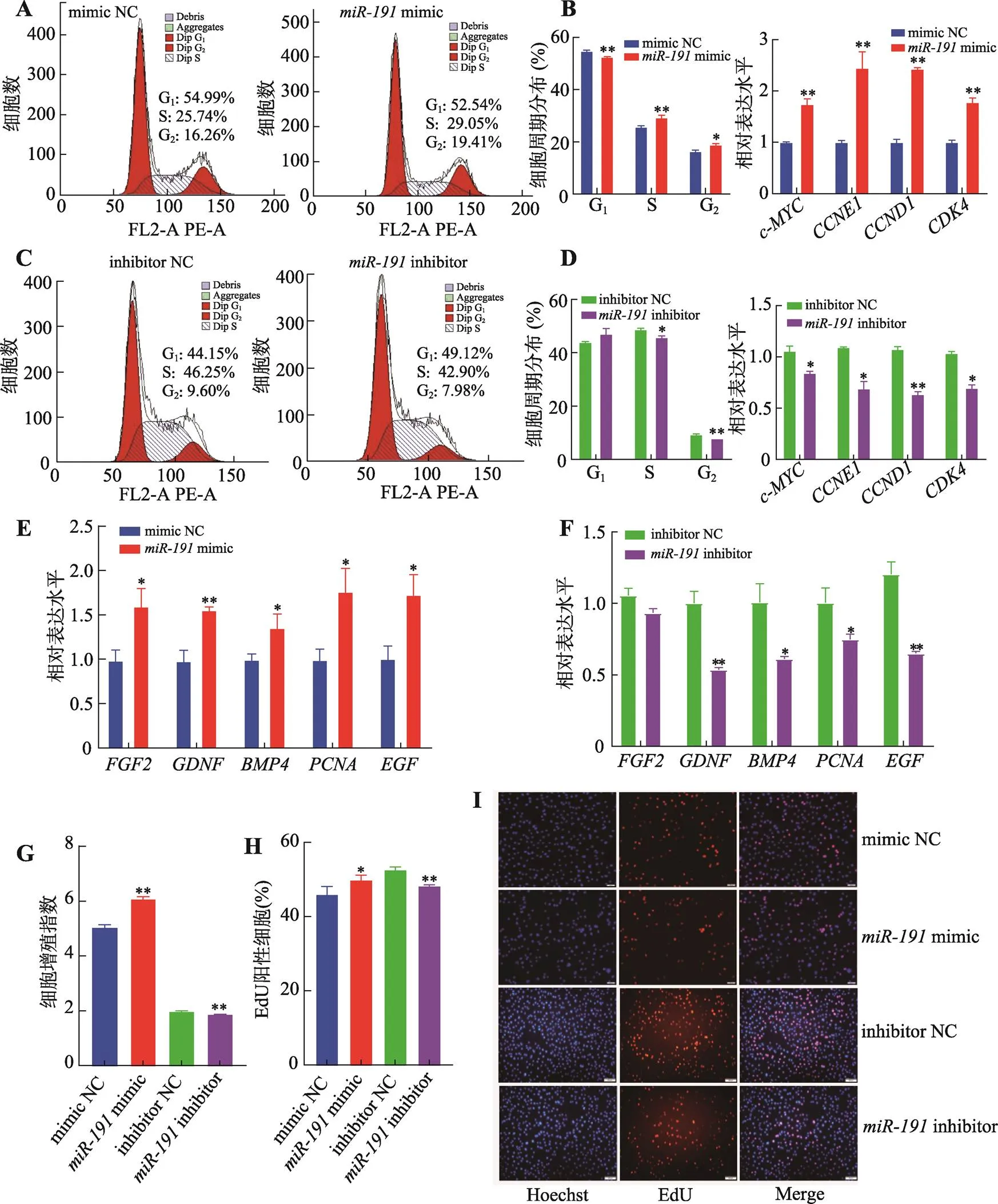
圖1 miR-191促進豬未成熟支持細胞增殖
A:轉染mimic和mimic NC,流式細胞儀檢測流式周期;B:轉染mimic和mimic NC,qRT-PCR檢測周期相關基因(、、和) mRNA相對表達水平;C:轉染inhibitor和inhibitor NC,流式細胞儀檢測流式周期;D:轉染inhibitor和inhibitor NC,qRT-PCR檢測周期相關基因(、、和) mRNA相對表達水平;E:轉染mimic和mimic NC,qRT-PCR檢測增殖相關基因(、、、和) mRNA相對表達水平;F:轉染inhibitor和inhibitor NC,qRT-PCR檢測增殖相關基因(、、、和) mRNA相對表達水平;G:轉染mimic、mimic NC、inhibitor和inhibitor NC,CCK-8試劑檢測細胞增殖指數;H:轉染mimic、mimic NC、inhibitor和inhibitor NC,EdU試劑盒檢測增殖細胞比例;I:細胞EdU實驗染色圖(藍色細胞為Hoechst染色,紅色細胞為EdU染色,Merge為Hoechst和EdU染色合并圖,標尺:50 μm)。*<0.05表示差異顯著,**<0.01表示差異極顯著。
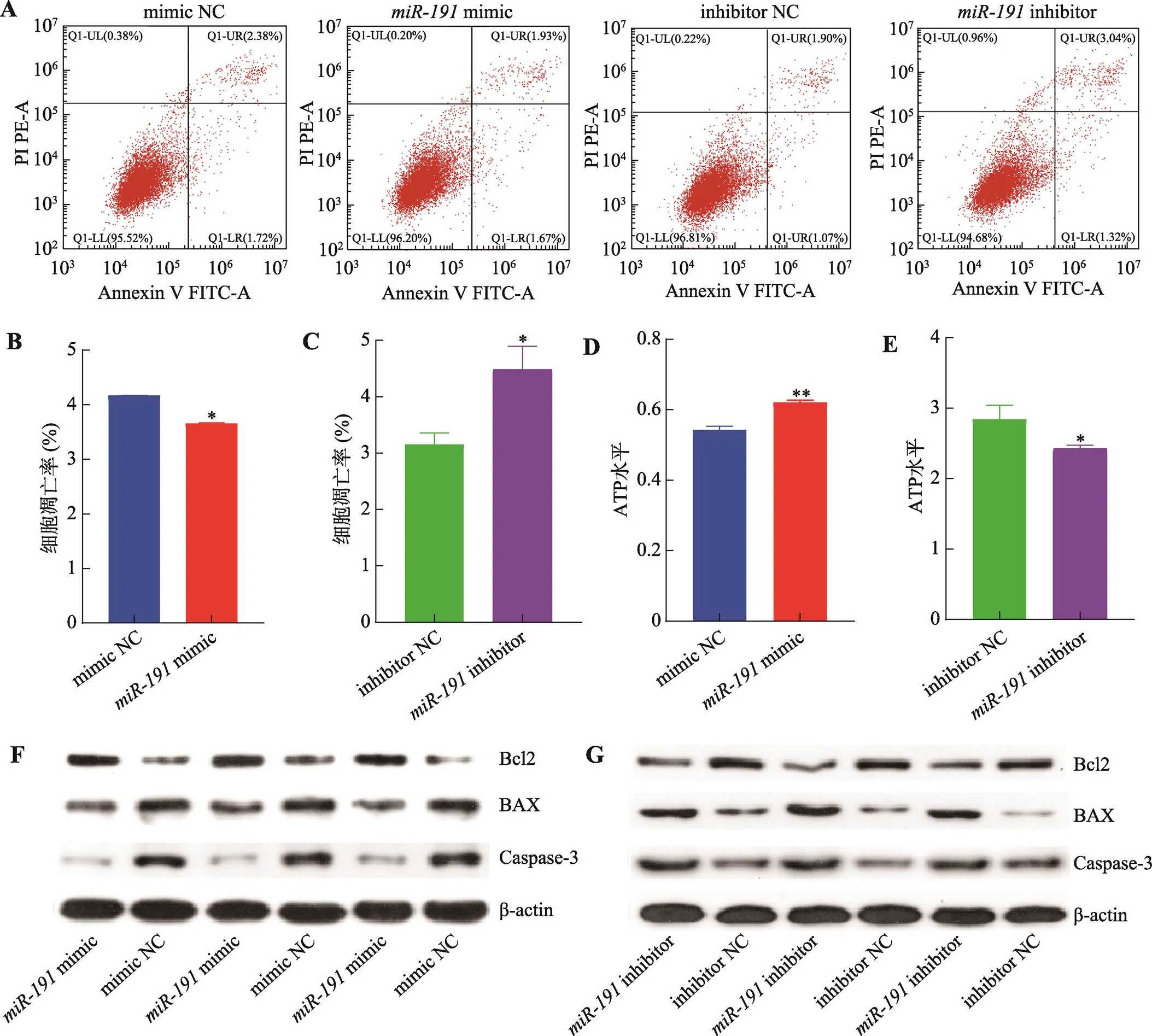
圖2 miR-191抑制豬未成熟支持細胞凋亡
A:轉染mimic、mimic NC、inhibitor和inhibitor NC,流式細胞儀檢測細胞凋亡情況;B:轉染mimic和mimic NC,細胞凋亡比例統計結果;C:轉染inhibitor和inhibitor NC,細胞凋亡比例統計結果;D:轉染mimic和mimic NC,細胞ATP水平;E:轉染inhibitor和inhibitor NC,細胞ATP水平;F:轉染mimic和mimic NC,Western blotting 檢測細胞凋亡標志基因Bcl2、BAX和Caspase-3蛋白表達水平;G:轉染inhibitor和inhibitor NC,Western blotting 檢測細胞凋亡標志基因Bcl2、BAX和Caspase-3蛋白表達水平。*<0.05表示差異顯著,**<0.01表示差異極顯著。
2.4 BDNF基因抑制豬未成熟支持細胞增殖而促進其凋亡
