馬鈴薯腐爛莖線蟲生防細菌的篩選與鑒定
2021-07-11 07:09:46趙遠征邱廷艷高騰達高志超周洪友
中國農學通報 2021年18期
張 妞,王 東,趙遠征,項 鵬,邱廷艷,趙 鑫,高騰達,高志超,周洪友
(1內蒙古農業大學園藝與植物保護學院,呼和浩特010018;2內蒙古自治區農牧業科學院植物保護研究所,呼和浩特010031;3黑龍江省農業科學院黑河分院,黑龍江黑河164312;4內蒙古喀喇沁旗農牧局植保植檢站,內蒙古喀喇沁旗024409)
0 引言
馬鈴薯腐爛莖線蟲Ditylenchusdestructor,屬于遷移性植物內寄生線蟲,是國際公認的檢疫性線蟲[1-2]。該線蟲主要為害馬鈴薯和甘薯,侵染薯塊后,薯塊表皮皺縮、龜裂,內部褐色并逐漸變黑,后期伴有細菌和螨類二次侵染,導致整薯腐爛變質[3]。馬鈴薯腐爛莖線蟲病在北京、天津、山東、河北、河南、江蘇、新疆、甘肅等地均有發生,并以山東、河北兩省最為嚴重,已成為北方甘薯最嚴重的病害之一[4]。該病害可造成甘薯和馬鈴薯減產30%~50%,嚴重時可減產80%以上,甚至絕收[5-7],嚴重影響其產量和品質。目前利用生物防治措施控制植物線蟲病害已經取得了一定的研究進展,張國鋒等[8]研究表明,利用30 kg/hm2的厚孢輪枝菌(孢子2.5億個/g)和45 kg/hm2的淡紫擬青霉顆粒劑(5億/g孢子)可有效防治甘薯莖線蟲病,防治效果分別達70.94%和67.56%。高春梅等[9]研究表明叢枝菌根真菌(AMF)和暗隔內生真菌(DSE)組合菌劑抑制南方根結線蟲的發育,降低線蟲繁殖量、根內定殖數、發病率和根結指數。Lax等[10]研究發現接種根內球囊霉Glomus intraradices能有效地減輕南方根結線蟲Meloidogyne incognita對番茄根系的傷害,降低線蟲繁殖量。Noor等[11]研究發現,來源于馬鈴薯根及塊莖的內生細菌能夠抑制馬鈴薯胞囊線蟲的胞囊和二齡幼蟲(J2)形成,其中對胞囊數量的抑制率為51.7%~65.4%,對J2數量的抑制效果達到48.6%~76.4%。芽胞桿菌是用于防控植物線蟲病害的重要生防因子。有研究表明,蘇云金芽胞桿菌B.thuringiensis和巨大芽胞桿菌B.megaterium對馬鈴薯腐爛莖線蟲均具有較高的殺線活性[12-13]。Zeng等[14]分離獲得的芽胞桿菌菌株SMrs28的發酵液粗提物表現出較強的殺線蟲活性,其主要化合物對腐爛莖線蟲具有毒殺作用。