呼倫貝爾沙地樟子松人工林土壤細菌群落結構與功能預測
2021-07-12 01:39:54丁鈺珮杜宇佳高廣磊曹紅雨朱賓賓楊思遠張儆醒劉惠林
生態學報 2021年10期
關鍵詞:功能
丁鈺珮,杜宇佳,高廣磊,3,*,張 英,曹紅雨,朱賓賓,楊思遠,張儆醒,邱 業,劉惠林
1 北京林業大學水土保持學院,水土保持國家林業和草原局重點實驗室,北京 100083
2 北京師范大學地理科學學部,北京 100875
3 寧夏鹽池毛烏素沙地生態系統國家定位觀測研究站,鹽池 751500
4 呼倫貝爾市林業科學研究所,呼倫貝爾 021000
土壤細菌參與凋落物分解和腐殖質形成等生態過程,聯接著地上和地下生態系統,與植物和土壤聯系密切,是陸地生態系統物質能量循環的重要環節,深刻地影響著陸地生態系統的穩定、調節和修復[1- 2]。一方面,植被可以通過凋落物和根系分泌物,改善土壤水分、養分和酸堿度條件,直接或間接地影響土壤細菌群落結構[3]。另一方面,土壤細菌能夠通過代謝活動參與土壤生態過程,為植物提供所需的營養元素,促進植物的定植、生長、發育和演替[4-5]。此外,土壤細菌對環境變化還具有高度敏感性,其結構和功能變化能夠有效指示土壤環境或植被的變化[6]。因此,土壤細菌是土壤微生物研究的重要和熱點領域,得到了國內外學者的廣泛關注[2]。
在研究早期,受制于相對落后的實驗條件和研究方法,土壤細菌研究進展較為緩慢。近年來,受益于現代生物技術的飛速發展,國內外學者圍繞土壤細菌開展了大量卓有成效的研究工作,在土壤細菌群落結構、多樣性及其驅動因子等方面取得了豐碩的研究成果,極大地深化了我們對于土壤細菌的認知[4,7]。但是,由于土壤細菌豐富的物種、遺傳和功能多樣性及其與環境要素強烈的相互作用,復雜時空尺度條件和氣候變化背景下土壤細菌群落結構依然是相關研究領域的重要內容。同時,土壤細菌功能也得到越來越多的關注,相關研究也逐漸轉向土壤細菌功能預測與分析[8]。其中,土壤細菌功能與環境因子的互饋作用成為土壤細菌研究的重要方向[9- 10]。
PICRUSt(Phylogenetic Investigation of Communities by Reconstruction of Unobserved States)是一種基于系統發育與功能聯系假設,判斷和預測標記基因功能組成的微生物功能預測方法[11]。該方法采用16S rRNA測序基因與Greengenes數據庫進行對比,依據基因相似性預測細菌代謝功能譜。與Tax4Fun和FAPROTAX等分析方法相比,PICRUSt數據庫完善,方法便捷,功能先進,成本低廉,結果準確[12],已經成功應用于林地、草地、沙地等陸地生態系統土壤微生物功能分析,為深入理解土壤細菌功能提供了重要途徑[12-13]。
