小蘇干湖濕地典型泌鹽植物海乳草葉性狀和光合特性對淹水的響應
2021-07-09 05:07:12李子琴趙成章趙夏緯茍芳珍馬俊逸
生態學報 2021年11期
關鍵詞:植物
李子琴, 趙成章, 趙夏緯, 李 群, 茍芳珍, 任 杰, 馬俊逸
西北師范大學地理與環境科學學院,甘肅省濕地資源保護與產業發展工程研究中心, 蘭州 730070
植物是一個功能和結構平衡的生命體[1- 2],為了提高自身的生境適合度,植物往往通過權衡葉片各功能性狀間的資源分配策略,形成了與環境相適應的葉片構型及其光合特性[3]。葉片是植物能量和物質轉化的重要場所,葉片功能性狀能夠反映植物在不同生境下植株生長的調整策略以及對所吸收光能的利用水平,體現了植物在葉片光截獲、碳同化和水分利用等方面的生態策略[4- 6]。葉面積作為衡量葉片光合能力大小的重要指標[7],影響植物與外界環境氣體、能量等的交換[4];葉厚度在植物生長方面扮演著重要角色,通過調控葉肉細胞中柵欄組織和氣孔的構建模式,影響著植物的碳同化產物生產和積累效率[8]。有研究表明植物葉片面積、厚度與葉片光合效率關系十分密切,葉片可以通過權衡葉面積、葉厚度之間的生物量分配機制,實現不同環境條件下植物基礎功能的有效發揮和植物光合作用的正常進行[3]。蒸騰速率和凈光合速率作為衡量光合能力的重要指標,不僅能反應植物通過調節自身水分損耗適應不同環境的能力[9],而且可反映特定生境中植物的資源利用效率[10]。植物能夠通過葉片功能性狀在形態和結構上的可塑性,改變葉片的光合生理特征實現植物葉片光合效率的最大化,反映了植物與環境協同適應的生存策略[10-12]。淹水條件是內陸鹽沼濕地植物最常見的逆境脅迫因子,季節性淹水所引起的土壤水鹽含量空間分異格局,能夠營造差異化的群落環境,迫使植物調整葉面積、葉厚度等葉片功能性狀間的資源分配模式,差異化地調控葉片凈光合速率、蒸騰作用等生理生態功能,最大限度地適應外界環境因子的變化,維持植物的正常生理代謝水平,增強植物種群的生長發育速度和更新能力[13-15]。因此,研究鹽沼濕地植物光合特性與葉性狀在不同淹水生境下的響應變化,有助于理解內陸鹽沼濕地植物葉片功能性狀對異質性生境的響應機制。
海乳草(GlauxmaritimaL)屬報春花科多年生草本植物,喜生于土壤潮濕或地勢低洼積水的生境,以及土壤輕中度鹽漬化的低濕草甸、荒漠、鹽堿地和沼澤草甸等,是典型的濕生泌鹽植物。目前學術界圍繞植物光合特性與環境因子的關系[16-17]、各生態型植物的耐鹽差異性[18]、鹽脅迫對光合指標的影響[19]、不同水鹽梯度對植物光合特性的影響[20-21]以及濕地植物功能性狀[22]等問題展開了廣泛研究,有關海乳草在不同鹽濃度環境中的葉片形態結構特征[23]和葉片形態解剖學特征[24]也得到了一定重視,但是有關水鹽空間異質條件下濕地泌鹽植物海乳草葉性狀與光合生理特性的變化規律還尚不明確。鑒于此,本研究以甘肅省小蘇干湖濕地的海乳草為研究對象,通過分析3種淹水梯度下海乳草光合特征與葉性狀的響應變化,試圖明晰:(1)不同淹水梯度下的葉性狀與光合特性是否存在差異性?(2)形成這種差異性的原因有哪些?旨在揭示植物在不同生境條件下的光合作用過程及生態適應機制。
1 材料與方法
1.1 研究區和樣地概況