為解析基因對豬未成熟支持細胞增殖和凋亡的影響,針對基因序列設計了siRNA和siRNA NC,并轉染至細胞。結果表明,相較于對照組,轉染siRNA后,處于G1期和G2期的細胞比例顯著下降(<0.05),S期細胞比例極顯著上升(<0.01) (圖4A),、和基因的表達水平顯著增加(<0.05) (圖4B)。此外,抑制基因的表達后,細胞增殖相關基因、、、和基因表達水平顯著增加(<0.05) (圖4C);CCK-8和EdU檢測結果表明,抑制基因極顯著促進豬未成熟支持細胞增殖(<0.01) (圖4:D,E和F)。細胞凋亡檢測結果顯示,抑制基因的表達可極顯著抑制細胞的凋亡率(<0.01) (圖5A),促進Bcl2蛋白的表達而抑制BAX和Caspase-3蛋白的表達(圖5B),增加細胞ATP水平(圖5C)。以上結果表明,抑制表達基因促進豬未成熟支持細胞增殖而抑制其凋亡,與過表達的結果一致。
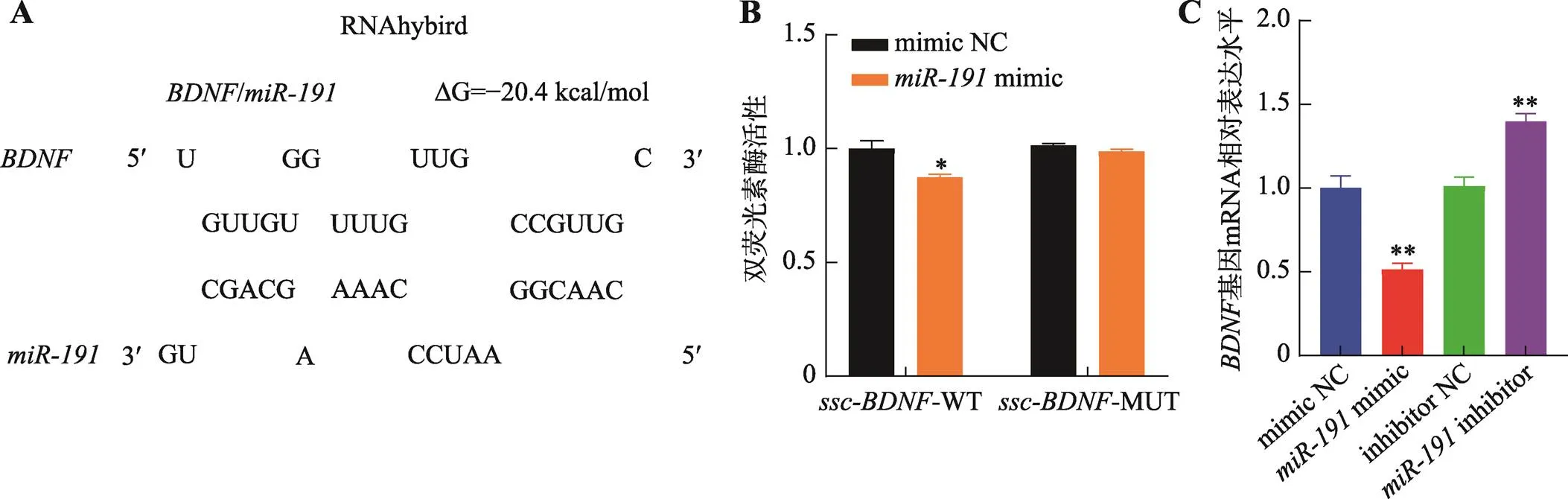
圖3 miR-191靶向BDNF基因3′-UTR
A:與基因3′-UTR的潛在結合位點;B:共轉染-WT + mimic NC、-WT +mimic、-MUT + mimic NC和-MUT +mimic后,檢測雙熒光素酶活性;C:轉染mimic NC、mimic、inhibitor NC和inhibitor,qRT-PCR檢測基因mRNA相對表達水平。*<0.05表示差異顯著,**<0.01表示差異極顯著。
2.5 BDNF基因拮抗miR-191的調控作用
為進一步明確是否通過靶向進而調控豬未成熟支持細胞增殖和凋亡,本研究將inhibitor +siRNA、inhibitor + siRNA NC和inhibitor NC + siRNA NC三組共轉染于豬未成熟支持細胞。EdU和CCK-8結果表明,相較于inhibitor NC + siRNA NC對照組,inhibitor + siRNA NC組的細胞增殖活性顯著降低(<0.05),而inhibitor +siRNA組的細胞增殖活性則極顯著升高(<0.01) (圖6:A和B)。同時,相較于inhibitor NC + siRNA NC對照組,inhibitor + siRNA NC組的細胞增殖相關基因、、、和的表達水平顯著降低(<0.05),而inhibitor +siRNA組的、和基因表達水平顯著增加(<0.05) (圖6C)。此外,相比于inhibitor NC + siRNA NC對照組,inhibitor + siRNA NC組的Bcl2蛋白表達水平降低且BAX和Caspase-3蛋白表達水平增加,而inhibitor +siRNA組的Bcl2蛋白表達水平升高且BAX和Caspase-3蛋白表達水平降低(圖6D)。以上結果表明,抑制表達基因可以拮抗對豬未成熟支持細胞增殖和凋亡的作用。