沙地樟子松(Pinussylvestrisvar.mongolica)具有耐寒、耐旱、耐瘠薄等優良特性,是我國“三北”地區重要的生態環境建設樹種[14]。呼倫貝爾沙地是我國四大沙地之一,既是沙地樟子松的原產地,也是沙地樟子松人工林的重要分布區,但營建沙地樟子松人工林對于土壤細菌群落結構和功能有何影響尚不明確。鑒于此,以呼倫貝爾沙地樟子松人工林為研究對象,采用16S rRNA高通量測序和PICRUSt功能預測相結合的研究方法,鑒定分析土壤細菌群落結構和功能特征,研究揭示土壤理化性質對于土壤細菌群落的影響,以期豐富完善土壤細菌群落結構、功能與環境因子互饋作用相關研究,并為沙地樟子松人工林的經營管理提供理論依據和科技支撐。
1 研究區概況
研究區位于內蒙古自治區呼倫貝爾市海拉爾國家森林公園(49°06′—49°28′ N,119°28′—120°34′ E),海拔約600—800 m,地處呼倫貝爾沙地,屬于中溫帶半干旱大陸性草原氣候。年均降水量約為346.5 mm,多集中于6—8月,年均氣溫-1.2℃,年均無霜期121 d,年均日照時數2701.5 h。境內河流主要有額爾古納河、海拉爾河和輝河等,干支流總長約960 km。土壤類型為風沙土,土層較厚。研究區林下基本無灌木,多為草本植物,主要包括羊草(Leymuschinensis(Trin.) Tzvel.)、針茅(StipacapillataLinn.)、地榆(SanguisorbaofficinalisL.)、委陵菜(PotentillachinensisSer.)和狗尾草(Setariaviridis(L.) Beauv.)等。
2 材料與方法
2.1 土壤樣品采集