Jonathand等[15]利用蠟狀芽胞桿菌B.cereus等根際細菌防治南方根結線蟲M.incognita,使根結指數由94%下降至25%~31%。生物防治具有對非靶標生物安全、致毒作用小、環境兼容性好、不產生抗藥性等優點,已成為防控腐爛莖線蟲病害的重要手段。本研究通過利用34個細菌菌株發酵液對馬鈴薯腐爛莖線蟲的殺線活性進行測定,篩選出具有高效觸殺活性的生防細菌菌株并明確其分類地位,為微生物殺線劑的研發與應用提供思路,旨在為生產上防治馬鈴薯腐爛莖線蟲病奠定基礎。
1 材料與方法
1.1 供試菌株、線蟲
供試細菌菌株共34個,-80℃條件下保存于內蒙古農業大學園藝與植保學院植物病理研究室;馬鈴薯腐爛莖線蟲D.destructor分離自發病的馬鈴薯病薯,并轉接于健康的馬鈴薯薯塊中進行擴繁,室溫條件下保存于內蒙古農業大學園藝與植保學院植物病理研究室。
1.2 主要試劑、藥品
細菌基因組DNA試劑盒,購自天根生化科技(北京)有限公司;EasyTaqPCR supermix,購自北京全式金生物技術有限公司;2×TaqPCR Mix購自生物工程大連有限公司;TAE緩沖液,購自購自上海生工生物工程有限公司;核酸染料,購自百泰克生物科技有限公司;瓊脂粉,購自上海致化化學科技有限公司;瓊脂,購自BioFroxx公司;葡萄糖、氯化鈉分析純,購自天津永晟精細化工有限公司;LP0021酵母浸粉,購自OXOID公司。
1.3 主要儀器
NSZ-606體視顯微鏡,寧波永新光學股份有限公司生產;5424離心機,德國Eppendorf公司生產;KGSX-500蒸汽滅菌鍋,日本Tomy Digital Biology公司生產;HWS-26電熱恒溫水浴鍋,上海-恒科學儀器有限公司生產;S1000 PCR擴增儀、Universal Hood II凝膠成像系統,美國BIO-RAD公司生產;JY-ECP3000電泳儀,北京君意東方電泳設備有限公司生產;XPH-160BSH-III恒溫恒濕箱,上海新苗醫療器械制造有限公司生產;JY2002電子天平,上海浦春計量儀器有限公司生產;EL20實驗室pH計,梅特勒-托利多儀器(上海)有限公司生產;Eco-Q30純水系統,上海和泰儀器有限公司生產;VS-840-U凈化工作臺,蘇州博萊爾凈化設備有限公司生產;3513型12孔細胞培養板,康寧生命科學(吳江)有限公司生產。
1.4 供試培養基
1.4.1 LB固態培養基 酵母提取物5 g、胰化蛋白胨10 g、NaCl 5 g、瓊脂15 g、蒸餾水1000 mL、pH 7.2。