2.6 miR-191靶向BDNF基因3′-UTR激活PI3K/AKT信號通路
生物信息學分析結果表明,和基因可能作用于PI3K/AKT信號通路,因此,采用Western blotting技術檢測了二者對PI3K/AKT信號通路中p-PI3K和p-AKT蛋白的磷酸化水平。結果表明,過表達極顯著增加p-PI3K和p-AKT蛋白的磷酸化水平(<0.01) (圖7A),而抑制表達極顯著抑制p-PI3K和p-AKT蛋白的磷酸化水平(<0.01)(圖7B);抑制表達基因極顯著增加p-PI3K和p-AKT蛋白的磷酸化水平(<0.01) (圖7C),與過表達一致;同時,相比于inhibitor NC + siRNA NC對照組,inhibitor + siRNA NC組的p-PI3K和p-AKT蛋白的磷酸化水平極顯著降低(<0.01),而inhibitor +siRNA組的p-PI3K和p-AKT蛋白的磷酸化水平則顯著增加(<0.05) (圖7D)。以上結果表明,靶向基因通過激活PI3K/AKT信號通路促進豬未成熟支持細胞的增殖而抑制其凋亡。

圖4 抑制表達BDNF基因促進豬未成熟支持細胞增殖
A:流式細胞術檢測流式周期分布情況;B:qRT-PCR檢測周期相關基因、、和的mRNA相對表達水平;C: qRT-PCR檢測增殖相關基因、、、和的mRNA相對表達水平;D:CCK-8試劑檢測細胞增殖指數;E:細胞EdU實驗染色圖(藍色細胞為Hoechst染色,紅色細胞為EdU染色,Merge為Hoechst和EdU染色合并圖,標尺:50 μm);F:EdU細胞染色統計結果。*<0.05表示差異顯著,**<0.01表示差異極顯著。
3 討論
支持細胞數量是影響精子生成能力的主要因素,而提高公豬的精子產量,對實際生產中提高豬的生產效率至關重要。但是,目前關于支持細胞增殖的分子機制尚不清楚[20]。研究表明,miRNA在發育過程中調控許多基因的表達,且一些特異性的miRNA參與了未成熟支持細胞增殖的調控過程,例如[15]、[10]、[8]和[13]等。然而,分析表明,現有的關于miRNA調控豬未成熟支持細胞增殖的研究,還不足以解析其潛在的分子調控機制[21]。本課題組前期采用高內涵篩選技術鑒定出包括在內的27個miRNA具有促進豬未成熟支持細胞增殖的潛在作用,但其作用的特定分子信號途徑有待進一步研究。目前,關于的研究主要集中于醫學領域,例如,促進肝癌細胞[16]、食管鱗狀細胞[17]和成纖維細胞[18]等的增殖,抑制膽管癌細胞[22]、乳腺癌細胞(MCF7和ZR-75)[23]和子宮內膜樣癌細胞[24]等的凋亡。本研究綜合采用流式細胞術、CCK-8、EdU和Western blotting等方法在細胞水平明確了促進豬未成熟支持細胞增殖而抑制其凋亡,表明廣泛參與調控細胞增殖和凋亡,并有可能是調控豬未成熟支持細胞的一個特異性分子。
