錳脅迫下雞眼草的富集特征及生理響應
2021-07-19 07:08:58李欣航肖澤華匡雪韶王悟敏羅亮宇劉文勝
草業學報 2021年7期
李欣航,肖澤華,匡雪韶,王悟敏,羅亮宇,劉文勝
(中南林業科技大學生命科學與技術學院,湖南長沙410004)
錳(Mn)是植物生長發育必需的微量營養元素,作為代謝酶的組分參與許多生物化學反應[1]。過量的錳會抑制植物生長,甚至可通過食物鏈危害人體健康[2]。然而,由于長期不合理的礦山開采、冶金以及錳肥等富錳物質的濫用,我國許多地區土壤錳污染日益嚴重。因此,錳污染區的生態修復與治理已成為刻不容緩的任務。
近年來,人們已采用多種方式開展礦區治理。植物修復因具有環境友好和經濟實惠等特點被廣泛關注,成為礦區污染治理的重要方式[3]。開展植物修復的關鍵是篩選一批耐錳性強、經濟美觀的植物或生態型,并揭示它們的錳耐受性機制。生長在重金屬污染土壤上的植物,由于長期經受高重金屬濃度的選擇壓力,逐漸適應該環境;其耐受重金屬脅迫的能力較生長在非污染土壤上的植物強,進而形成了不同的生態型[4]。因而,重金屬污染區植物具有與非污染區植物不同的重金屬耐受性。開展兩種來源植物重金屬耐受性特征的比較,是篩選植物修復材料并揭示其耐受機制的重要途徑。
植物生長、生理生化特征和重金屬富集能力均為衡量植物對重金屬耐受性的重要指標,可反映植物對重金屬脅迫的適應能力。例如,劉玉通等[5]對比研究不同生態型蜈蚣草(Pteris vittata)在鉛(Pd)脅迫下的表型特征,結果發現礦區生態型蜈蚣草的生物量受脅迫影響較小,比非礦區生態型蜈蚣草有更強的鉛耐受性;胡國濤等[6]通過研究兩個品種速生樹種竹柳(Salixsp.)對重金屬的生理反應,發現在重金屬脅迫作用下竹柳五號可維持較高的可溶性糖、可溶性蛋白含量和過氧化物酶、超氧化歧化酶活性,得出竹柳五號比竹柳三號對重金屬脅迫有更強的耐受性;王一峰等[7]比較研究了污染區與非污染區的烏蘇里風毛菊(Saussurea ussuriensis)的葉綠素含量、抗氧化酶系的酶活性、丙二醛含量、可溶性蛋白含量等生理生化特征,發現污染區烏蘇里風毛菊這些指標受脅迫變化較非污染區的小,表明其已經產生了適應性抗性進化。Zambrosia 等[8]通過對不同錳濃度下生長的6 個甘蔗(Saccharumspp.)品種的研究表明,耐性品種植物組織的錳含量與敏感品種相當或更高,植物組織錳含量是錳耐受性的重要指標。