1.4.2 LB液態培養基 酵母提取物5 g、胰化蛋白胨10 g、NaCl 5 g、蒸餾水1000 mL、pH 7.2。
1.4.3 碳源利用基礎培養基 KH2PO42.38 g、K2HPO4·3H2O 5.65 g、(NH4)2SO42.64 g、CuSO45H2O 6.4 mg、MgSO4·7H2O 1 g、ZnSO4·7H2O 1.5 mg、MnCl2·7H2O 7.9 mg、FeSO4·7H2O 1.1 mg、瓊脂15 g、蒸餾水1000 mL,pH 7.2。
1.4.4 氮源利用基礎培養基 葡萄糖10 g、K2HPO4·3H2O1g、MgSO4·7H2O5g、NaCl5g、FeSO4·7H2O10mg、瓊脂15 g、蒸餾水1000 mL、pH 7.2。
1.4.5 淀粉水解培養基 可溶性淀粉10 g、K2HPO4·3H2O 0.3 g、MgCO30.3 g、KNO31 g、NaCl 0.5 g、瓊脂15 g、蒸餾水1000 mL,pH 7.2。
1.4.6 V-P測定液態培養基 胰化蛋白胨7 g、葡萄糖5 g、K2HPO45 g、蒸餾水1000 mL,pH 7.2。
1.4.7 接觸酶培養基 胰化蛋白胨20 g、NaHPO42 g、葡萄糖10 g、瓊脂15 g、蒸餾水1000 mL,pH 7.2。
1.4.8 H2S產生培養基 胰化蛋白胨2.5 g、NaCl 5 g、酵母提取物7.5 g、瓊膠120 g、蒸餾水1000 mL,pH 7.2。
1.4.9 吲哚產生液態培養基 胰化蛋白胨20 g、NaHPO42 g、葡萄糖10 g、蒸餾水1000 mL,pH 7.2。
1.4.10 硝酸鹽液態培養基 胰化蛋白胨10 g、KNO31 g、蒸餾水1000 mL,pH 7.2。
1.4.11 明膠培養基 葡萄糖20 g、蛋白胨5 g、明膠200 g、蒸餾水1000 mL。
1.5 線蟲分離與懸浮液制備
采用改良貝曼漏斗法[16-17]從馬鈴薯病薯中分離馬鈴薯腐爛莖線蟲,用吸管將分離到的線蟲轉移至10 mL離心管中,加入35%的蔗糖溶液后3500 r/min離心2 min;取上層線蟲懸浮液,轉移至無菌水中,4000 r/min離心5 min;棄上清液,用0.5%的次氯酸鈉消毒1 min后,4000 r/min離心5 min;棄上清液,加入0.5%硫酸鏈霉素和0.5%青霉素的混合液中消毒6 min,離心后吸出消毒液,用無菌水沖洗3次,線蟲懸浮液(約1000條/mL),4℃保存備用[18]。
1.6 菌株發酵液的制備