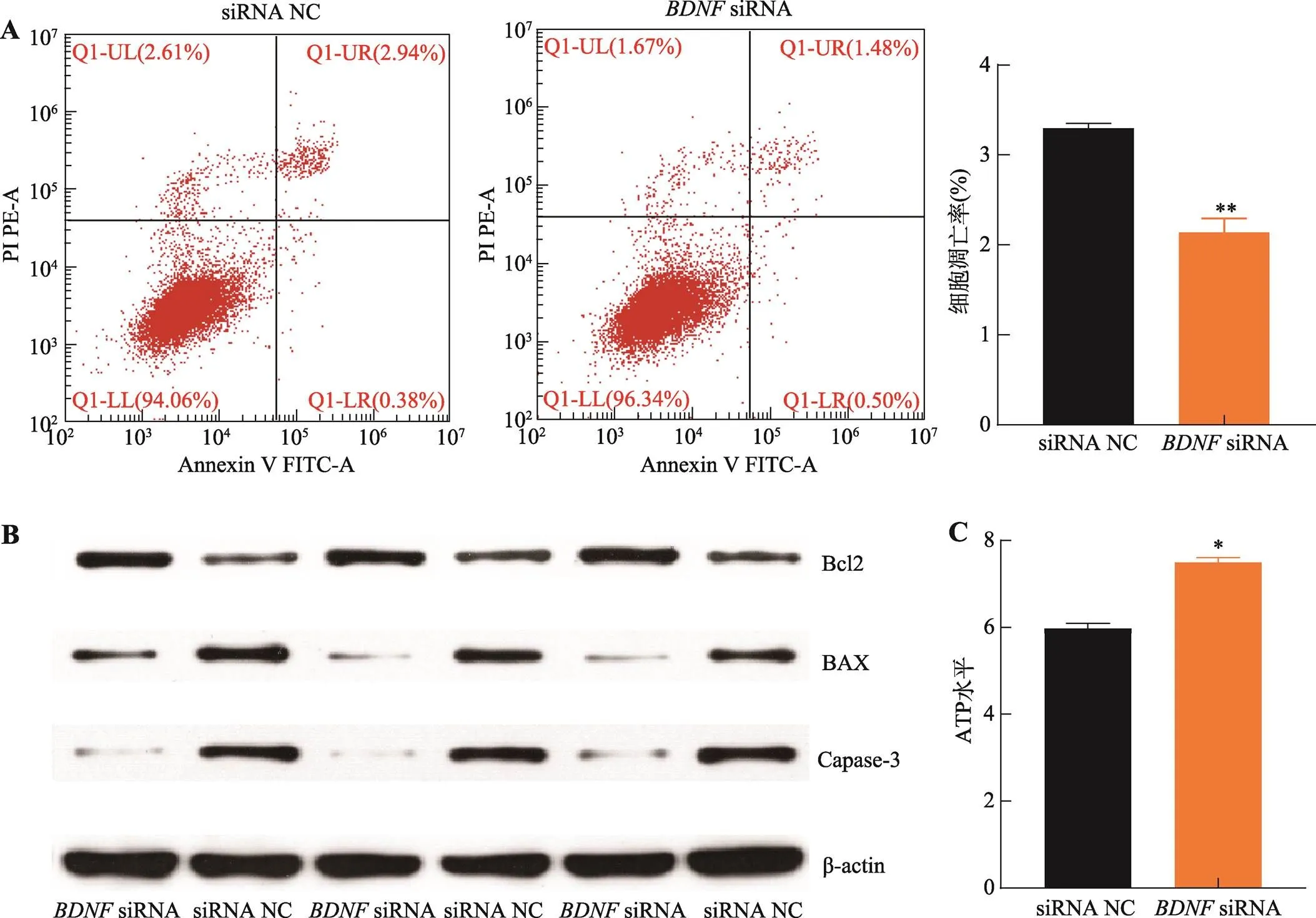
圖5 抑制表達BDNF基因促進豬未成熟支持細胞凋亡
A:流式細胞儀檢測不同凋亡期細胞分布及細胞數統計;B:Western blotting 檢測細胞凋亡標志基因Bcl2、BAX和Caspase-3蛋白表達水平;C:細胞內ATP水平檢測。*<0.05表示差異顯著,**<0.01表示差異極顯著。
miRNA通常靶向結合蛋白編碼基因3′-UTR以抑制其翻譯過程。本研究采用miRwalk、TargetScan和miRanda軟件預測的靶基因,并取結果的交集,再根據結合位點的保守性篩選出基因為其潛在靶基因。隨后,通過雙熒光素酶報告基因實驗進一步證實直接靶向基因3′-UTR的預測結合位點。但是,目前尚沒有豬基因商業化蛋白抗體,導致無法在現有條件下進一步驗證是否抑制了基因的蛋白翻譯過程。但是,在成肌細胞和海馬神經元細胞中已證實可以靶向基因并抑制其翻譯過程[25~27]。基于與基因結合位點在物種間的高度保守性,因此我們推測在豬未成熟支持細胞中同樣可以抑制基因的翻譯過程。同時,本研究明確了抑制表達基因可促進豬未成熟支持細胞增殖并抑制細胞凋亡的,其調控作用與過表達一致。在此基礎上,本文進一步設計了關于與基因的共轉染試驗,結果表明基因可以拮抗對豬未成熟支持細胞增殖和凋亡的調控作用。綜上所述,本文基本確定靶向基因3′-UTR并調控了其表達。
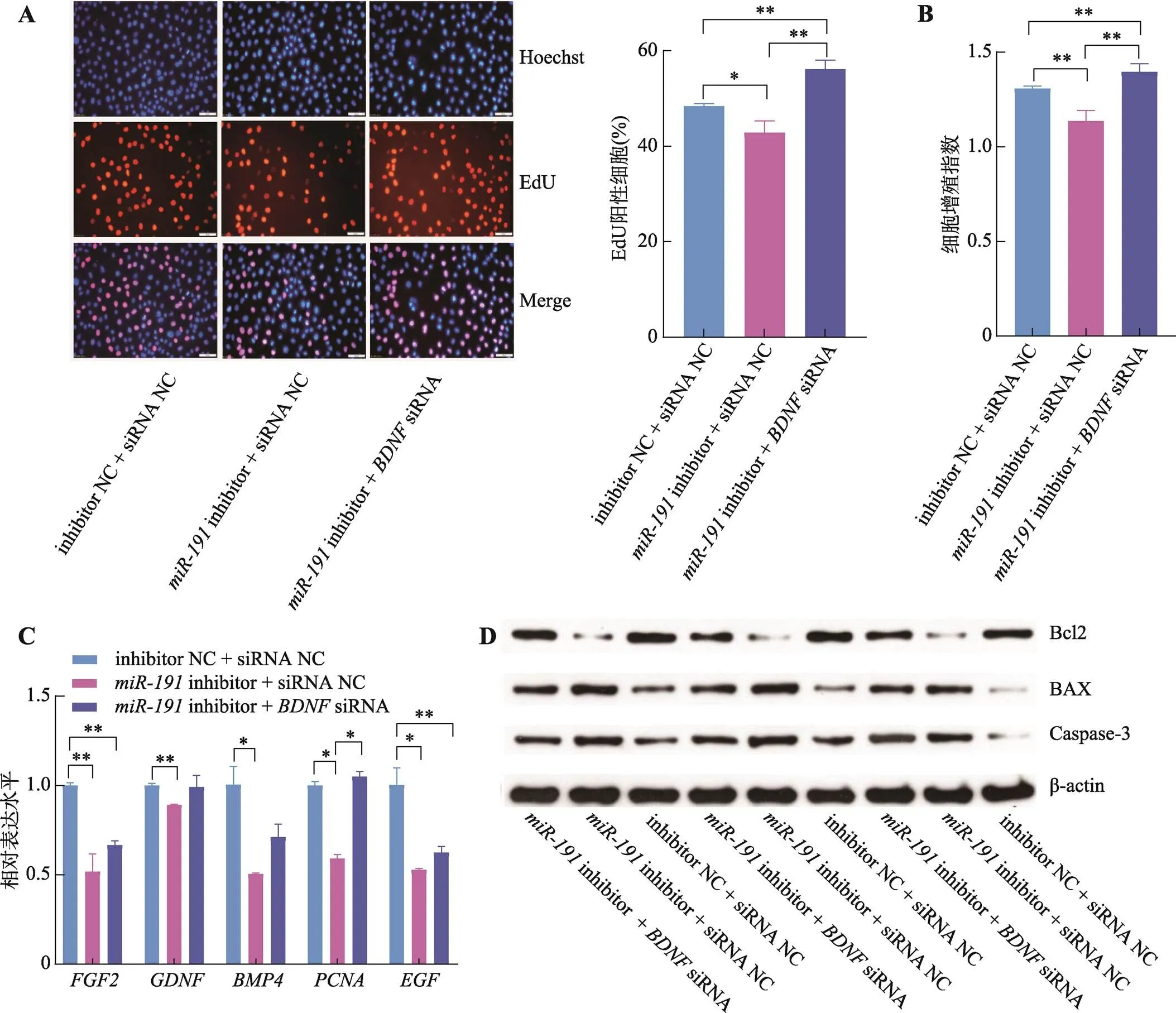
圖6 BDNF基因拮抗miR-191對豬未成熟支持細胞增殖和凋亡的調控作用
A:細胞EdU實驗染色圖(藍色細胞為Hoechst染色,紅色細胞為EdU染色,Merge為Hoechst和EdU染色合并圖,標尺:50 μm);B:CCK-8試劑檢測細胞增殖指數;C:qRT-PCR檢測增殖相關基因、、、和的mRNA相對表達水平;D:Western blotting 檢測細胞凋亡標志基因Bcl2、BAX和Caspase-3蛋白表達水平。*<0.05表示差異顯著,**<0.01表示差異極顯著。