2018年7月,在海拉爾國家森林公園用樹木生長錐測量各林分樹木年齡后,選取無經營管理措施、林下植被相似的25 a、34 a和43 a沙地樟子松人工林為實驗組,并選取鄰近地段沙質草地作為對照組(表1)。每個人工林樣地內分別選取3株標準木,在樹冠投影處采集土壤樣品,草地則在樣地內隨機選取3個取樣點采集土壤樣品,樣品采集深度為0—10和10—20 cm。在每個取樣點的同一土層隨機采集3個樣品并充分混和為1個樣品,4個樣地共獲得24份土壤樣品。期間,為防止樣品污染,采用75%酒精對取土鉆和鐵鏟滅菌消毒,以保證土壤微生物樣品采集在無菌條件下進行。同時,分別用環刀和鋁盒采集土壤樣品,用于測定土壤孔隙度和土壤含水量。所有土壤樣品置于4℃便攜式保溫箱保存,運回實驗室后轉入-80℃冰箱中保存。
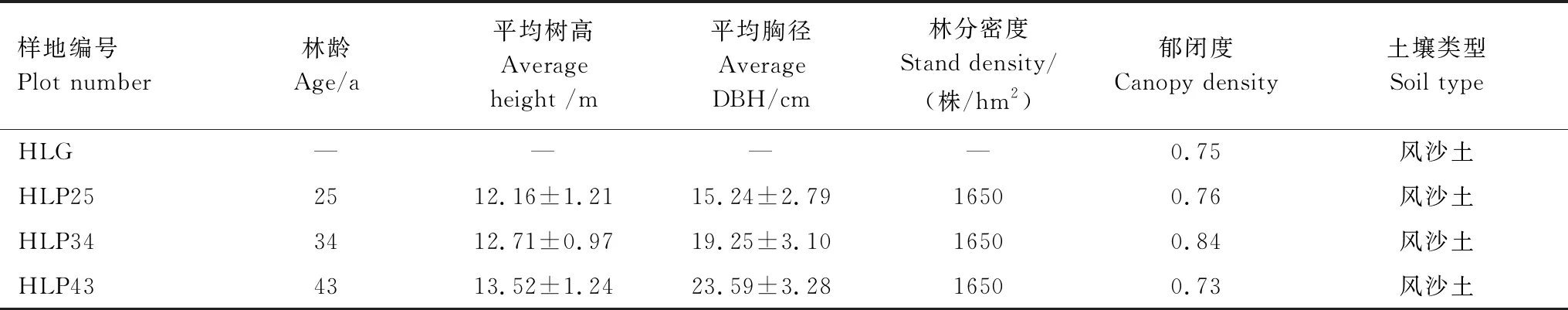
表1 樣地基本概況
2.2 土壤理化性質分析
將各樣地的新鮮土壤樣品自然風干后過2 mm篩進行土壤理化性質的分析。采用烘干法測定土壤含水量(soil water content, SWC),環刀法測定土壤孔隙度(soil porosity, SP),重鉻酸鉀稀釋熱法測定土壤有機質(soil organic matter, SOM),靛酚藍比色法測定全氮(total nitrogen, TN),鉬銻抗比色法測定全磷(total phosphorus, TP),火焰分光光度法測定速效鉀(available potassium, AK)[15]。
2.3 土壤細菌分離鑒定
使用PowerSoil DNA Isolation Kit試劑盒(MoBio Laboratories, Carlsbad, CA)提取土壤細菌樣品DNA,完成抽提后,利用1%瓊脂糖凝膠電泳檢測抽提的基因組DNA。對16S rRNA基因的V3—V4區域進行PCR擴增,引物序列為338F(5′-CCTACGGGAGGCAGCAG-3′)、806R(5′-ATTACCGCGGCTGCTGG-3′)。擴增條件設置:95℃預變性5 min,95℃變性45 s,55℃退火50 s,72℃延伸45 s,28個循環。每個樣品3次重復,將同一樣品的DNA混合,作為每份土壤樣品的總DNA。將 PCR產物混合后用2%瓊脂糖凝膠電泳檢測,使用AxyPrepDNA 凝膠回收試劑盒(AXYGEN 公司)切膠回收PCR產物,用Tris-HCl緩沖液洗脫后,使用2%瓊脂糖凝膠電泳檢測。
16S rRNA基因的PCR產物構建上機文庫,使用Qubit進行檢測定量,Nanodrop 粗檢文庫濃度,Agilent 2100 檢測文庫片段,qPCR 精確定量文庫濃度。在Illumina Miseq平臺對最后的文庫進行上機測序。測序結果使用uclust、uparse、cdhit等方法對97%的相似水平下的OTU進行聚類統計分析,得到門、綱、目、科、屬各水平群落的生物信息[16]。