將34個菌株于LB培養基平板上進行活化,利用9 mm打孔器打成菌餅放入250 mL三角瓶,每瓶裝入100 mL液體培養基中,25℃~28℃下,150 ~180 r/min搖床發酵2~3天后將菌株的發酵液6000 r/min離心5 min,取上清液備用。
1.7 菌株發酵液對馬鈴薯腐爛莖線蟲的室內觸殺活性篩選
采用直接觸殺法[19],將馬鈴薯腐爛莖線蟲懸浮液分裝在2 mL離心管中,每管約含100條線蟲,5000 r/min離心1 min,棄上清液,用移液槍將菌株發酵液上清液先加入2 mL離心管中,每個離心管中加入1.5 mL菌液,在微型漩渦混合儀充分震蕩,每個處理重復3次,以無菌水為對照。用橡皮筋固定,放入無菌聚乙烯塑料袋中,25℃恒溫培養48 h后在顯微鏡下進行觀察,記錄馬鈴薯腐爛莖線蟲的總數以及死亡數。
計算馬鈴薯腐爛莖線蟲的死亡率公式(1)和校正死亡率公式(2),并使用IBM SPSS Statistics 24.0進行數據進行顯著性分析。
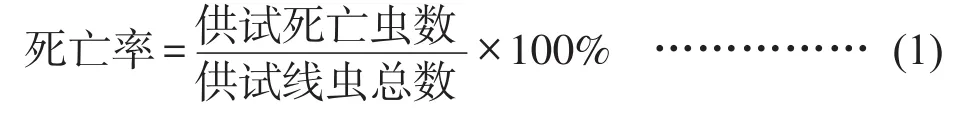
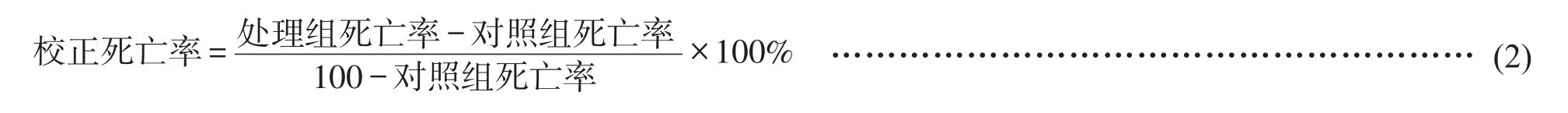
1.8 菌株鑒定
1.8.1 菌株生理生化鑒定 參考《伯杰細菌鑒定手冊》和《常見細菌系統鑒定手冊》對菌株生理生化特征進行鑒定,主要有碳素化合物的利用:L-阿拉伯糖、D-木糖、葡萄糖、D-山梨醇、蔗糖、i-肌醇、水楊苷、V.P.試驗;氮素化合物的利用:硝酸鉀、硝酸銨、L-羥脯氨酸、硝酸鹽還原、H2S的產生、吲哚的產生;其他反應測定:淀粉水解、接觸酶、明膠液化[20-22]。
1.8.2 菌株的16S rDNA鑒定 篩選獲得的細菌菌株在LB固體培養基上劃線,挑取單菌落放入裝有LB液體培養基的10 mL離心管中,28℃、180 r/min搖培24 h。采用細菌基因組DNA提取試劑盒,參考使用說明書進行DNA提取,-20℃保存備用。利用16S通用引物27F(5’-AGAGTTTGATCCTGGCTCAG-3’)和 1492R(5’-GGTTACCTTGTTACG-ACTT-3’)對菌株 16 S rDNA片段進行PCR擴增。PCR擴增體系25 μL:模板DNA 3 μL、dd H2O 7.5 μL,2×TaqPCR Mix 12.5 μL,上下游引物各1 μL。反應條件為:94℃ 3 min,1個循環;94℃30 s,55℃ 1 min,72℃ 90 s,29個循環;72℃ 10 min,1個循環。PCR產物經1%瓊脂糖凝膠電泳檢測合格后,送往華大基因科技股份有限公司進行測序,獲得序列經NCBI數據庫在線BLAST比對分析,并利用MEGA 7.0軟件中的鄰接法構建系統發育樹。