Ding 等[3]對兩個紅麻(Hibiscus cannabinus)品種在鉻(Cr)脅迫下的表型、生理生化與富集特征進行了比較研究,結果顯示隨著鉻濃度升高,植物體內重金屬含量增加,這些指標均可反映其耐受性。劉攀道等[9]通過研究對比兩種基因型圭亞那柱花草(Stylosanthes guianensis)在不同Mn2+濃度下的表型、生理生化及Mn積累特性,發現維持根系生長、降低植物組織錳含量和提高抗氧化酶活性是苗期柱花草TF226 耐錳的生理機制。這些研究說明植物生長、生理生化特征和富集特性可較好地揭示植物耐受性機制。
雞眼草(Kummerowia striata)為豆科一年生草本植物,具有生態適應性廣、繁殖能力強和能進行生物固氮等優點;還是一種優良的園林地被植物和重要中草藥[10],值得進一步開發利用。雞眼草在錳礦廢棄地的高錳含量地段生長繁茂,這表明該植物對錳具有較強耐受性,是一種兼性金屬型植物,也是一種理想的生物修復備選植物。近年來已有許多學者開展了雞眼草的種子萌發[10]、生物學特征[11]和水分脅迫響應[12]等方面的研究,這些研究表明了雞眼草具有較強的耐脅迫能力。本研究以污染區和非污染區雞眼草為材料,研究不同濃度Mn 脅迫下,兩種來源雞眼草的生長、重金屬累積特征及生理生化反應的變化規律,以期揭示雞眼草對錳脅迫的耐受機制,研究結果也可用于礦山恢復實踐。
1 材料與方法
1.1 種子采集及盆栽試驗
供試污染區雞眼草種子于2017 年11 月中旬采自湖南省湘潭市錳礦區(北緯28°03′,東經112°55′),該地段土壤錳含量達 17990.21 mg·kg?1(湖南省土壤錳含量平均值為 459 mg·kg?1[13])。非污染區雞眼草種子于 2017 年 11月下旬采自位于湖南省長沙市的中南林業科技大學校園內,其土壤錳含量達138.25 mg·kg?1。現場調查顯示這兩個地段雞眼草的分布面積均超過1 hm2,長勢較好,且均為自然生長。在兩個地段各選擇100 株長勢一致、果實已成熟的雞眼草植株,采集其地上部分并用信封裝好后帶回實驗室分離萌發所需種子。
分別選取兩種來源籽粒飽滿、大小均勻的雞眼草種子進行種子萌發。用0.5% NaClO 溶液將種子浸泡消毒30 min,然后用去離子水沖洗3~4 次后,最后將種子分別放置于干凈沙子中。根據預實驗結果及參照吳哲等[10]的結果,種子萌發在25 ℃、12 h 光照/12 h 黑暗、相對濕度80%的人工氣候箱中進行。待到雞眼草生長出5~6 片真葉時移栽至溫室大棚的花盆中繼續培養。