2.4 數據處理與分析
采用SPSS 23.0計算平均值和標準差,進行單因素方差分析(One-way ANOVA),顯著性水平0.05。采用Mothur軟件計算Chao 1豐富度指數、Shannon多樣性指數和Pielou均勻度指數。將由QIIME軟件得到的biom文件上傳到Galaxy網站進行PICRUSt功能基因預測分析,得到土壤細菌功能基因組成。選擇土壤細菌相對豐度與土壤性質為數據矩陣,采用Canoco for Windows 4.5基于線性模型進行冗余分析(Redundancy Analysis, RDA),并進行蒙特卡洛檢驗(Monte-Carlo permutation test),檢驗重復數為499。
3 結果與分析
3.1 土壤細菌群落結構與多樣性
呼倫貝爾沙地樟子松人工林和沙質草地土壤中共獲得土壤細菌35門92綱109目210科267屬,其中相對豐度≥1%土壤細菌門類均為12個(圖1)。變形菌門(Proteobacteria)、放線菌門(Actinobacteria)和酸桿菌門(Acidobacteria)是研究區的主要土壤細菌優勢門,平均相對豐度分別為24.29%±3.39%、23.72%±4.10%和23.40%±2.55%。隨著林齡的增加,沙地樟子松人工林土壤優勢細菌門相對豐度呈現波動變化趨勢,25 a人工林0—10 cm層次變形菌門相對豐度顯著高于43 a人工林(P<0.05);各林地間放線菌門和酸桿菌門相對豐度不存在顯著差異(P>0.05)。沙質草地變形菌門和酸桿菌門相對豐度與人工林存在顯著差異(P<0.05),變形菌門相對豐度比人工林低6.05%,酸桿菌門相對豐度比人工林高4.54%。各樣地不同土層間土壤細菌組成較為相似,相對豐度無明顯變化。

圖1 呼倫貝爾沙地樟子松人工林土壤細菌相對豐度熱圖(門水平)
沙地樟子松人工林土壤細菌群落的Chao 1、Shannon和Pielou指數不存在顯著差異(P>0.05)(表2)。沙地樟子松人工林土壤細菌群落的Chao 1和Shannon指數在43 a人工林中最高,在25 a人工林中最低;Pielou指數在各林地中基本一致,土壤細菌分布較為均勻。沙質草地土壤細菌群落的Chao 1和Shannon指數與人工林不存在顯著差異(P>0.05),Pielou指數僅在0—10 cm土層與25 a和34 a人工林存在顯著差異(P<0.05)。此外,研究區土壤細菌群落的Chao 1、Shannon和Pielou指數均隨土層深度增加而降低,說明表層土壤細菌群落多樣性水平相對較高,且分布更為均勻。