2 結果與分析
2.1 不同細菌菌株發酵液對馬鈴薯腐爛莖線蟲的觸殺活性結果
不同細菌菌株發酵液對馬鈴薯腐爛莖線蟲的觸殺活性見表1。表1可知,34個細菌菌株發酵液處理馬鈴薯腐爛莖線蟲48 h后,對該線蟲均表現出一定的致死作用。其中菌株Chi9-7與Xia4-11發酵液對馬鈴薯腐爛莖線蟲的校正死亡率在80%以上,菌株Xia4-11發酵液對馬鈴薯腐爛莖線蟲致死作用最強,校正死亡率達82.59%;菌株Nong3-5、Z-1、Gu4-7、b5-4、b1-3、Nong5-6和Chi9-2發酵液對馬鈴薯腐爛莖線蟲的校正死亡率為72.36%~77.94%,且不同菌株間觸殺作用差異并不顯著;另外菌株 X1''、Chitu8-4、Gen7-3、S-6、Gen9-3、Gen6-10和CF4-51發酵液對馬鈴薯腐爛莖線蟲的校正死亡率為60.53%~69.16%;其余菌株對馬鈴薯腐爛莖線蟲的致死作用較差,校正死亡率均在24.49%~59.42%之間,其中菌株Chi1-15、3-6、C-11發酵液對馬鈴薯腐爛莖線蟲正死亡率分別為僅為24.49%、27.04%、29.51%。
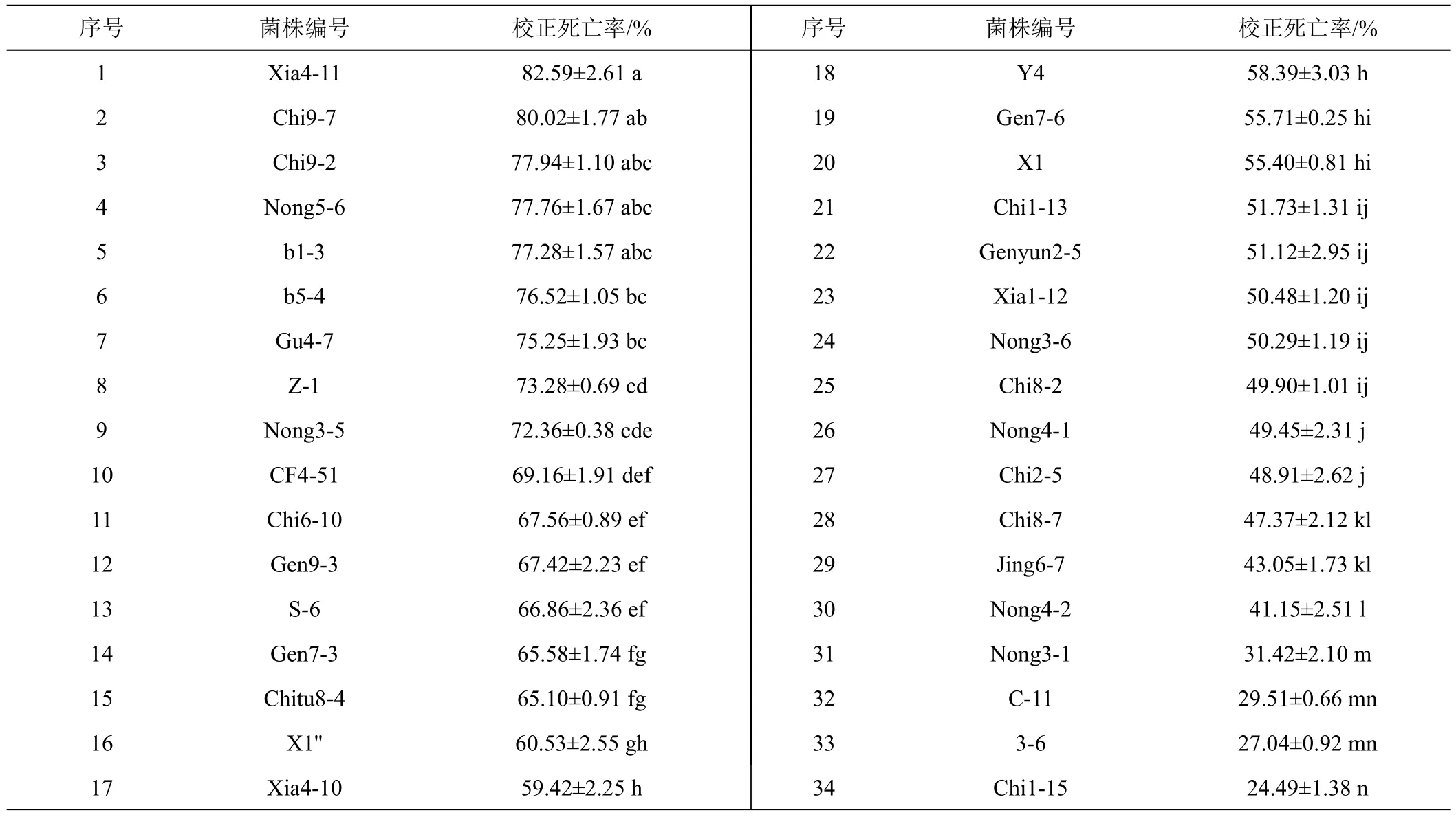
表1 不同菌株發酵液對馬鈴薯腐爛莖線蟲48 h的觸殺活性
2.2 菌株的鑒定
2.2.1 形態鑒定 9個細菌菌株菌落形態見圖1、表2。圖1、表2可知,對馬鈴薯腐爛莖線蟲校正死亡率大于72.36%的9個細菌菌株進行鑒定,9個菌株接種于LB固態培養基后,于34 h時觀察各個菌株的具體形態特征及生長情況發現菌株Xia4-11、Chi9-2、b5-4、Nong3-5、Chi9-7和Z-1單菌落直徑3~4 mm,Nong5-6、b1-3和Gu4-7單菌落直徑7~8.6 mm,其中菌株Gu4-7生長速度最快34 h時8.6 mm,Nong5-6和b1-3生長速度較快,分別為7 mm和7.5 mm,其余菌株生長較慢菌落直徑3~4 mm;9個菌株中Chi9-2菌落邊緣形狀為圓形,其余菌株均不規則;菌株Xia4-11、Chi9-2、Nong3-5、Nong5-6和Chi9-7的單菌落形態平滑,其余菌株但菌落形態隆起;菌株b1-3菌落顏色為乳白色,其余菌株顏色均為淡黃色;菌株Chi9-2、Z-1和Gu4-7菌株質地濕潤,Nong3-5、Xia4-11、Nong5-6、Chi9-7菌株質地干燥,其余菌株質地粘稠;菌株Xia4-11和Z-1菌株氣味分別為腥味和臭味,其余7個菌株均無味,如圖1、表2所示。