通過預備試驗以及參考潘高等[14]的研究,雞眼草的盆栽試驗在溫室大棚中開展,利用MnCl2·4H2O 設置0(對照)、1000、5000、10000、15000、20000 μmol·L?16 個錳濃度梯度。兩種來源的雞眼草每個濃度各設置 10 個花盆,每個花盆7 株幼苗,共設置120 個花盆、840 株幼苗。每個塑料花盆(直徑18 cm)中裝入2 kg 按1∶1 混合的珍珠巖和細沙。日常澆水,防病蟲害,每周澆一次Hoagland’s 營養液,保證植物正常生長。待幼苗在花盆中適應生長15 d 后,加入相應濃度的MnCl2溶液。在脅迫第30 天時,收獲試驗材料,測量雞眼草生長、生理生化和富集等相關指標。生長相關指標包括芽長、根長、鮮重和干重;生理生化指標包括可溶性糖、可溶性蛋白、脯氨酸和葉綠素含量及超氧化物歧化酶(superoxide dismutase,SOD)、過氧化物酶(peroxidase,POD)活性和丙二醛(malondialdehyde,MDA)含量;富集相關指標包括根、莖和葉各部分的Mn 含量。
1.2 測定項目及方法
1.2.1 生長指標的測定 植株芽長和根長的測定:污染區和非污染區雞眼草的每個處理隨機選取15 株植株,采用精度為1 mm 的直尺分別測量幼苗芽長、根長等指標。
植株鮮重和干重的測定:將雞眼草植株分為地上(芽)與地下(根)兩個部分,分別稱量鮮重,然后再于95 ℃下殺青10 min,65 ℃下烘干至恒重后稱重。

1.2.2 生理生化指標的測定 隨機選取污染區和非污染區雞眼草幼苗葉片,測定生理生化指標。采用蒽酮比色法測量可溶性糖(soluble sugar,SS)含量;采用考馬斯亮藍G-250 染色法測量可溶性蛋白(soluble protein,SP)含量;采用酸性茚三酮比色法測量脯氨酸含量;采用氮藍四唑法測量超氧化物歧化酶活性;采用愈創木酚法測量過氧化物酶活性;采用硫代巴比妥酸比色法測量丙二醛含量[16]。以上各指標均分別稱取污染區和非污染雞眼草新鮮葉片各0.1 g,重復3 次。
1.3 富集指標的測定
收獲的樣品先用去離子水沖洗干凈,再將根浸入20 mmol·L?1EDTA-2Na 溶液中交換15 min,以去除表面吸附的金屬離子,最后再用去離子水沖洗干凈,吸干植株表面的水分。將樣品分為根、莖、葉,測定各部分鮮重。將新鮮樣品放在95 ℃的烘箱內保持30 min,然后再在65 ℃下烘48 h,測定植物各部分干重,最后用不銹鋼粉碎機磨細,過2 mm 尼龍網篩,供分析測定。