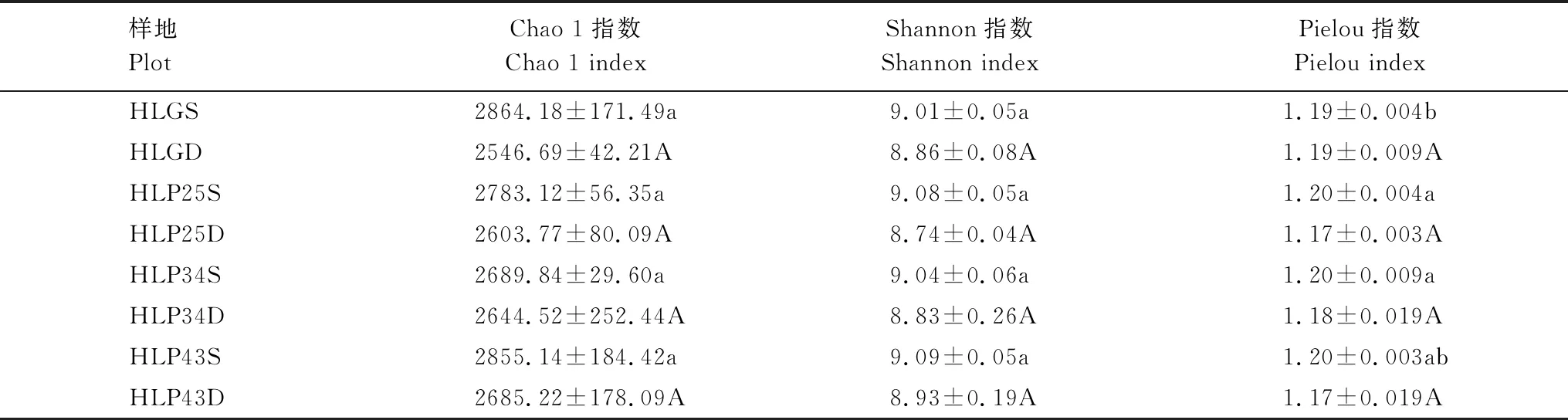
表2 呼倫貝爾沙地樟子松人工林細菌群落的Alpha多樣性指數
3.2 土壤細菌群落結構與土壤理化因子的相關性
呼倫貝爾沙地樟子松人工林土壤理化性質波動性較強(表3)。隨著林齡的增加,表層土壤含水量和有機質含量顯著降低(P<0.05),土壤孔隙度先降低后升高,全氮、全磷和速效鉀含量均表現為先增加再減少。沙質草地土壤的全氮和速效鉀含量顯著高于人工林(P<0.05),土壤孔隙度和全磷含量與人工林不存在顯著差異(P>0.05)。
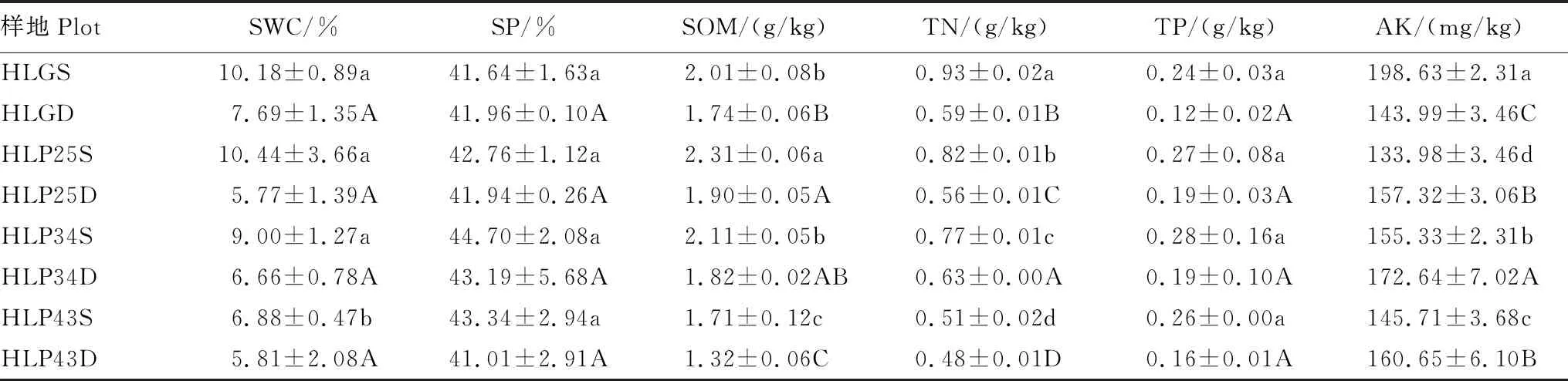
表3 呼倫貝爾沙地樟子松人工林土壤理化性質
冗余分析結果顯示(圖2),第一排序軸特征值為42.4%,第二排序軸特征值為10.8%,前兩軸累積解釋量占特征值的總和50%以上,RDA前兩軸能很好地反映土壤細菌群落與土壤因子之間關系的大部分信息[17- 18]。樣本點分布相對離散,說明不同樣地土壤細菌群落結構存在差異且受土壤理化性質影響較大。經蒙特卡洛檢驗,速效鉀、全磷和全氮是研究區土壤細菌群落的主要影響因子(P<0.05)。不同林齡沙地樟子松人工林土壤細菌群落結構與土壤理化因子的相關性有差異,25 a和34 a人工林土壤細菌的主要影響因子為全氮(P<0.05),43 a人工林土壤細菌的主要影響因子為土壤含水量(P<0.05)。沙質草地土壤細菌群落的主要影響因子為全磷(P<0.05)。此外,各樣地0—10 cm土層細菌群落與土壤含水量呈正相關,10—20 cm土層情況相反。
3.3 土壤細菌功能預測
呼倫貝爾沙地樟子松人工林土壤細菌共得到5個生物代謝通路(圖3)。其中,環境信息處理、代謝和遺傳信息處理是人工林土壤細菌功能的重要組成,其相對豐度分別為32.62%±0.98%、26.43%±0.31%和8.13%±0.28%。沙質草地一級代謝功能種類與人工林相同。