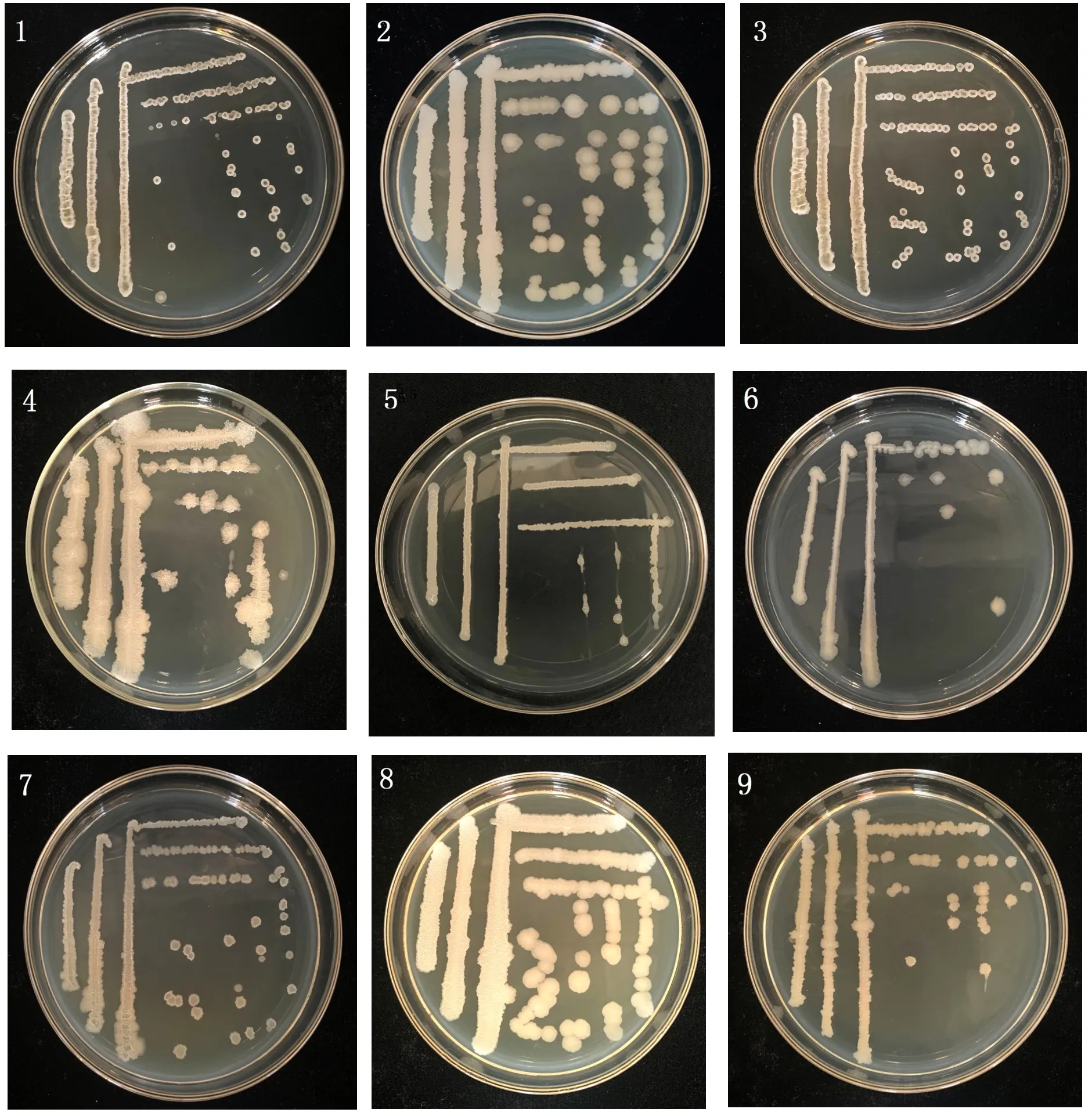
圖1 9個細菌菌株菌落形態
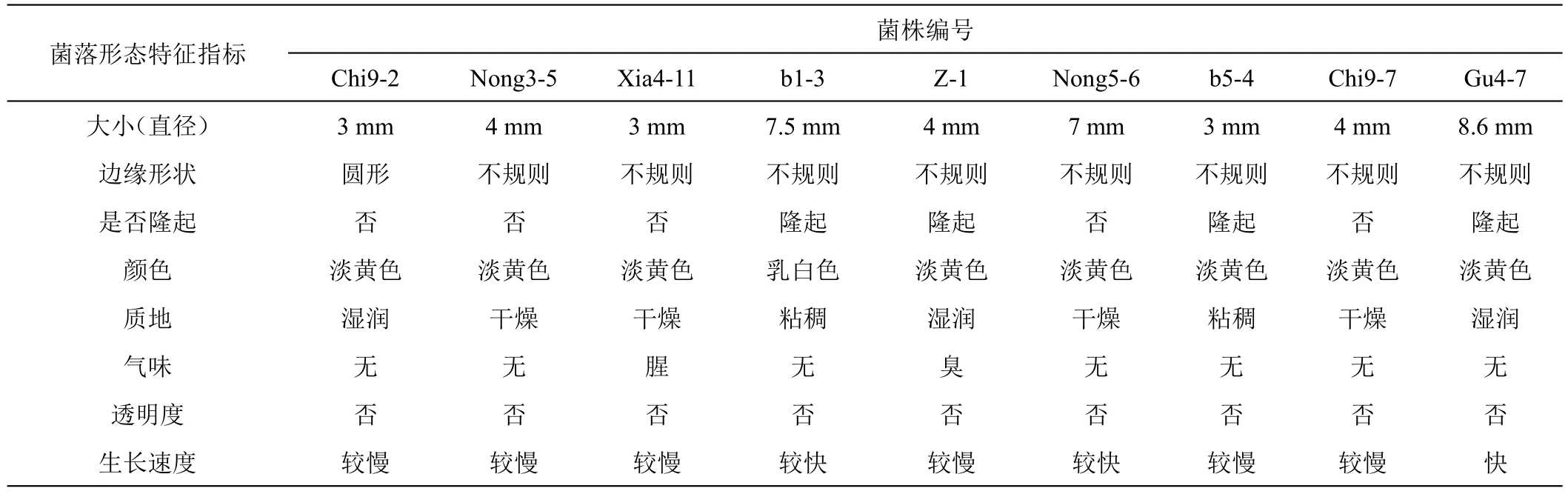
表2 9個細菌菌株菌落形態特征
2.2.2 生理生化特征鑒定 9個細菌菌株生理生化特征見表3。表3可知,在碳源利用上9個菌株均能夠利用D-木糖、葡萄糖、蔗糖、D-山梨醇;除菌株Z-1和Gu4-7外,其余7個菌株均能利用L-阿拉伯糖;除菌株Gu4-7外,其余8個菌株均能利用i-肌醇;菌株b5-4、Nong5-6和Gu4-7不能利用水楊苷,但其余6株菌均能利用其生長;在氮源利用上,9個菌株均能夠利用硝酸銨,除菌株b5-4外,其余8株菌均能利用硝酸鉀;此外,淀粉水解、V-P測定、接觸酶、硝酸鹽還原、明膠液化、H2S產生、吲哚試驗中9個株菌均為陽性。
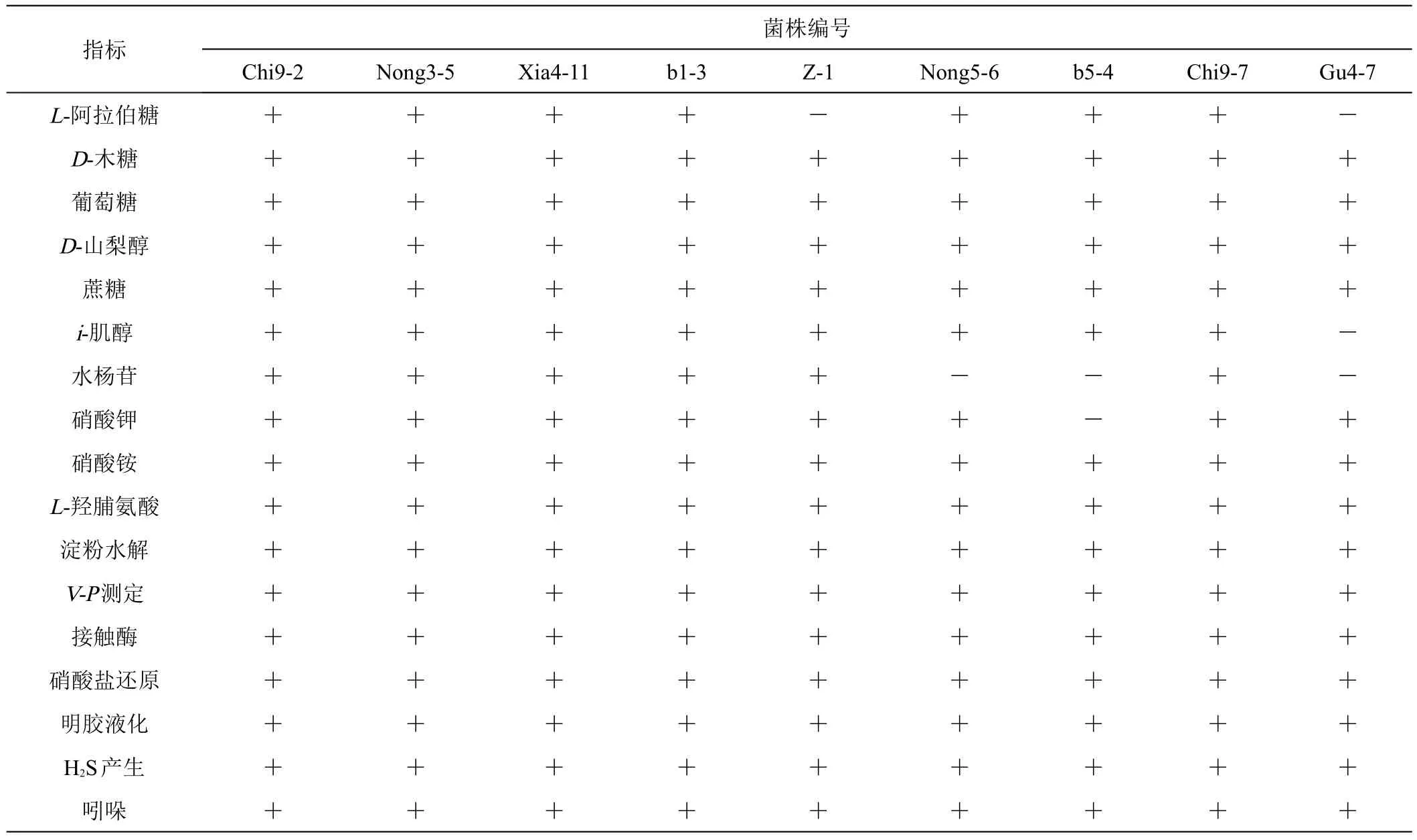
表3 9個細菌菌株生理生化特征
2.2.3 16S rDNA分子鑒定 利用16S rDNA對篩選獲得的9個細菌菌株進行分子鑒定結果見圖2~3。由圖2可知,PCR擴增得到的片段大小為1500 bp左右。由圖3可知,菌株Chi9-7、Nong3-5、Nong5-6、b1-3與枯草芽胞桿菌Bacilluicssubtilis處于同一分支,Chi9-7、Nong3-5與B.subtilis(登錄號:MT590663.1)相似性均為100%,Nong5-6與B.subtilis(登錄號:KC1978.5.1)相似性為99.79%,b1-3與B.subtilis(登錄號:MT487684.1)相似性為99.93%;b5-4菌株與貝萊斯芽胞桿菌B.velezensis屬于同一分支,相似性為100%;Chi9-2、Xia4-11、Gu4-7和Z-1與解淀粉芽胞桿菌B.amyloliquefaciens屬于同一分支,Chi9-2與B.amyloliquefaciens(登錄號:MT613661.1)相似性 為100%,Xia4-11、Gu4-7和Z-1與B.amyloliquefaciens(登錄號:KC417346.1)相似性分別為99.93%、99.72%和99.86%。
