不同捕食者暴露對(duì)鯽幼魚行為、運(yùn)動(dòng)和形態(tài)特征的影響
2021-07-09 05:07:08肖玲韜趙昕雨黃恩梅徐佳佳付世建
生態(tài)學(xué)報(bào) 2021年11期
關(guān)鍵詞:實(shí)驗(yàn)
付 成,肖玲韜,趙昕雨,黃恩梅,徐佳佳,王 亞,付世建
重慶師范大學(xué) 進(jìn)化生理與行為學(xué)實(shí)驗(yàn)室 重慶市動(dòng)物生物學(xué)重點(diǎn)實(shí)驗(yàn)室,重慶 401331
捕食作為一種重要的自然選擇壓力,可能導(dǎo)致獵物在行為和生理功能等方面產(chǎn)生表型和(或)遺傳層次的改變,因此一直備受關(guān)注[1- 3]。已有研究發(fā)現(xiàn),相比直接致命性的捕食,捕食者對(duì)獵物所產(chǎn)生的風(fēng)險(xiǎn)效應(yīng)通常對(duì)獵物種群的影響更為顯著[1]。獵物應(yīng)對(duì)被捕食風(fēng)險(xiǎn)所做出的行為以及生理功能等方面的調(diào)整可以稱之為獵物的“反捕食響應(yīng)”,反捕食響應(yīng)通常可以有效提高獵物在捕食脅迫下的生存機(jī)率[4-5]。
就魚類而言,其反捕食響應(yīng)多種多樣,例如在被捕食風(fēng)險(xiǎn)較高時(shí),有些魚類會(huì)降低自發(fā)運(yùn)動(dòng)[6]、減少覓食行為[7]和冒險(xiǎn)行為[7],降低勇敢性[8]以及提高隱蔽場(chǎng)所利用率[9],因?yàn)檫@些行為的調(diào)整都有利于降低獵物魚被捕食者發(fā)現(xiàn)的機(jī)率;在運(yùn)動(dòng)能力方面,研究者發(fā)現(xiàn)無論是自然生境中長(zhǎng)期的捕食壓力[10],還是實(shí)驗(yàn)室內(nèi)短期的捕食脅迫處理[5]均可提高獵物魚的快速啟動(dòng)逃逸能力,較強(qiáng)的快速啟動(dòng)逃逸能力可以保證獵物魚在被捕食者發(fā)現(xiàn)或襲擊時(shí)能順利逃脫[11-12]。此外,在形態(tài)上部分魚類會(huì)通過增加體高的方式對(duì)捕食者形成口裂限制[13],體高的提高也會(huì)間接導(dǎo)致魚類快速啟動(dòng)逃逸能力的提升[11],因此對(duì)于魚類逃避捕食者十分關(guān)鍵。盡管關(guān)于魚類反捕食響應(yīng)的研究較多,但大多數(shù)研究?jī)H集中于某一種反捕食方式,該領(lǐng)域仍缺乏多種反捕食策略的整合性研究,使得我們對(duì)魚類反捕食響應(yīng)的理解依舊有限。因?yàn)轸~類應(yīng)對(duì)捕食脅迫可能是利用多種反捕食策略的綜合,以此獲得最大的反捕食效果;也可能是當(dāng)某種或數(shù)種反捕食策略足以確保安全時(shí),放棄其他方式的反捕食策略,因?yàn)榇蠖鄶?shù)的反捕食響應(yīng)都是有代價(jià)的。例如提高隱蔽場(chǎng)所利用率雖然可以確保安全,但也同時(shí)使獵物喪失了獲取食物、配偶的機(jī)會(huì)[14]。
自然水體中魚類的捕食者種類繁多,這些捕食者從形態(tài)特征(如口裂的大小)到捕食方式(如追擊或伏擊取食),再到覓食頻率等均存在著較大的差異[15- 17]。不同類型的捕食者(或者不同類型的捕食者同時(shí)出現(xiàn))給獵物魚所帶來捕食脅迫的程度及其影響方式可能是截然不同的,那么獵物魚針對(duì)這些不同的捕食脅迫是否會(huì)做出相異的策略選擇呢?組合多種反捕食策略以最大化生存,還是僅維持少數(shù)的反捕食策略以保證生長(zhǎng)呢?