研究區位于阿克塞小蘇干湖自然保護區,處于青藏高原柴達木盆地北緣阿爾金山、黨河南山、塞什騰山和吐爾根達坂山之間的花海子高原盆地,地理位置為39°01′25″—39°05′32″N,94°10′33″—94°14′43″E,海拔2807—2808 m,屬典型的內陸高寒半干旱氣候,年均氣溫< -0.4℃,1月份平均氣溫為-14℃,7月份平均氣溫為13.8℃,年均降水量77.6 mm,蒸發量1964.8 mm,年日照時數3100—3500 h,干燥度30,無霜期90 d左右。蘇干湖水系屬柴達木內流河水系,土壤主要以草甸土、草甸沼澤土和鹽土為主,植被在應對寒冷、干旱和土壤鹽漬化等生境條件,形成了植株矮小、根系發達且具深根性和泌鹽功能的區域性生態特征,主要植物有珠芽蓼(Polygonumviviparum)、海乳草(Glauxmaritima)、堿蓬(Suaedaglauca)、蘆葦(Phragmitesaustralis)、芨芨草(Achnatherumsplendens)、賴草(Leymussecalinus)、海韭菜(Triglochinpalustre)、草地早熟禾(Poapratensis)、水麥冬(Triglochinmaritimum)、鹽角草(Salicorniaeuropaea)等。
1.2 實驗方法
小蘇干湖是一個具有出口的微咸水湖,蘇干湖周圍鹽沼濕地的水淹頻率每年一次,每年11月至次年4月濕地表面有靜水持留,5月回歸,其它時間濕地暴露在空氣中,根據自然保護區20多年來的水文記錄,湖水泛濫區寬度650—715 m左右。為便于實驗的調查研究,2018年8月中下旬,在小蘇干湖北邊地勢平緩區域從湖岸邊緣向多年的潮水線末端設置1條寬50 m,長350 m的平行樣帶,該樣帶海乳草為濕地群落的主要植物;依據歷年小蘇干湖洪水泛濫和退水痕跡,測算的鹽沼濕地淹水時長,將該樣帶劃分為3個樣地(I、II和III),分別如下:(1)樣地I為輕度淹水區,離湖岸220—350 m,靜水持留時間:當年11月下旬—次年2月上旬,60—90 d,土壤鹽漬化嚴重,海乳草植株矮小,莖呈匍匐狀,葉片肉質化,堿地風毛菊和早熟禾為共同優勢種,海乳草、鹽角草伴生呈片狀分布;(2)樣地II為中度淹水區,離湖岸50—220 m,靜水持留時間:當年11月下旬—次年4月上旬,90—150 d,土壤鹽漬化較輕,葉片輕度肉質化,海乳草為群落優勢種,堿地風毛菊和鹽角草零星分布;(3)樣地III為重度淹水區,離湖岸0—50 m,靜水持留時間:當年11月下旬—次年6月上旬,淹水150—210 d,以泥炭土為主,土壤鹽漬化程度低,植被覆蓋度較高,海乳草莖稈直立,葉片肉質化不明顯,海乳草和早熟禾為群落的共同優勢種。在每個樣地內設置2個10 m×10 m的實驗樣方,共計6個固定實驗樣方。
(1)濕地群落學調查:在每個實驗樣方內用交叉對角線法選取6個1 m×1 m的小樣方,分別測量樣方內全部植物的高度、蓋度、密度和地上生物量;每個樣方內隨機選取6株海乳草,齊地剪取放入信封袋中帶回實驗室進行其他數據測量。
(2)光合特征值測定:在天氣晴朗的上午10:00—11:00測定海乳草的光合特征,使用GFS- 3000便攜式光合測量系統(Heinz Walz GmbH, Bavaria, Germany),測量時使用人工紅藍光源,將光合有效輻射(Photosynthetically active radiation, PAR)、CO2濃度、流速和相對濕度(Relative humidity,RH)分別設置為1200 μmol m-2s-1、340 μmol/mol、750 μmol/s和40%—50%,從樣地隨機選取5株海乳草,在每個植株上選取4片健康成熟的葉片分別測定凈光合速率、蒸騰速率等參數,并計算水分利用效率(Water use efficiency, WUE=Pn/Tr)。