沙地樟子松人工林中,43 a人工林0—10 cm土層環境信息處理功能基因相對豐度顯著低于其他林地,代謝、遺傳信息處理和有機系統功能基因相對豐度顯著高于其他林地(P<0.05)。與沙質草地相比,25 a和34 a人工林0—10 cm土層環境信息處理和細胞過程功能基因相對豐度顯著高于沙質草地,代謝、遺傳信息處理和有機系統功能基因相對豐度顯著低于沙質草地(P<0.05)。此外,研究區土壤細菌代謝、遺傳信息處理、有機系統和細胞過程功能基因相對豐度均表現為0—10 cm>10—20 cm,沙質草地和43 a人工林土壤細菌環境信息處理功能基因相對豐度表現為0—10 cm<10—20 cm。
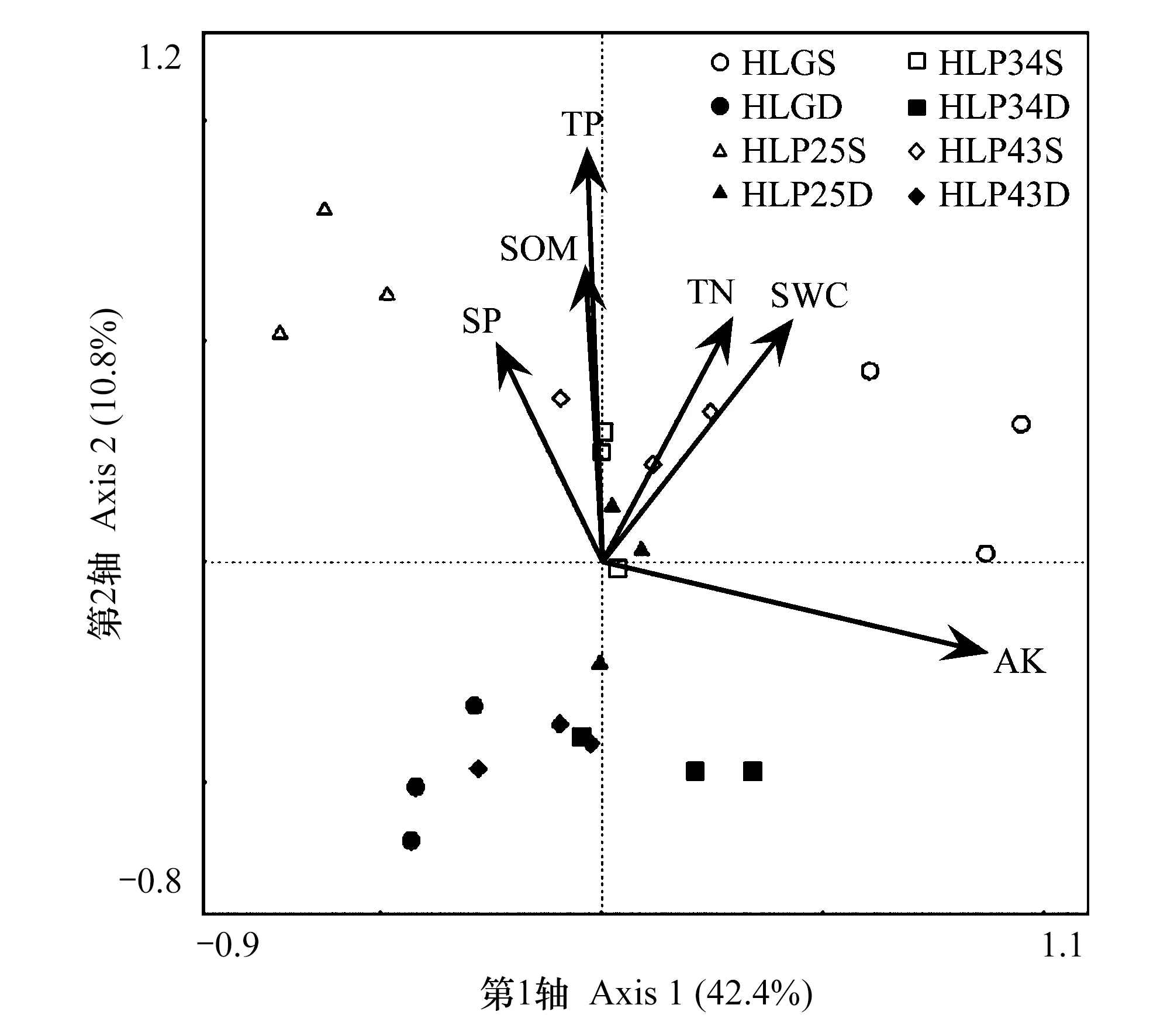
圖2 呼倫貝爾沙地土壤細菌群落與環境因子的冗余分析
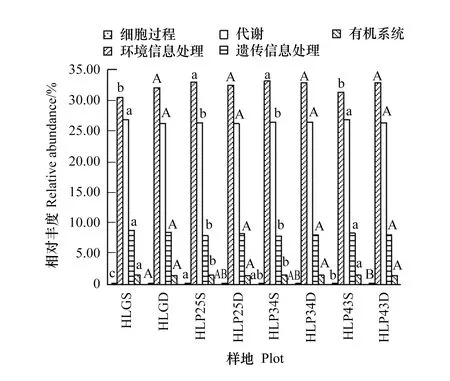
圖3 呼倫貝爾沙地樟子松人工林土壤細菌功能預測(一級功能層)
沙地樟子松人工林和沙質草地土壤細菌功能基因的5個一級功能層共有31個子功能。其中,共有10個二級功能層相對豐度>1%(圖4),且膜運輸功能占主導地位,其相對豐度為32.60%±0.98%,其他功能相對豐度均低于10%。沙地樟子松人工林中,43 a人工林0—10 cm土壤細菌復制和修復、酶家族、核苷酸代謝、能量代謝和碳水化合物代謝的功能基因相對豐度顯著高于其他林地,膜運輸功能基因相對豐度顯著低于其他林地(P<0.05)。25 a與34 a人工林所有二級功能相對豐度均不存在顯著差異(P>0.05)。與沙質草地相比,25 a和34 a人工林0—10 cm土層膜運輸功能基因相對豐度顯著高于沙質草地,其他二級功能基因相對豐度均顯著低于沙質草地(P<0.05)。此外,人工林和沙質草地各功能基因在不同土層深度相對豐度基本一致,無明顯變化規律。
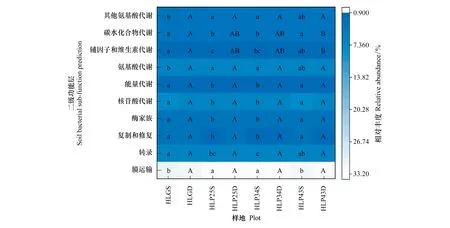
圖4 呼倫貝爾沙地樟子松人工林土壤細菌功能預測熱圖(二級功能層)
4 討論
4.1 呼倫貝爾沙地樟子松人工林土壤細菌群落結構特征
呼倫貝爾沙地樟子松人工林土壤細菌優勢門為變形菌門、放線菌門和酸桿菌門,與此前學者在呼倫貝爾地區的研究結果一致[12]。酸桿菌屬于典型的寡營養型細菌,而變形菌屬于富營養型細菌,適生于養分含量高的土壤環境[19]。因此,變形菌門和酸桿菌門相對豐度可以用作土壤質量評價[20]。