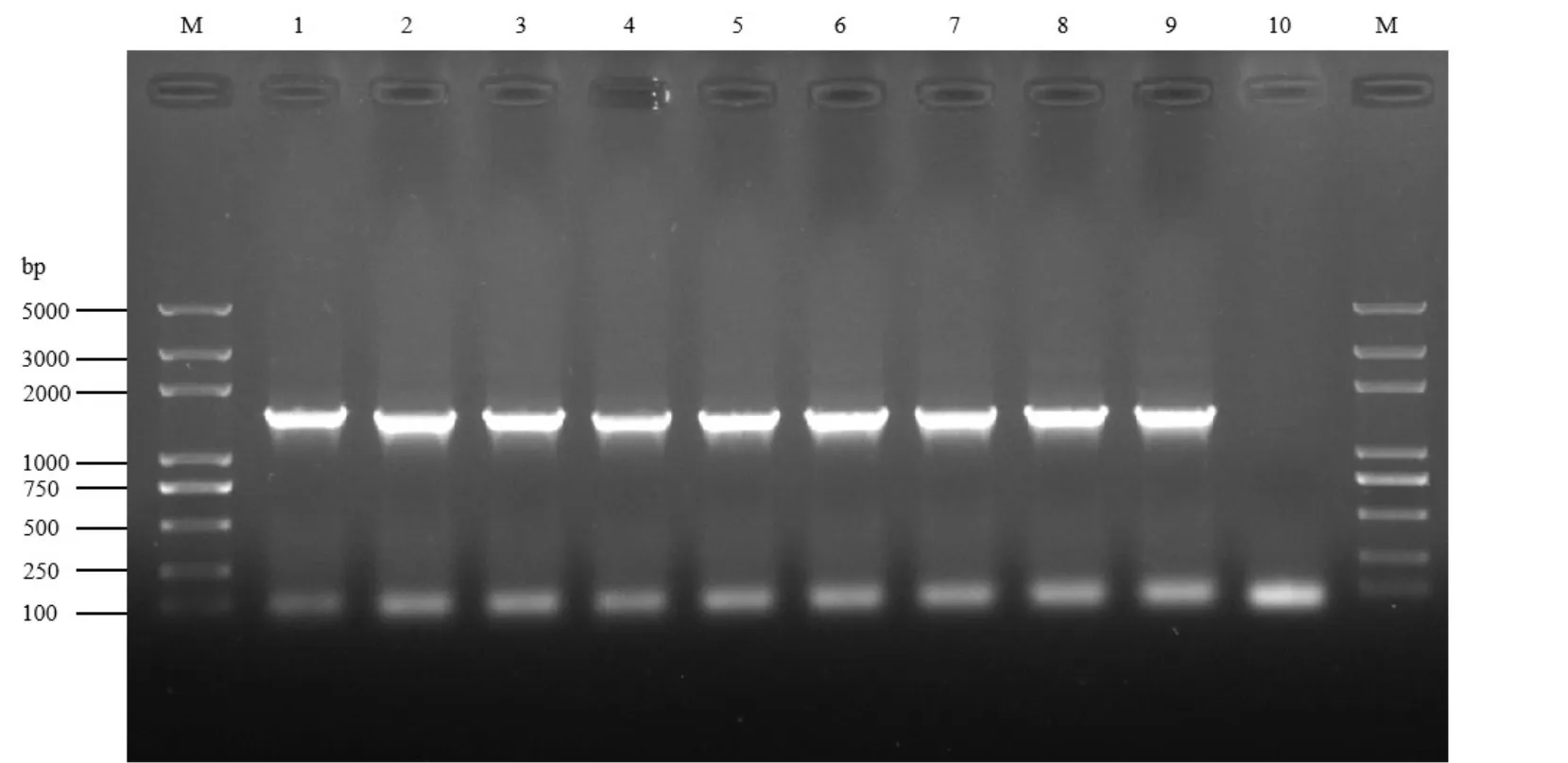
圖2 9個細菌菌株16S rDNA的PCR擴增結果
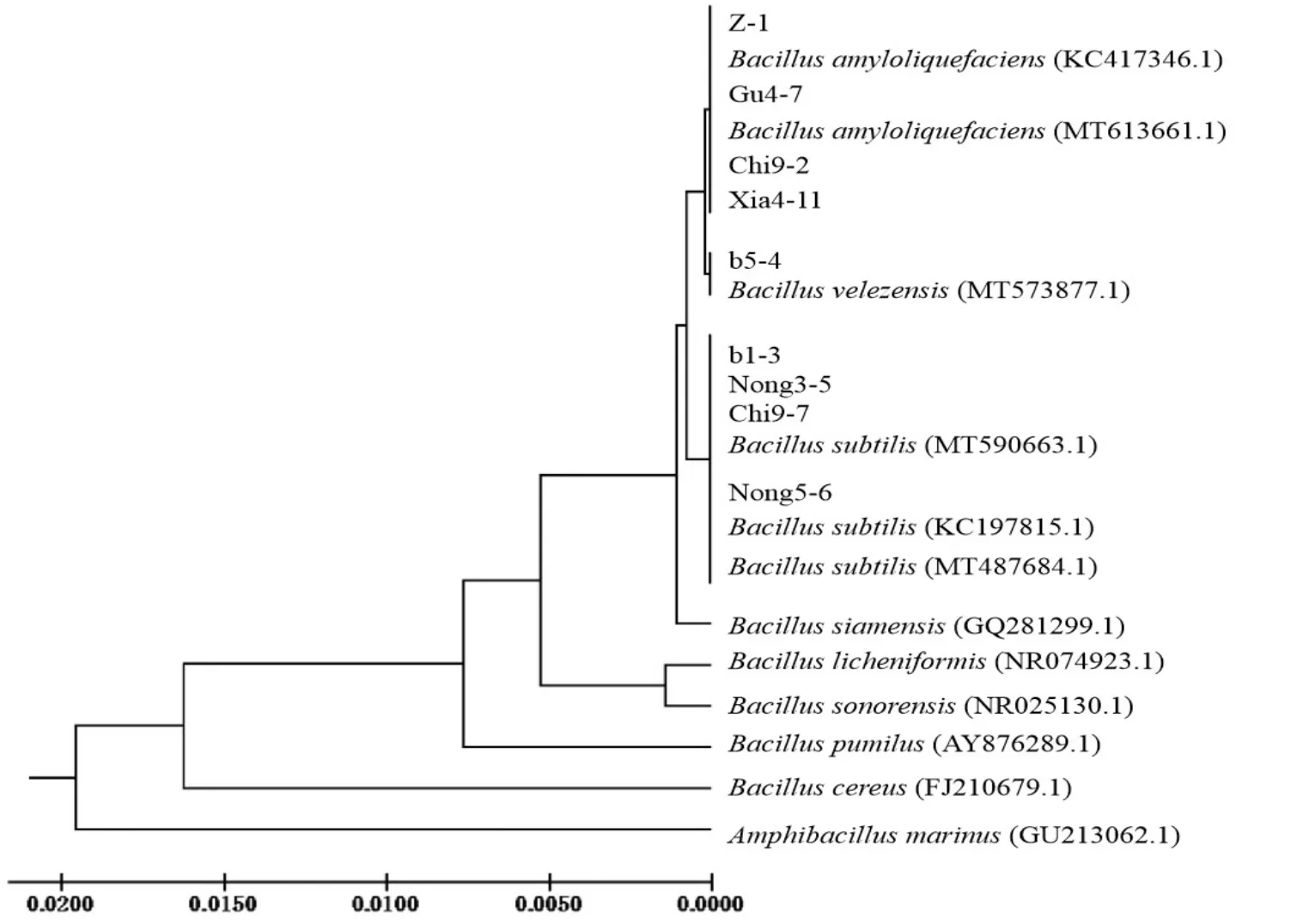
圖3 9個細菌菌株系統進化樹
經細菌16S rDNA分子鑒定,并結合各菌株的形態特征和生理生化特性,將9個供試菌株分別鑒定為菌株Chi9-7、Nong3-5、Nong5-6和b1-3為枯草芽胞桿菌(B.subtilis),菌株Chi9-2、Xia4-11、Gu4-7和Z-1為解淀粉芽胞桿菌(B.amyloliquefaciens),菌株B5-4為貝萊斯芽胞桿菌(B.velezensis)。
3 討論與結論
芽胞桿菌(Bacillusspp.)作為重要的生防因子,廣泛應用于農作物的病害防控,是當前的研究熱點[23],已在植物線蟲病害的生物防治領域發揮著重要作用。近年來,研究人員篩選出多個解淀粉芽胞桿菌(B.amyloliquefaciens)菌株對植物線蟲具有較高的抑制活性。例如,蔣盼盼等[24]室內研究發現,解淀粉芽胞桿菌菌株B1619粗提液原液分別處理蔬菜根結線蟲,其24、48 h的線蟲死亡率分別達78.8481%和82.4923%。Zhu等[25]研究發現解淀粉芽胞桿菌JK-JS3培養原液濾液對松材線蟲、擬松材線蟲、大核滑刃線蟲、霍夫曼尼傘滑刃線蟲、李氏長尾線蟲、吳氏長尾線蟲和外滑刃科線蟲共7種侵染松樹的植物線蟲具有一定殺線活性;在此基礎上,朱麗梅等[26]對解淀粉芽胞桿菌JK-JS3分泌的殺線蟲活性物質進行了研究,發現該物質在堿性條件下殺線活性較高,且在常溫25℃和冷藏4℃條件下保存60天后仍具有較高的殺線活性。結合前人的研究結果表明,解淀粉芽胞桿菌對于多種植物線蟲均具有較強的殺線活性。但是,對于馬鈴薯腐爛莖線蟲,往往比其他植物線蟲具有更高的抗逆性[27]。因此,篩選出對馬鈴薯腐爛莖線蟲具有高效抑制活性的生防菌株難度較大。夏彥飛[28]研究發現解淀粉芽胞桿菌FZB42和B3培養濾液處理馬鈴薯腐爛莖線蟲48 h的抑制率分別達到28%和65%;而本研究篩選出4株解淀粉芽胞桿菌Xia4-11、Chi9-2、Gu4-7和Z-1,具有更強的觸殺活性,對馬鈴薯腐爛莖線蟲48 h的抑制率達到73.28%~82.59%。一般來說,理想的生防微生物除了要具有較高的抑病活性,在田間環境中的定殖能力往往影響其生防效果,因此今后有必要將繼續開展田間試驗,從而進一步篩選獲得更具開發潛力和應用前景的生防菌株。