(3)葉性狀測定:使用100分度游標卡尺避開葉脈,在10片供試葉片不同部位測量5次,取平均值為該植株葉厚度。將采集的每株海乳草從莖基部向上隨機選取10片健康的葉片,擦干凈后用便攜式激光葉面積儀(CI- 202, Walz, Camas, USA)測定每片葉子的面積并記錄,取平均值為該植株的單葉葉面積,將測完葉面積的葉片裝入信封放入烘箱80℃烘干,48 h以后取出秤重,并計算比葉面積。
(4)葉綠素測定:使用葉綠素計(SPAD- 502, Minolta, Japan),測量時盡量避開葉片葉脈部分,在每個葉片的不同部位測量5次,取平均值作為該個體的葉綠素含量。
(5)光合有效輻射測定:選擇晴朗無云的天氣,于9:00—11:00對3個生境中的海乳草植株上方、中層和地表,用手持光量子計(3415F, Walz, Plainfield, USA)進行光合有效輻射測定。
(6)土壤含水量測定:在濕地群落學調查樣方內隨機挖掘1 m×1 m×0.5 m土壤剖面,用環刀(200 cm3)分5層間隔10 cm采取土樣,重復3次,現場編號、各土層均勻混合稱鮮質量,然后帶回實驗室在105 ℃的烘箱內烘12 h,取出稱質量,計算出各樣地0—50 cm土層土壤質量含水量。
(7)土壤鹽分和電導率測定:在每個采樣點用土鉆(內徑35 mm)以10 cm為間隔分3層采集地下30 cm土樣(每份土樣濕重不少于600 g),除去地上凋謝物和石塊等雜質后帶回室內,用電導法測定土壤含鹽量[25]。
1.3 數據處理
不同樣地海乳草葉面積、葉厚度、凈光合速率和蒸騰速率進行以10為底的對數轉換,使之符合正態分布后采用線性回歸方法分析(Regression Analysis),采用單因素方差分析(ANOVA)比較不同梯度間海乳草的葉面積、葉厚度和葉綠素等功能性狀,實驗數據采用SPSS 22.0軟件進行處理分析,用Excel 2013軟件繪圖。
2 結果分析
2.1 不同樣地的群落生物學特征和土壤理化性質
受淹水時間長短的影響,3個樣地的土壤含水量和電導率均呈現出顯著的差異性(P< 0.05, 表1)。樣地(I)土壤含水量最小、電導率和PAR最大,植物受干旱與鹽堿脅迫的影響蓋度和密度處于較低水平,高度和地上生物量呈適中狀態;樣地(III)土壤含水量最大、電導率最小,群落光照充足,植物具有充足的水源供給,土壤鹽堿化程度較弱以泥炭土為主,濕度群落的高度處于最高水平比樣地(II)增加了309.44%,其蓋度、密度和地上生物量也均達到最高水平,分別比樣地(I)增加了32.97%、21.08%和132.01%;樣地(II)處于過渡帶,土壤含水量和電導率,濕地群落的密度和蓋度都呈適中狀態,高度和地上生物量則達到最低水平。
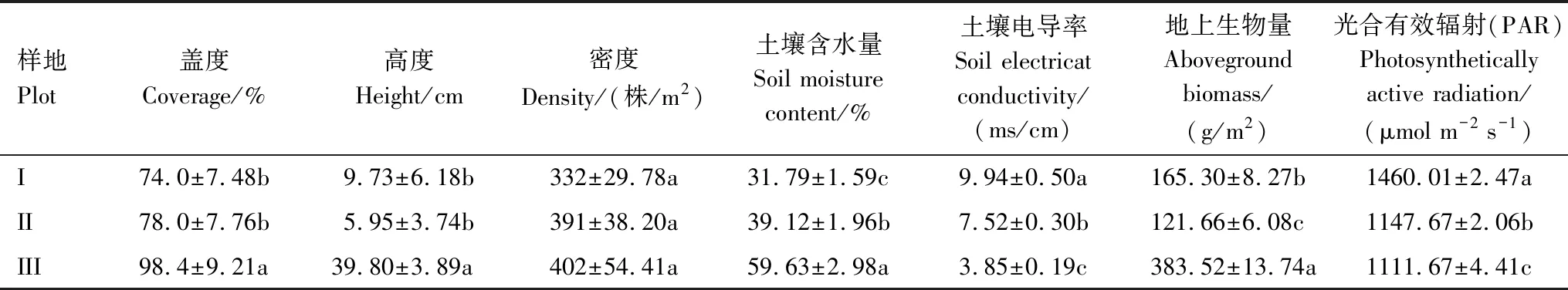
表1 不同濕地群落的生物學特征和土壤特性(數值為平均值±標準誤差)
2.2 海乳草葉性狀分析