人工林變形菌門相對豐度顯著高于沙質草地,酸桿菌門顯著低于沙質草地,這是由于人工林和沙質草地的環境條件明顯不同,人工林產生的大量枯落物以及根系分泌物能夠提高土壤養分含量,導致土壤細菌生活的微生境及養分需求有差異,進而改變細菌類群比例[21]。但隨林齡的增加,研究區沙地樟子松人工林中變形菌門相對豐度逐漸降低,這表明土壤養分狀況下降,可能與植被根系生物量的衰退以及凋落物分解速度降低有關[22]。此外,研究區厚壁菌門(Firmicutes)相對豐度僅為1%左右,與陜北沙化地區和內蒙荒漠草原存在較大差異[23- 24],說明不同的植被能夠通過凋落物輸入及根系分泌物等差異改變土壤環境,進而影響土壤細菌群落組成。
土壤細菌多樣性對提高土地生產力、增強脅迫環境下群落穩定性有著重要影響。研究區各林地土壤細菌群落多樣性無顯著差異,說明隨著沙地樟子松人工林林齡的增加,土壤細菌群落結構較為穩定。同時,人工林土壤表層細菌多樣性水平較高,這與其他學者的研究結果相似[25]。這是由于密集分布在土壤表層的樟子松根系及其分泌物能為細菌提供豐富的能源物質[26]。
4.2 呼倫貝爾沙地樟子松人工林土壤細菌群落的影響因素
土壤細菌群落結構與土壤理化性質緊密相關且相互影響。隨著人工林林齡的增加,全磷含量先增加再降低,土壤有機質和全氮含量逐漸下降,且林地全氮含量低于沙質草地,這與前人研究存在差異[27]。呼倫貝爾沙地位于寒溫帶大陸性半干旱地區,冬季漫長,植物生長期短,林地土壤表面養分含量與凋落物分解速率緊密相關。當沙地樟子松為20—30齡時,樟子松生長旺盛,凋落物量增加,土壤養分積累量增加,土壤養分含量升高[28];而后,隨著林木生長,樟子松生長變緩,植物凋落物和根系分泌物的養分歸還量小于植物的養分吸收量,沙地樟子松人工林土壤養分庫存在消耗的現象[22,29]。呼倫貝爾沙地樟子松人工林土壤細菌群落的主要影響因子為速效鉀、全磷和全氮,與前人研究結果相似[30- 31],說明土壤細菌群落結構與氮、磷、鉀等營養元素密切相關,土壤限制性資源對土壤細菌相對豐度有顯著影響。25 a和34 a人工林土壤細菌的主要影響因子為全氮,可能是由于該地土壤氮資源與林地初級生產力緊密相關[32],兩林地內全氮含量較高,充足的能源物質也有利于土壤細菌的存活和繁衍;43 a人工林土壤細菌的主要影響因子為土壤含水量,該林地的土壤含水量顯著低于其他樣地,較低的土壤水分含量會限制微生物的代謝功能以及呼吸底物的擴散,導致土壤呼吸作用減弱,并且水分脅迫還會通過改變根際環境來影響土壤細菌區系[33- 34]。