細胞周期是一個受多個因子調控的復雜過程,主要參與生物的生長和發育,C作為細胞進程中一個重要的調節因子,活化后可以縮短G1期的時間,是從G1期轉化至S期必不可少的調節因子之一[28],還可以通過活化細胞周期蛋白、和而促進周期進程,若抑制的表達時,細胞周期的G1期向S期的轉化將受到抑制,進而抑制細胞周期進程[29]。本研究發現,過表達和抑制表達基因后,支持細胞由G1期進入S期的比例均顯著增加,且、、和的表達水平顯著增加。此外,本研究還發現,過表達和抑制表達基因可顯著增加、、、和的表達水平。研究表明,支持細胞本身表達以上細胞增殖相關基因,且和基因可通過激活MAPK信號通路促進細胞增殖[30],基因則可以通過調控細胞周期相關基因的表達促進細胞周期的進程以及促進DNA復制進而促進細胞增殖[31],基因則可以通過激活基因家族促進細胞增殖[32]。此外,本研究采用CCK-8和EdU實驗進一步證實和基因可以調控豬睪丸支持細胞增殖。
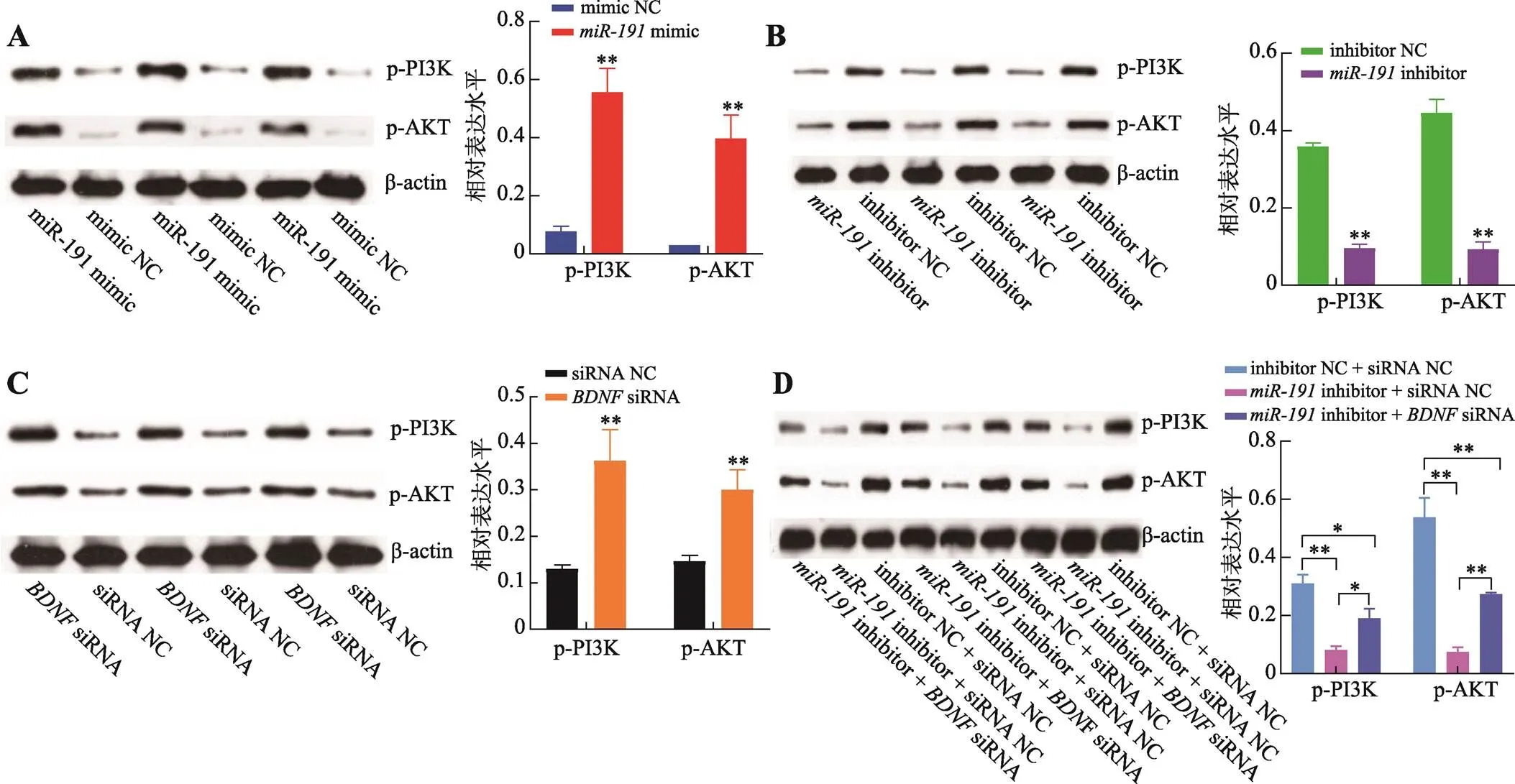
圖7 miR-191靶向BDNF基因激活PI3K/AKT信號
A:轉染mimic和mimic NC,Western blotting檢測p-PI3K和p-AKT蛋白相對表達水平及結果分析;B:轉染inhibitor和inhibitor NC,Western blotting 檢測p-PI3K和p-AKT蛋白相對表達水平及結果分析;C:轉染siRNA和siRNA NC,Western blotting檢測p-PI3K和p-AKT蛋白相對表達水平及結果分析;D:共轉染inhibitor NC + siRNA NC、inhibitor + siRNA NC和inhibitor +siRNA,Western blotting 檢測p-PI3K和p-AKT蛋白相對表達水平及結果分析。*<0.05表示差異顯著,**<0.01表示差異極顯著。
細胞凋亡是由內源性或外源性通路引起的細胞程序性死亡的過程,多個基因參與到細胞凋亡調控過程中,但基因在整個過程起決定性作用,細胞凋亡基因調控的方式分為促進細胞凋亡和抑制細胞凋亡兩種[33]。是促細胞凋亡基因,是抑細胞凋亡基因,二者都是通過激活下游基因以調節細胞凋亡。對各種因素引起的凋亡都有抑制作用,但能與線粒體上的結合從而促進細胞凋亡[34]。Caspase-3是Caspase家族中重要的一員,是一類與細胞密切相關的蛋白水解酶,通過自身活化或相互激活形成活性復合物引起瀑布式的級聯反應,導致細胞凋亡,在整個過程中發揮著最后的樞紐作用[35]。本研究結果表明,過表達和抑制表達基因后,豬睪丸支持細胞中Bcl2蛋白表達水平增加,而BAX和Capase-3蛋白表達水平降低,且細胞ATP水平顯著增加,細胞凋亡率顯著減少。