不同生境下海乳草的葉性狀的變化如表2所示。隨著土壤含水量的增加和含鹽量的降低,海乳草葉片的葉面積、比葉面積呈逐漸增大的趨勢,樣地III比樣地I分別增加了40.77%、45.36%,樣地II與樣地I無顯著差異(P>0.05);海乳草的葉厚度則呈逐漸減小的趨勢,從樣地I到樣地III減小了24.01%,樣地II與樣地I也無顯著差異(P>0.05);海乳草的葉干重呈先增大后減小的趨勢,極大值為樣地II,但樣地I,II,III之間無顯著差異(P>0.05);海乳草的葉綠素呈逐漸減小的趨勢,由樣地(I)到樣地(III)減小了48.62%。

表2 不同樣地海乳草葉性狀特征(數值為平均值±標準誤差)
2.3 海乳草光合特征分析
不同生境下海乳草的光合特征變化如表3所示。隨著葉面積增大、葉厚度減小,海乳草葉片的凈光合速率、氣孔導度和水分利用效率呈減小趨勢,從樣地I到樣地III分別減小了65.05%、64%和60%,不同生境下海乳草葉片的凈光合速率、氣孔導度和水分利用效率均存在顯著差異(P<0.05);海乳草的蒸騰速率呈減小趨勢,從樣地I到樣地III減小了4.23%,但樣地I,II,III之間無顯著差異(P>0.05)。

表3 不同樣地海乳草光合生理參數特征(數值為平均值±標準誤差)
2.4 海乳草Pn和葉性狀的相關性分析
通過分析發現不同靜水持留條件下海乳草凈光合速率(Pn)與葉面積(LA)、葉厚度(LT)之間均存在顯著差異(P< 0.05)。海乳草葉片凈光合速率和葉面積在樣地(I)和樣地(III)均呈極顯著負相關關系(yⅠ=-0.4135x+0.6037,R2=0.7211,P< 0.01;yⅢ=-1.1882x-0.1609,R2=0.7267,P<0.01,圖1),在樣地(II),二者呈顯著負相關關系(yⅡ=-1.0409x-4E-05,R2=0.4795,P< 0.05,圖1);葉片凈光合速率和葉厚度在樣地(I)和樣地(III)均呈極顯著正相關關系(yⅠ=1.3922x+2.3610,R2=0.7083,P< 0.01;yⅢ=1.2043x+1.8597,R2=0.7419,P<0.01,圖1),在樣地(II),二者呈顯著正相關關系(yⅡ=0.8652x+1.5256,R2=0.5048,P< 0.05,圖1)。表明靜水持留時間較長和較短的生境中海乳草的葉性狀和凈光合速率呈顯著的權衡關系。
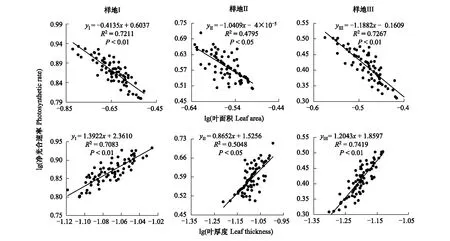
圖1 不同樣地海乳草凈光合速率(Pn)與葉面積(LA)、葉厚度(LT)之間關系Fig.1 Relationship between net photosynthetic rate and leaf area, leaf thickness of Glaux maritima among different plots
2.5 海乳草Tr和葉性狀的相關性分析
Pearson相關性分析表明,不同靜水持留條件下海乳草蒸騰速率(Tr)與葉面積(LA)、葉厚度(LT)之間存在著顯著差異(P<0.05,圖2)。海乳草葉片蒸騰速率和葉面積在樣地(I)和樣地(II)均呈極顯著負相關關系(yⅠ=-0.0469x+0.2982,R2=0.8383,P< 0.01;yⅡ=-1.3338x-0.4362,R2=0.8036,P<0.01,圖2),在樣地(III),二者呈顯著負相關關系(yⅢ=-0.9837x-0.1683,R2=0.5856,P< 0.05,圖2);葉片蒸騰速率和葉厚度在樣地(I)和樣地(II)均呈極顯著正相關關系(yⅠ=0.1628x+0.5028,R2=0.8766,P< 0.01;yⅡ=1.0806x+1.4882,R2=0.8038,P<0.01,圖2),在樣地(III),二者呈顯著正相關關系(yⅢ=1.0064x+1.5157,R2=0.6045,P< 0.05,圖2)。表明隨著靜水持留時間的逐漸增長,海乳草的Tr逐漸減小,葉片葉面積逐漸增大、葉厚度逐漸減小。