采用濕法消解法[V(HNO3)∶V(HClO4)=3∶1][17]將植物材料進行消解至澄清,消解液用去離子水定容至 50 mL,用火焰原子吸收分光光度計(北京普析,TAS-990 型)測定Mn 含量。轉運系數反映植物吸收重金屬后從根部轉移到地上部分的能力,轉運系數(translocation factor,TF)為雞眼草地上部分與根部Mn 含量的比值。
1.4 數據處理
采用SPSS 21.0 軟件對兩種來源雞眼草的生長指標、生理指標、富集指標與脅迫濃度進行雙因素方差分析,并對每個指標不同處理之間的差異顯著性使用LSD 法進行多重比較。比較的數據結果均以“平均值±標準差”表示。用SigmaPlot 12.5 作圖。
2 結果與分析
2.1 錳脅迫對植物生長的影響
隨著錳濃度的升高,兩種來源雞眼草的根長、芽干重、根干重、根冠比均逐漸降低,芽長則呈先升后降的趨勢(表1)。當錳濃度達5000~20000 μmol·L?1時,與對照相比,污染區雞眼草的芽干重下降幅度為4.34%~27.71%,根干重下降幅度為19.00%~66.06%;非污染區雞眼草芽干重下降幅度為16.33%~49.77%,根干重下降幅度為27.90%~77.54%。兩種來源雞眼草在對照及1000 μmol·L?1時芽干重、根干重和根冠比無顯著差異;當錳濃度達5000 μmol·L?1及以上時,污染區雞眼草的根干重則均顯著高于非污染區。這說明雞眼草的生長受高錳脅迫抑制,且在高錳濃度時,污染區雞眼草受到抑制程度較低。
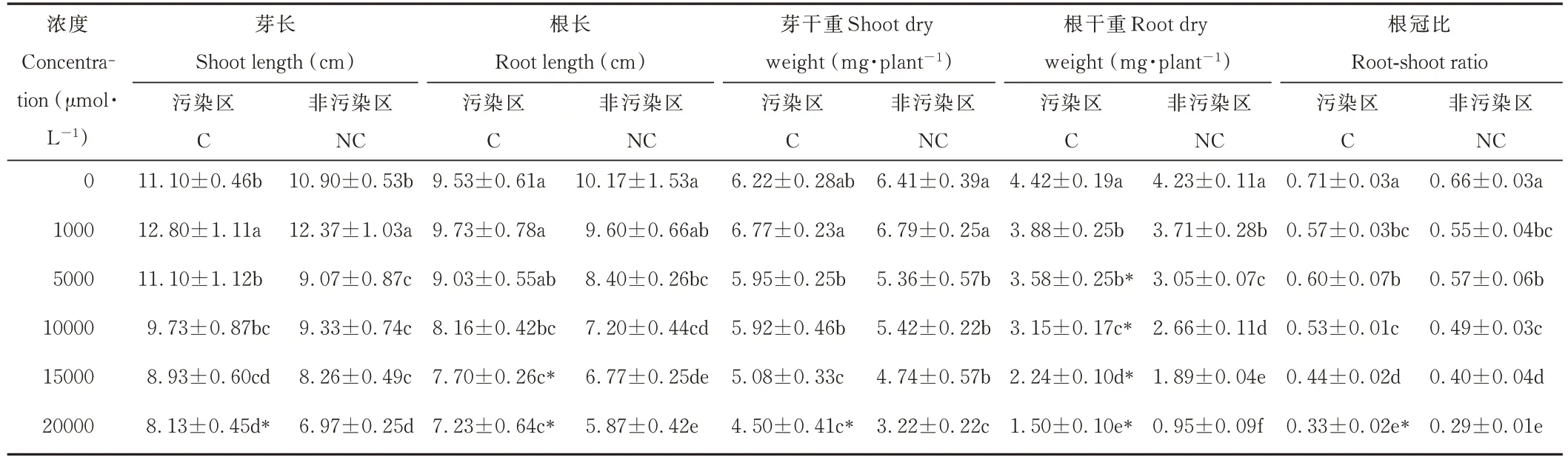
表1 錳脅迫對兩種來源雞眼草生長的影響Table 1 Effect of Mn stress on K.striata growth from two sources
2.2 錳脅迫對植物生理生化特征的影響
2.2.1 葉綠素和滲透調節物質含量 雙因素方差分析結果顯示,植物來源與錳濃度對葉綠素、可溶性糖、可溶性蛋白和脯氨酸含量均有極顯著影響,且兩者交互作用極顯著(圖1)。隨著錳濃度的升高,兩種來源雞眼草的葉綠素、可溶性糖和脯氨酸含量均呈先升后降的趨勢,可溶性蛋白的含量則呈逐漸下降的趨勢。錳濃度為1000 μmol·L?1時,兩種來源雞眼草的葉綠素含量達最大值;相同錳濃度下污染區雞眼草的葉綠素含量均顯著高于非污染區雞眼草。錳濃度為5000 μmol·L?1時,兩種來源雞眼草的可溶性糖和脯氨酸含量均達到最大值;當錳濃度高于5000 μmol·L?1時,污染區雞眼草可溶性糖和脯氨酸含量則均顯著高于非污染區。錳濃度低于1000 μmol·L?1時,污染區與非污染區雞眼草的可溶性蛋白含量無顯著差異;當錳濃度高于5000 μmol·L?1時,污染區雞眼草可溶性蛋白含量均顯著高于非污染區。這說明錳濃度高于5000 μmol·L?1時,污染區雞眼草葉綠素和滲透調節物質含量受到的影響小于非污染區。