為進一步解析靶向基因調控豬未成熟支持細胞增殖和凋亡的信號機制,本文對所有的潛在靶基因進行了功能富集分析,預測出PI3K/AKT信號通路為潛在路徑。同時,研究表明,基因與PI3K/AKT信號通路之間具有調控關系。本研究結果表明,過表達和抑制表達基因均可顯著促進PI3K/AKT信號通路中關鍵蛋白PI3K和AKT的磷酸化水平,且基因同樣拮抗對PI3K和AKT蛋白的調控作用。在我們的前期研究中發現,740Y-P激活劑誘導的PI3K/AKT信號通路激活顯著促進豬未成熟支持細胞周期進程和細胞增殖,而抑制細胞凋亡[15],且LY294002抑制劑誘導的PI3K/AKT信號通路減活則顯著抑制豬未成熟支持細胞增殖而促進其凋亡[10]。此外,甲狀腺激素、促卵泡激素和鄰苯二甲酸二丁酯等均可以通過抑制PI3K/AKT信號通路活性抑制支持細胞增殖而促進細胞凋亡[36~38],17β-雌二醇則通過激活PI3K/AKT信號通路促進支持細胞增殖[39]。同時,[40]、[10]和[41]等均被報道通過調節PI3K/AKT信號通路活性以調控支持細胞增殖和凋亡。因此,PI3K/AKT信號通路的活性可能對于豬未成熟支持細胞的增殖和凋亡具有重要作用。綜上所述,靶向基因通過激活PI3K/AKT信號通路促進豬未成熟支持細胞增殖而抑制其凋亡。
[1] Franca LR, Hess RA, Dufour JM, Hofmann MC, Griswold MD. The Sertoli cell: one hundred fifty years of beauty and plasticity, 2016, 4(2): 189–212.
[2] Rebourcet D, Darbey A, Monteiro A, Soffientini U, Tsai YT, Handel I, Pitetti JL, Nef S, Smith LB, O'Shaughnessy PJ. Sertoli cell number defines and predicts germ and leydig cell population sizes in the adult mouse testis, 2017, 158(9): 2955–2969.
[3] Bertoldo MJ, Guibert E, Faure M, Guillou F, RaméC, Nadal-Desbarats L, Foretz M, Viollet B, Dupont J, Froment P. Specific deletion of AMP-activated protein kinase (α1AMPK) in mouse Sertoli cells modifies germ cell quality, 2016, 423: 96–112.