圖2 不同樣地海乳草蒸騰速率(Tr)與葉面積(LA)、葉厚度(LT)之間關系Fig.2 Relationship between transpiration rate and leaf area, leaf thickness of Glaux maritima among different plots
3 討論
3.1 不同靜水持留時間下海乳草群落及葉功能性狀的響應變化
植物在受到水鹽脅迫時,能積累滲透調節物質,從而提高滲透勢和調節能力,以適應生存環境[26-27]。通過研究發現,隨著樣地靜水持留時間的變長,濕地群落的平均蓋度、平均密度和土壤含水量逐漸增大,土壤電導率逐漸減小,群落的高度與地上生物量則呈先減小后增大的變化趨勢(表1);海乳草葉片葉面積和比葉面積呈增加趨勢,而葉厚度呈下降趨勢(表2),形成這種變化的主要原因是:(1)樣地I的靜水持留時間最短,地表鹽堿累積時間較長,鹽漬化程度嚴重(表1),耐鹽和泌鹽植物大量生長,加之樣地I土壤含水量最低(表1),因此該生境的密度和蓋度處于較低水平(表1),植株間鄰體干擾和光照資源的競爭不明顯,高鹽脅迫和水分供給成為影響海乳草的主要因素,海乳草為更好適應水鹽脅迫的生境,葉片肉質化,將更多的干物質用于構建保衛組織和葉肉細胞密度[28],并在柵欄組織之間分化出貯水組織應對生理干旱,根據任昱坤[23]的研究可知,貯水組織中含有多糖物質,該物質可以提高葉片的滲透壓和植物對水分的吸收能力,減少葉片水分虧缺,促使植物具有較強的耐旱能力[23];同時在資源有限的條件下,植物為降低陽光對葉片的直射范圍,避免細胞水勢和膨壓的降低[29],以及容納更多的葉綠體,選擇發育了小而厚的葉片以及較低的比葉面積(表2);(2)樣地II的靜水持留時間中等,是鹽漬化程度和土壤含水量的過渡區域(表1),水鹽脅迫比樣地I相對減弱,海乳草成群落優勢種,為滿足植物貯水的需要,維持正常的生理代謝,海乳草選擇了葉面積與葉厚度均衡生長的策略(表2),形成適中的葉綠體和比葉面積(表2),促使植物更好的發展地上生物資源(表1),擴大生存空間,減少生態位重疊,避免了種群內不必要的資源競爭[29];(3)樣地III的靜水持留時間最長,鹽漬化程度較其它樣地最低(表1),耐水淹植物大量發育,植株生長態勢較好,因此群落內植物密度、高度和蓋度最大(表1),植株間鄰體干擾和生存空間、光照資源的競爭激烈,密度制約和鄰體干擾條件成為影響海乳草生長的主要因素。在群落中海乳草鄰體遮陰嚴重,為使葉片盡可能多的接收有限的光資源,提高葉片的光捕捉面積[28],葉片將更多物質用于葉表面積的建設(表2),減少“防御性”的投入,使得葉片柵欄組織細胞層數減少,葉肉細胞排列較疏松,植物單位面積葉片分配的生物量減少,以及葉干重和葉綠素的相應減少,形成了大而薄的葉片以及較大的比葉面積(表2)。表明隨著靜水持留時間的變化,海乳草通過調整自身葉片構型和內在生理生態功能以適應特殊生境,體現了內陸鹽沼濕地植物在嚴酷環境下較強的環境耐受性和抗逆性。