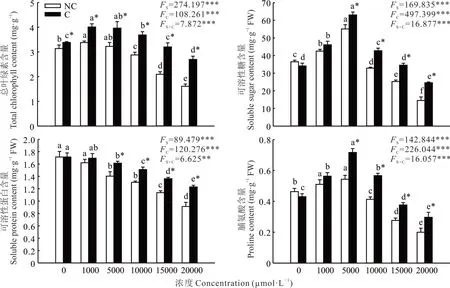
圖1 錳脅迫對兩種來源雞眼草生長過程中總葉綠素、可溶性糖、可溶性蛋白和脯氨酸含量的影響Fig.1 Effect of Mn stress on the total chlorophyll,soluble sugar,soluble protein and proline contents of leaves from two sources of K.striata
2.2.2 過氧化物酶、超氧化物歧化酶活性和MDA 含量 雙因素方差分析結果表明,錳濃度和植物來源對SOD、POD 酶活性和MDA 含量均有極顯著影響,且二者有極顯著的交互作用(圖2)。兩種來源雞眼草葉片的POD、SOD 活性隨錳濃度的升高呈先升后降的變化趨勢,且錳濃度高于5000 μmol·L?1時,污染區雞眼草的POD、SOD 活性均顯著高于非污染區;與對照相比,污染區雞眼草的SOD、POD 活性的下降程度低于非污染區。這說明錳濃度高于5000 μmol·L?1時,污染區雞眼草的SOD、POD 活性受到的抑制程度低于非污染區。
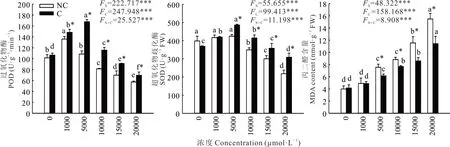
圖2 錳脅迫對兩種來源雞眼草過氧化物酶、超氧化物歧化酶活性和丙二醛含量的影響Fig. 2 Effects of Mn stress on activities of peroxidase,superoxide dismutase and malondialdehyde content in two sources of K.striata
隨著錳濃度的升高,兩種來源雞眼草葉片的MDA 含量均逐漸升高。當錳濃度小于1000 μmol·L?1時,兩種來源雞眼草的MDA 含量差異不顯著。錳濃度高于5000 μmol·L?1時,污染區雞眼草的MDA 含量則顯著低于非污染區(圖2)。
2.3 錳脅迫對植物組織錳含量的影響