[4] Ran ML, Chen B, Wu MS, Liu XC, He CQ, Yang AQ, Li Z, Xiang YJ, Li ZH, Zhang SW. Integrated analysis of miRNA and mRNA expression profiles in development of porcine testes, 2015, 5(78): 63439–63449.
[5] Ding HS, Liu M, Zhou CF, You XB, Su T, Yang YB, Xu DQ. Integrated analysis of miRNA and mRNA expression profiles in testes of Duroc and Meishan boars, 2020, 21(1): 686.
[6] Luo ZY, Dai XL, Ran XQ, Cen YX, Niu X, Li S, Huang SH, Wang JF. Identification and profile of microRNAs in Xiang pig testes in four different ages detected by Solexa sequencing, 2018, 117: 61–71.
[7] Chen XX, Zheng Y, Li XL, Gao Q, Feng TY, Zhang PF, Liao MZ, Tian XE, Lu HZ, Zeng WX. Profiling of miRNAs in porcine Sertoli cells, 2020, 11: 85.
[8] Ma CP, Song HB, Yu L, Guan KF, Hu PD, Li Y, Xia XY, Li JL, Jiang SW, Li FG. miR-762 promotes porcine immature Sertoli cell growth via the ring finger protein 4 (RNF4) gene, 2016, 6: 32783.
[9] Zhang SX, Guo J, Liang MD, Qi JJ, Wang ZB, Jian XR, Zhang ZB, Sun BX, Li ZH. miR-196a promotes proliferation and inhibits apoptosis of immature porcine Sertoli cells, 2019, 38(1): 41–48.
[10] Gao H, Ran ML, Luo H, Weng B, Tang XW, Chen Y, Yang AQ, Chen B. miR-499 promotes immature porcine Sertoli cell growth by the PI3K/AKT pathway by targeting the PTEN gene, 2019, 152(2): 145–157.
[11] Hu Y, Deng J, Tian K, Yang WR, Luo NJ, Lian Y, Gan L, Tang XY, Luo HY, Zhang JJ, Wang XZ. MiR-8-3p regulates hyperthermia-induced lactate secretion by targeting PPP2R5B in boar Sertoli cells, 2019, 86(11): 1720–1730.
[12] Guo J, Yang C, Zhang SX, Liang MD, Qi JJ, Wang ZB, Peng YH, Sun BX. MiR-375 induces ROS and apoptosis in ST cells by targeting the HIGD1A gene, 2019, 685: 136–142.
[13] Zhang JJ, Wang Y, Yang WR, Kee JD, Wang XZ. MicroRNA-1285 regulates 17β-estradiol-inhibited imma-ture boar Sertoli cell proliferation via adenosine mono-phosphate-activated protein kinase activation, 2015, 156(11): 4059–4070.
[14] Ran M, Li Z, Cao R, Weng B, Peng F, He C, Chen B. miR-26a suppresses autophagy in swine Sertoli cells by targeting ULK2, 2018, 53(4): 864– 871.
[15] Luo H, Chen B, Weng B, Tang XW, Chen Y, Yang AQ, Chu D, Zeng XY, Ran ML. miR-130a promotes immature porcine Sertoli cell growth by activating SMAD5 through the TGF-β-PI3K/AKT signaling pathway, 2020, 34(11): 15164–15179.
[16] Tian F, Yu CT, Wu M, Wu XY, Wan LF, Zhu XJ. MicroRNA-191 promotes hepatocellular carcinoma cell proliferation by has_circ_0000204/miR-191/KLF6 axis, 2019, 52(5): e12635.
[17] Gao XT, Xie ZQ, Wang ZG, Cheng KL, Liang K, Song ZQ. Overexpression of miR-191 predicts poor prognosis and promotes proliferation and invasion in esophageal squamous cell carcinoma, 2017, 58(6): 1101–1110.
[18] Polioudakis D, Abell NS, Iyer VR. MiR-191 regulates primary human fibroblast proliferation and directly targets multiple oncogenes, 2015, 10(5): e0126535.
[19] Ma CP, Song HB, Guan KF, Zhou JW, Xia XY, Li FG. Characterization of swine testicular cell line as immature porcine Sertoli cell line, 2016, 52(4): 427–433.
[20] Ni FD, Hao SL, Yang WX. Multiple signaling pathways in Sertoli cells: recent findings in spermatogenesis, 2019, 10(8): 541.
[21] Berger T, Nitta-Oda BJ. Sertoli cell proliferation in juvenile boars and microRNA, 2020, 233: 103954.
[22] Kang PC, Leng KM, Liu YP, Liu Y, Xu Y, Qin W, Gao JJ, Wang ZD, Tai S, Zhong XY, Cui YF. miR-191 inhibition induces apoptosis through reactivating secreted frizzled- related protein-1 in cholangiocarcinoma, 2018, 49(5): 1933–1942.
[23] Sharma S, Nagpal N, Ghosh PC, Kulshreshtha R. P53-miR-191-SOX4 regulatory loop affects apoptosis in breast cancer, 2017, 23(8): 1237–1246.
[24] Tian XY, Xu LM, Wang P. MiR-191 inhibits TNF-α induced apoptosis of ovarian endometriosis and endo-metrioid carcinoma cells by targeting DAPK1, 2015, 8(5): 4933–4942.
[25] Varendi K, Kumar A, H?rma MA, Andressoo JO. miR-1, miR-10b, miR-155, and miR-191 are novel regulators of BDNF, 2014, 71(22): 4443–4456.
[26] Li HX, Du MQ, Xu WM, Wang ZF. MiR-191 down-regulation protects against isoflurane-induced neuroto-xicity through targeting BDNF, 2021, 31(5): 367–373.
[27] Mohammadipoor-Ghasemabad L, Sangtarash MH, Sheibani V, Sasan HA, Esmaeili-Mahani S. Hippocampal microRNA- 191a-5p regulates BDNF expression and shows correlation with cognitive impairment induced by paradoxical sleep deprivation, 2019, 414: 49–59.
[28] Trumpp A, Refaeli Y, Oskarsson T, Gasser S, Murphy M, Martin GR, Bishop JM. c-Myc regulates mammalian body size by controlling cell number but not cell size, 2001, 414(6865): 768–773.