研究表明,枯草芽胞桿菌B.subtilis也對多種植物線蟲具有較高的抑制活性。如夏彥飛等[29]發現,枯草芽胞桿菌OKB105發酵液處理南方根結線蟲、擬松材線蟲、豆傘滑刃線蟲和程氏傘滑刃線蟲,4種線蟲24 h的死亡率均達到100%。陶樹興等[30]通過盆栽試驗發現,枯草芽胞桿菌2-3-2發酵液對黃瓜根結線蟲病防效達89.41%,且明顯增加植株干重。丁國春等[31]研究發現,枯草芽胞桿菌AR11稀釋菌懸液對南方根結線蟲的二齡幼蟲有抑制作用,且高濃度菌懸液可以強烈抑制卵的孵化,同時對番茄具有明顯的促生作用。對于馬鈴薯腐爛莖線蟲,劉瑋瑋[32]研究發現枯草芽胞桿菌BL02和G8產生的揮發物能夠對該線蟲產生強烈的抑制作用,經菌株揮發物處理后大部分線蟲在1~12 h內活動逐漸減弱,24 h后完全停止活動。本研究利用直接觸殺的測定方法,發現枯草芽胞桿菌Chi9-7、Nong5-6、b1-3和Nong3-5發酵液對馬鈴薯腐爛莖線蟲具有明顯的抑制活性,48 h抑制率可達到72.36%~80.02%;但在試驗過程中這些菌株是否產生揮發性物質并對該線蟲產生熏蒸活性,還有待于后續試驗的驗證。
貝萊斯芽胞桿菌B.velezensis是一種新型的生防因子,具有廣譜抑菌活性和促進植物生長的作用,在農業生產中極具開發潛力[33]。張文博等[34]研究發現貝萊斯芽胞桿菌FZB42稀釋培養液處理松材線蟲48 h的抑制率達50%。茆振川等[35]研究發現貝萊斯芽胞桿菌Bv-25對根結線蟲病具有良好的防治效果,利用該菌株進行病害的早期防控,從而有助于實現蔬菜的安全和無公害生產。針對于馬鈴薯腐爛莖線蟲,之前尚未檢索到具有較高抑制活性的貝萊斯芽胞桿菌菌株;而本研究篩選了一株貝萊斯芽胞桿菌b5-4,對馬鈴薯腐爛莖線蟲具有較強的觸殺效果,48 h時線蟲的校正死亡率達76.52%,這一發現填補了相關領域的空白。生防微生物資源十分豐富,蘊藏其中的代謝產物可以為新型殺線劑的研發提供新的思路。作為新型生防因子,貝萊斯芽胞桿菌b5-4抑制馬鈴薯腐爛莖線蟲的機制有待于深入研究。
本研究以馬鈴薯腐爛莖線蟲為靶標,從34個細菌菌株中篩選出9株芽胞桿菌,表現出較高的觸殺活性,其48 h的抑制率達到72.36%~82.59%;進而利用16S rDNA基因,并結合菌落形態和生理生化特征,鑒定出4株解淀粉芽胞桿菌、4株枯草芽胞桿菌和1株貝萊斯芽胞桿菌。本研究通過篩選高效殺線蟲生防菌,為微生物殺線劑的研發提供了新資源,也為馬鈴薯腐爛莖線蟲病的有效防控奠定了基礎。