由表2 可知,雞眼草各器官錳含量高低順序為:葉>根>莖。隨著錳濃度的升高,污染區和非污染區雞眼草的根、莖、葉中的錳含量均顯著增加;當錳濃度達20000 μmol·L?1時,污染區和非污染區雞眼草葉的錳含量分別達到了12553.03 和8558.05 mg·kg?1,為各自對照的16.53 和13.41 倍。兩種來源雞眼草相比,低濃度時污染區與非污染區雞眼草的根、莖、葉錳含量無顯著差異;高錳濃度時污染區雞眼草的根、莖、葉錳含量則均高于非污染區。污染區和非污染區雞眼草的轉運系數隨著錳濃度的升高而增加,污染區和非污染區雞眼草的轉運系數則無顯著差異。
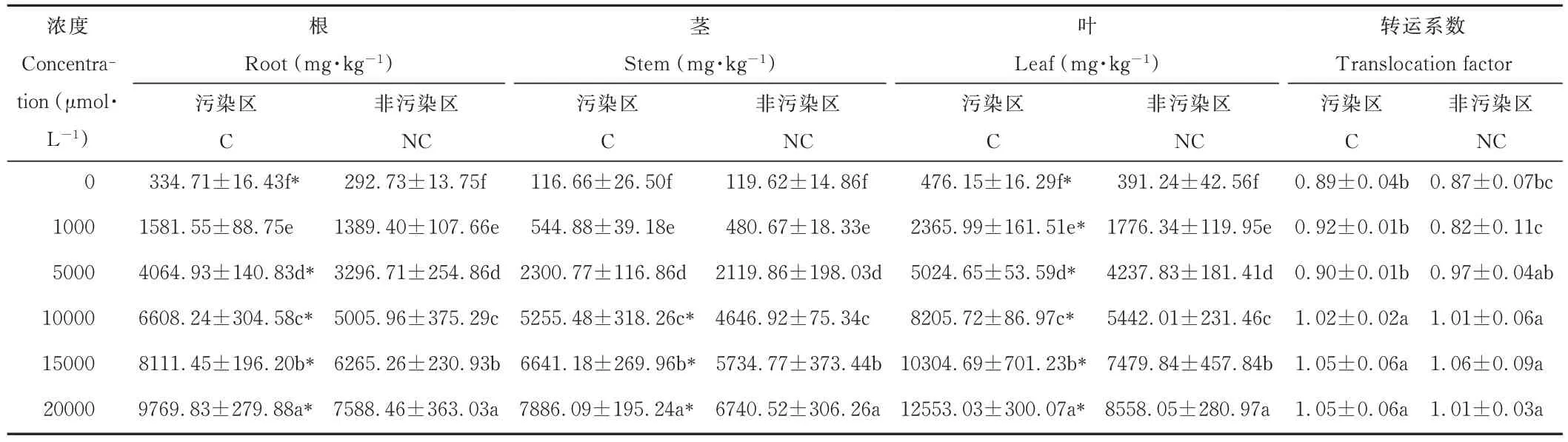
表2 錳脅迫下兩種來源雞眼草根、莖、葉的Mn 含量Table 2 Mn content of roots,stems and leaves from two sources of K.striata under Mn stress
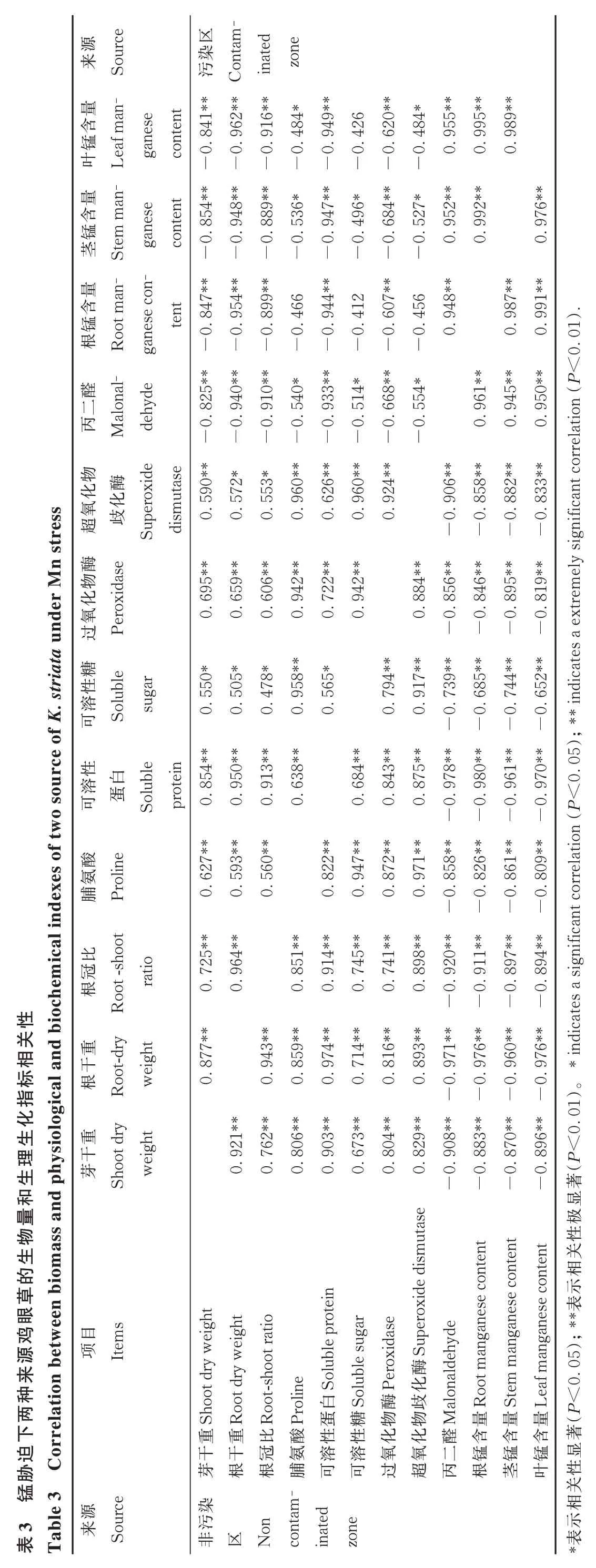
2.4 各指標的相關性
由表3 可知,兩種來源雞眼草的丙二醛含量、根錳含量、莖錳含量、葉錳含量與根干重、芽干重、根冠比、脯氨酸含量、可溶性蛋白含量、可溶性糖含量、SOD 活性、POD 活性之間均呈顯著負相關關系。兩種來源雞眼草的丙二醛含量、根錳含量、莖錳含量、葉錳含量兩兩之間均呈顯著正相關關系。兩種來源雞眼草的根干重、芽干重、根冠比、脯氨酸含量、可溶性蛋白含量、可溶性糖含量、SOD 活性、POD 活性兩兩之間均呈顯著正相關關系。
3 討論
3.1 生物量的變化
植物在逆境下的生長情況是植物對該環境適應性的最直接反映[18]。本研究結果顯示,隨著錳濃度的升高,兩種來源雞眼草的芽干重、根干重均逐漸降低,污染區雞眼草的芽干重、根干重的下降幅度低于非污染區雞眼草。Ding 等[3]、劉玉通等[5]、Zambrosia 等[8]的研究也發現隨著重金屬濃度的升高,植物生物量降低,耐性較強的基因型(或生態型)生物量受重金屬脅迫影響較低,本研究結果與此基本一致。這說明高濃度錳脅迫抑制雞眼草生長,污染區較非污染區雞眼草具有更強的錳耐受性。其原因可能是污染區雞眼草在長期處于高錳脅迫環境條件下,逐漸適應立地環境,已產生較強的錳耐受性。
3.2 生理生化特征的變化