此外,各林地0—10 cm土層細菌群落相對豐度與含水量呈正相關,10—20 cm土層中兩者呈負相關。10—20 cm土層土壤含水量較低,僅為表層的0.55—0.74倍,進一步說明了當土壤中水分含量較低時,土壤含水量會成為限制土壤細菌群落分布的主要因素。
4.3 呼倫貝爾沙地樟子松土壤細菌功能特征
植被對土壤細菌群落結構的影響會進一步影響土壤細菌功能組成[35]。呼倫貝爾沙地樟子松人工林土壤細菌主要涉及5個一級功能層和31個二級功能層。其中,土壤細菌代謝功能活躍且極為重要,如碳水化合物代謝與固氮、溶磷等作用密切相關,有益于植物的氮、磷循環[36- 37];氨基酸代謝可以幫助細菌吸收氨基酸,有利于細菌的存活與繁衍[38],部分土壤細菌類群還可以通過代謝產生抗生素、抑菌蛋白及促生長激素等,抑制病原菌并促進植物生長[39]。43 a人工林代謝功能基因相對豐度顯著高于其他林地,說明沙地樟子松人工林能夠提高土壤細菌代謝活動,從而促進自身對養分的吸收和利用。同時,研究區土壤細菌活躍的膜運輸功能有利于維持細菌各項生命活動,是土壤細菌正常發揮其功能的基礎[40]。沙質草地的部分功能基因相對豐度顯著高于25 a和34 a人工林,這些功能的活躍也許能使土壤細菌在較為嚴苛的環境中更好地吸收養分。此外,土壤細菌功能隨不同樣地的變化均表現在0—10 cm土層,說明土層深度對細菌功能基因組成的影響較大,表層土壤細菌功能對環境變化更為敏感。
5 結論
呼倫貝爾沙地樟子松人工林土壤細菌隸屬35門92綱109目210科267屬,主要優勢門為變形菌門、放線菌門和酸桿菌門。人工林變形菌門相對豐度顯著高于沙質草地,酸桿菌門相對豐度顯著低于沙質草地。不同林齡人工林土壤細菌群落多樣性無顯著差異。研究區沙地樟子松人工林土壤細菌群落結構的主要影響因子為速效鉀、全磷和全氮。沙地樟子松人工林土壤細菌群落主要涉及5個一級功能層和31個二級功能層,43 a人工林土壤細菌代謝功能活躍,有利于促進植物對養分的吸收和利用。沙地樟子松人工林的種植改善了土壤細菌群落結構,促進土壤細菌代謝功能。研究成果有助于深入理解沙地樟子松與土壤細菌的互饋關系,并可為沙地樟子松人工林可持續經營管理提供理論依據。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