3.2 不同靜水持留時間下海乳草光合性狀的響應變化
葉片光合特性易受環境的影響,植物通過葉片結構性狀和功能性狀的組合和權衡,形成了能夠契合外部環境的形態、生理及物候屬性[30-31]。通過實驗發現,隨著靜水持留時間的增加,葉片的凈光合速率、水分利用效率和氣孔導度均呈下降趨勢,但蒸騰速率差異不明顯(表3)。主要原因是:(1)樣地I海乳草受水鹽脅迫的影響,形成了高度肉質化的厚葉片,能夠容納更多的葉綠體(表2),提高葉肉細胞的光合能力;同時,海乳草葉面積和比葉面積的減小(表2),增加了葉片內部水分向葉片表面擴散的路徑長度,減少了水分的蒸騰散失[1,32],提高了植物的光能利用效率和水分利用效率(表3),促進了植物體內水分和鹽分的運輸[23,33],植物因此獲得了較大的蒸騰速率和凈光合速率(表3),促使其葉厚度與凈光合速率、蒸騰速率均呈極顯著正相關關系(P<0.01,圖1, 2),其葉面積與凈光合速率、蒸騰速率均呈極顯著負相關關系(P<0.01,圖1, 2);(2)樣地II處于樣地I和樣地III過渡區,在資源有限的情況下,海乳草形成適中的葉面積、葉厚度和比葉面積(表2),既能滿足植物貯水的需要,維持正常的生理代謝,又能構建足夠的保護組織防止植物蒸騰失水,實現了葉片對光能利用和截獲能力的最大化,促使凈光合速率和葉面積呈顯著負相關關系,與葉厚度呈顯著正相關關系(P<0.05,圖1);蒸騰速率則和葉面積呈極顯著負相關關系,與葉厚度呈極顯著正相關關系(P<0.01,圖2);(3)樣地III土壤含水量較大,耐水淹濕地植物的大量發育,引起群落中植被間的遮陰加劇,使海乳草種群植株間產生強烈的光競爭,葉片葉綠素含量大幅度減少(表2),葉肉細胞光合能力受到抑制;同時大而薄的葉片能夠使CO2和水分等從氣孔到達葉綠體的路徑縮短,有利于光的透過和光能的吸收[34],相較于高鹽環境該生境的比葉面積較大(表2),植物生長速率高,養分元素保有能力降低,較小的葉厚度造成了葉脈中維管束極不發達和貯水組織的逐漸退化,促使植物葉水勢的降低,鹽分輸送效率的減小[23],氣孔的部分關閉以及該群落嚴重的遮陰降低了葉片的蒸騰速率,造成了葉片水分利用效率較低(表3),植物因此獲得了較低的凈光合速率,促使其凈光合速率分別與葉面積、葉厚度呈極顯著負相關和極顯著正相關關系(P<0.01,圖1);蒸騰速率則分別與葉面積、葉厚度呈顯著負相關和顯著正相關關系(P<0.05,圖2)。因此,在各樣地之間凈光合速率、蒸騰速率與葉面積、葉厚度存在著顯著差異(P<0.05,圖1, 2),表明隨著生境的改變,海乳草葉片積極調整各構件的光合碳同化產物權衡分配機制,體現了植物葉片在不同生境下為規避環境壓力所選擇的生態策略。
4 結論
植物葉片形態和光合生理特征在不同生境之間差異性,顯示了植物在外在形態和內在生理方面的協同變異性,這是植物長期適應環境的結果。本研究發現:當海乳草面臨水鹽雙重脅迫的生境時,植株選擇減小葉面積增大葉厚度的生長策略,提高凈光合速率,以實現植物自身碳同化產物權衡分配;在水分供給充足的生境,海乳草葉片選擇增大葉面積減小葉厚度,提高了葉面積的資源分配和光合效應,實現植物正常的生長繁殖。本文僅從靜水持留時間影響下葉性狀與光合特性的變化規律進行了分析,葉綠素熒光、光譜等也與葉性狀變化有密切關系,對這些方面的研究是今后關注的重點。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