3.2.1 葉綠素和滲透調節物質含量 葉綠素是植物光合作用的基礎物質,其含量反映了植物光合能力的高低。本研究結果顯示,隨著錳濃度的升高,兩種來源雞眼草葉綠素含量均呈先上升后下降的趨勢,且同濃度下污染區雞眼草的葉綠素含量高于非污染區,這與劉玉通等[5]、劉攀道等[9]、王一峰等[7]對重金屬脅迫下不同生態型對比的結果一致。這說明兩種來源雞眼草在高濃度錳脅迫時,其葉綠素均受到抑制;污染區雞眼草具有更強的耐受性。其原因是當錳濃度過高時,高濃度的錳會破壞葉綠體的結構,導致葉綠素合成受阻,從而降低了葉綠素的含量及光合作用能力;而污染區雞眼草可能已產生特殊機制來提高錳的耐受性。
生物體在代謝過程中,可通過有機滲透調節物質的積累和分解來調節細胞滲透平衡,以緩解重金屬脅迫導致的傷害[19]。本研究結果表明,隨著錳濃度的升高,兩種來源雞眼草幼苗的可溶性糖和脯氨酸含量呈先升高后下降的趨勢,可溶性蛋白含量呈逐漸下降的趨勢;當錳濃度高于5000 μmol·L?1時,污染區雞眼草的可溶性糖、可溶性蛋白和脯氨酸含量的下降程度均低于非污染區。這與劉攀道等[9]、王一峰等[7]、潘高等[14]的研究結果一致。其原因是錳作為植物的必需元素,低濃度的錳可以促進植物體內所需物質的合成,這可能使植物體內的糖和脯氨酸的含量增加;過量的錳則會使植物受到錳毒害,導致植物體內可溶性糖和脯氨酸的合成受阻,植物體內的可溶性蛋白、可溶性糖和脯氨酸含量降低。在錳脅迫條件下,污染區雞眼草的耐受性高于非污染區,可能是因為長期在污染區生長的雞眼草已經產生了對錳的適應性抗性進化,對錳產生了一定的抗性。
3.2.2 SOD、POD 活性和 MDA 含量 植物長期生長在逆境環境中,為了適應環境而進化出抗氧化保護酶系統。其中,SOD 是植物清除活性氧的重要物質,它可以將O?快速歧化為H2O2和O2;而 POD 則可將 H2O2進一步分解成 H2O 和 O2,從而達到植物的自我保護作用[20]。本研究中,隨著錳濃度的升高,兩種來源雞眼草葉片的SOD、POD 活性呈先升后降的變化趨勢,且污染區雞眼草的POD、SOD 活性變化幅度低于非污染區,這與劉玉通等[5]、王一峰等[7]的研究結果一致。說明在錳脅迫環境下,抗氧化酶的去氧化是植物重要的防御反應,使植物對逆境脅迫具有一定的調節能力;較強的去氧化能力,加強了活性氧的清除能力,使植物具有較強的重金屬耐受能力;而在高濃度重金屬脅迫下,抗氧化保護酶活性受到抑制,過高濃度的重金屬使植物抗氧化體系遭到破壞,從而削弱了其保護能力,造成對植物體的損傷[21]。另一方面,本研究也說明污染區雞眼草具有較強的去氧化能力,其對逆境的自我調節能力高于非污染區。其原因可能是污染區生長的雞眼草已經產生了對錳的適應性抗性進化,具有較強的抗氧化保護酶系統,是污染區雞眼草具有較強錳耐受性的重要生理機制。
?