[29] Ramana KV, Tammali R, Srivastava SK. Inhibition of aldose reductase prevents growth factor-induced G1-S phase transition through the AKT/phosphoinositide 3-kinase/ E2F-1 pathway in human colon cancer cells, 2010, 9(4): 813–824.
[30] Blum Y, Mikelson J, Dobrzynski M, Ryu H, Jacques MA, Jeon NL, Khammash M, Pertz O. Temporal perturbation of ERK dynamics reveals network architecture of FGF2/ MAPK signaling, 2019, 15(11): e8947.
[31] Strzalka W, Ziemienowicz A. Proliferating cell nuclear antigen (PCNA): a key factor in DNA replication and cell cycle regulation, 2011, 107(7): 1127–1140.
[32] Duval N, Vaslin C, Barata TC, Frarma Y, Contremoulins V, Baudin X, Nedelec S, Ribes VC. BMP4 patterns Smad activity and generates stereotyped cell fate organization in spinal organoids, 2019, 146(14): 175430.
[33] Volkmann N, Marassi FM, Newmeyer DD, Hanein D. The rheostat in the membrane: Bcl-2 family proteins and apoptosis, 2014, 21(2): 206–215.
[34] Correia C, Lee SH, Meng XW, Vincelette ND, Knorr KLB, Ding HS, Nowakowski GS, Dai HM, Kaufmann SH. Emerging understanding of Bcl-2 biology: implications for neoplastic progression and treatment, 2015, 1853(7): 1658–1671.
[35] Savitskaya MA, Onishchenko GE. Mechanisms of apoptosis, 2015, 80(11): 1393–1405.
[36] Riera MF, Regueira M, Galardo MN, Pellizzari EH, Meroni SB, Cigorraga SB. Signal transduction pathways in FSH regulation of rat Sertoli cell proliferation, 2012, 302(8): E914–E923.
[37] Sun Y, Yang WR, Luo HL, Wang XZ, Chen ZQ, Zhang JJ, Wang Y, Li XM. Thyroid hormone inhibits the proliferation of piglet Sertoli cell via PI3K signaling pathway, 2015, 83(1): 86–94.
[38] Wang C, Zheng P, Adeniran SO, Ma MJ, Huang FS, Adegoke EO, Zhang GX. Thyroid hormone (T3) is involved in inhibiting the proliferation of newborn calf Sertoli cells via the PI3K/Akt signaling pathway in vitro, 2019, 133: 1–9.
[39] Yang WR, Zhu FW, Zhang JJ, Wang Y, Zhang JH, Lu C, Wang XZ. PI3K/Akt activated by GPR30 and Src regulates 17β-estradiol-induced cultured immature boar Sertoli cells proliferation, 2017, 24(1): 57–66.
[40] Hu PD, Guan KF, Feng Y, Ma CP, Song HB, Li Y, Xia XY, Li JL, Li FG. miR-638 inhibits immature Sertoli cell growth by indirectly inactivating PI3K/AKT pathway via SPAG1 gene, 2017, 16(23): 2290–2300.
[41] Luo H, Peng FZ, Weng B, Tang XW, Chen Y, Yang AQ, Chen B, Ran ML. miR-222 suppresses immature porcine Sertoli cell growth by targeting the GRB10 gene through inactivating the PI3K/AKT signaling pathway, 2020, 11: 581593.
MiR-191 promotes the porcine immature Sertoli cell proliferation by targeting thegene through activating the PI3K/AKT signaling pathway
Xiangwei Tang, Dan Chu, Saina Yan, Yanfei Yin, Qiao Bian, Bo Weng, Bin Chen, Maoliang Ran
The number of Sertoli cells in the testis is a major regulator on the sperm production capacity. MicroRNAs (miRNAs) participate in regulating the proliferation and apoptosis of porcine immature Sertoli cells. However, the functions and mechanisms of action of most identified miRNAs in porcine Sertoli cells remain largely unknown. In the present study, based on our previous results from an EdU-based high-content screening assay, we further studied the mechanism of action ofon the proliferation and apoptosis of porcine immature Sertoli cells through flow cytometry, Western blotting, and dual-luciferase activity analyses. The results demonstrated that overexpression ofpromoted cell cycle progression from G1phase to the S and G2phases, enhanced cell proliferation, and inhibited apoptosis in the porcine immature Sertoli cells, whereasinhibition resulted in the opposite effects. The results from a luciferase reporter assay showed thatdirectly targeted the 3′-UTR of thegene.knockdown also promoted cell cycle progression to the S phase, cell proliferation and inhibited cell apoptosis, which were consistent with the effects of theoverexpression. A co-transfection experiment showed thatknockdown abolished the effects ofinhibition. Furthermore, bothoverexpression andinhibition elevated the phosphorylation of PI3K and AKT, the key components of the PI3K/AKT signaling pathway, whereasinhibition offset the effects of theknockdown. Overall, these data indicated thatpromotes cell proliferation and inhibits apoptosis in porcine immature Sertoli cells by targeting thegene through activating the PI3K/AKT signaling pathway. This study provides a novel scientific basis for further investigation on the biological functions ofon porcine spermatogenesis.
;gene; PI3K/AKT signaling pathway; proliferation; porcine Sertoli cells
2021-04-25;
2021-06-02
湖南省自然科學基金項目(編號:2020JJ4348),湖南省重點研發計劃項目(項目編號:2020NK2024)和湖南省生豬產業技術體系崗位專家項目資助[Supported by the Natural Science Foundation of Hunan Province (No. 2020JJ4348), the Key Research and Development Plan Projects of Hunan Province(No. 2020NK2024), and the Project from Porcine Industry and Technology System of Hunan Province]
唐湘薇,在讀碩士研究生,專業方向:豬遺傳育種。E-mail: 993808546@qq.com
冉茂良,博士,副教授,碩士生導師,研究方向:豬遺傳育種。E-mail: ranmaoliang0903@126.com
陳斌,博士,教授,博士生導師,研究方向:豬遺傳育種。E-mail: chenbin7586@126.com
10.16288/j.yczz.21-154
2021/6/11 17:50:36
URI: https://kns.cnki.net/kcms/detail/11.1913.R.20210611.1452.002.html
(責任編委: 李明洲)
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
學苑創造·A版(2015年11期)2016-01-14 09:03:27