植物在正常生長情況下,體內的活性氧物質會處于平衡狀態。當植物受到外界脅迫時,這種平衡被打破,導致植物體產生過量的活性氧物質。當植物體吸收一定量的重金屬后,會使植物組織中的重金屬含量增加,引起植物膜脂過氧化,MDA 含量升高,從而打破植物體內活性氧物質的平衡。本研究中,兩種來源雞眼草的MDA 含量均隨錳濃度的升高逐漸上升,同濃度下污染區雞眼草的MDA 含量顯著低于非污染區。說明污染區雞眼草在錳脅迫下表現出更強的抵抗能力,受到的傷害更小。這與趙建橋等[22]對鉛脅迫下2 種生態型五節芒(Miscanthus floridulus)的研究及王一峰等[7]對鉛脅迫下污染區與非污染區烏蘇里風毛菊研究中MDA 含量變化的結果相似。其原因可能是因為長期在污染區生長的雞眼草已經產生了對錳的適應性抗性進化,形成了對錳的抗性居群,因此在低錳濃度下產生的MDA 少于非污染區雞眼草。
3.3 錳脅迫下雞眼草的錳累積特征
本研究結果顯示,污染區和非污染區雞眼草根、莖、葉中的錳含量均隨著錳濃度的升高而增加。Ding 等[3]對鉻脅迫下紅麻體內鉻含量的研究及陳小米等[23]對鎘和鋅復合污染條件下竹柳重金屬含量的研究結果均顯示,植物體內重金屬含量隨著環境重金屬濃度的增加而增加。本研究結果與此一致,說明雞眼草具有較強的錳富集能力;植物體內重金屬含量隨環境重金屬濃度的增加而增加是一個普遍規律。本研究還顯示,相同濃度錳脅迫下,污染區雞眼草根、莖、葉的錳含量均高于非污染區。這與Gan 等[24]對銅(Cu)脅迫下長萼雞眼草(Kummerowia stipulacea)的研究、Pan 等[25]對錳脅迫下污染區與非污染區蒼耳(Xanthium strumarium)的研究結果一致。這說明污染區雞眼草對錳的吸收與富集能力高于非污染區。其原因可能是雞眼草長期生長在礦區高錳濃度的環境條件下,其種群產生了適應性遺傳分化,已進化形成了相應的能在體內累積較高濃度的錳而不受傷害的耐受性機制,例如螯合機制等。需要有更多的研究來驗證并揭示相關機制。
兩種來源雞眼草在錳濃度20000 μmol·L?1的脅迫條件下仍能生長,且污染區和非污染區雞眼草的葉片中的錳含量分別達到了12553.03 和8558.05 mg·kg?1,這說明雞眼草對錳具有較強的富集能力。雞眼草具有耐不良生長環境、繁殖能力強、生長迅速等特點,是一種非常好的園林地被植物。同種錳濃度下,污染區雞眼草植株內錳的含量顯著高于非污染區,所以利用污染區雞眼草是礦區土壤Mn 污染生態修復的一種較好的選擇。
4 結論
隨著錳濃度的升高,兩種來源雞眼草的芽干重、根干重均逐漸降低,污染區雞眼草降低幅度低于非污染區雞眼草。隨著錳濃度的升高,兩種來源雞眼草的葉綠素、可溶性糖和脯氨酸含量、SOD、POD 活性均呈先升后降的趨勢,可溶性蛋白含量逐漸下降,MDA 含量逐漸升高。隨著錳濃度的升高,污染區和非污染區雞眼草根、莖、葉中的錳含量均增加,污染區雞眼草較非污染區錳富集能力強。污染區雞眼草錳耐受能力及富集能力均高于非污染區,提高抗氧化酶活性是其重要的生理機制。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