鯽(Carassiusauratus)屬鯉科魚類,身體呈梭形,體高較高,適應(yīng)能力強(qiáng),在我國(guó)分布范圍極為廣泛,以往研究發(fā)現(xiàn)其對(duì)環(huán)境變化具備較高的生理可塑性[18-19]。有研究認(rèn)為,其能夠如此廣泛的分布于我國(guó)各大水系,可能與其較強(qiáng)的反捕食能力有一定關(guān)聯(lián)[20]。因此本研究選取鯽幼魚作為獵物魚。烏鱧(Channaargus)和胡子鯰(Clariasfuscus)這兩種魚類捕食者與鯽的棲息地有較多重疊,為鯽幼魚在自然界中常見的捕食者[21-22]。這兩種捕食者在形態(tài)特征及捕食習(xí)性等方面存在較大差異:烏鱧為鱸形目,口裂很大,常靜待獵物靠近時(shí)突然發(fā)動(dòng)襲擊[15];胡子鯰為鲇形目,口裂較小,覓食行為活躍,頻繁尾隨和追擊獵物,待接近獵物時(shí)發(fā)動(dòng)襲擊[21, 23]。本研究選擇鯽幼魚為獵物魚,烏鱧和胡子鯰為捕食者,將鯽幼魚分別暴露于空白(對(duì)照)、烏鱧、胡子鯰和雙捕食者(同時(shí)存在烏鱧和胡子鯰)環(huán)境中持續(xù)2個(gè)月,隨后通過考查和對(duì)比不同組鯽幼魚的生長(zhǎng)、行為特征、形態(tài)和運(yùn)動(dòng)能力等指標(biāo)來探討鯽幼魚是否會(huì)針對(duì)不同的捕食者做出截然不同的反捕食策略選擇。
1 材料與方法
1.1 實(shí)驗(yàn)魚來源與馴養(yǎng)
實(shí)驗(yàn)用鯽幼魚、烏鱧和胡子鯰購(gòu)置于當(dāng)?shù)貪O場(chǎng),分別放入本實(shí)驗(yàn)室規(guī)格為2m×1m×0.5m的水槽內(nèi)馴養(yǎng)一個(gè)月。馴養(yǎng)期間,每天上午9:00,鯽幼魚以商業(yè)顆粒飼料飽足投喂一次,烏鱧和胡子鯰每隔兩天投喂一次鰱魚肉片搭配商業(yè)餌料至飽足。所有實(shí)驗(yàn)魚均在每次投喂30min后使用虹吸管將未被吃完的食物以及糞便清理干凈。實(shí)驗(yàn)用水為曝氣除氯后的自來水,日換水量約為水槽總水體體積的20%,馴化溫度為(20.0±1)℃。用充氣泵不斷向水體充入空氣使其溶氧量接近飽和,自然光周期(正式實(shí)驗(yàn)于8—11月完成)。
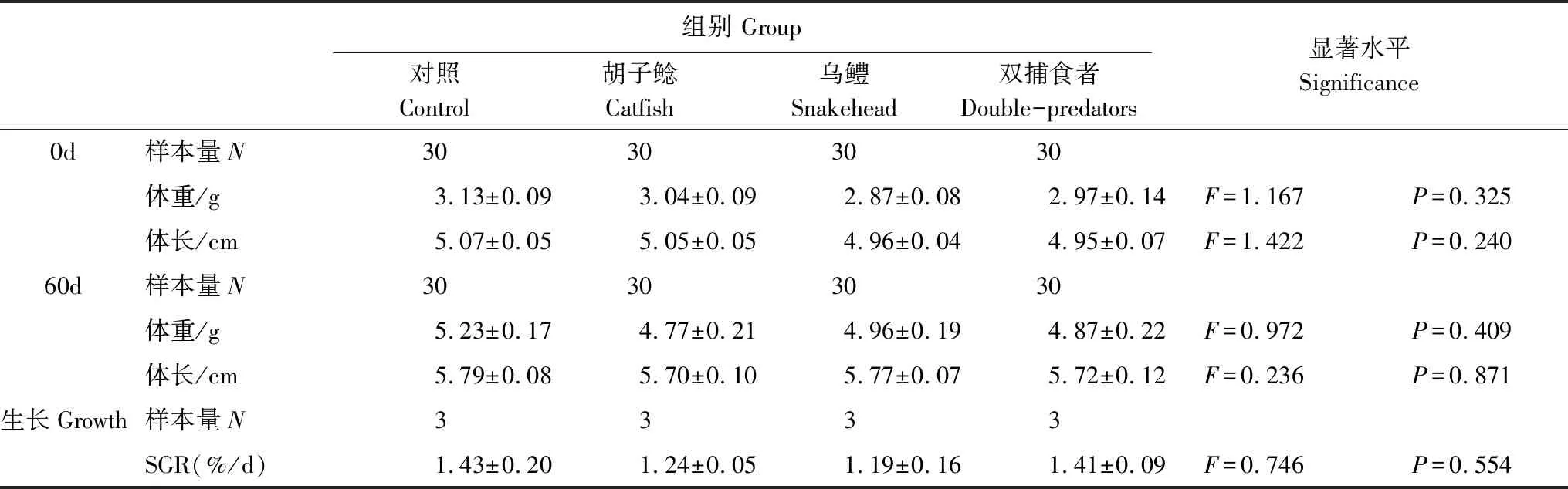
表1 不同捕食者暴露對(duì)鯽幼魚生長(zhǎng)的影響 (Mean±SE)
1.2 實(shí)驗(yàn)設(shè)計(jì)
馴養(yǎng)結(jié)束后,挑選身體健康、大小接近的鯽幼魚隨機(jī)分成四組(表1),即一個(gè)對(duì)照組和胡子鯰暴露組、烏鱧暴露組、雙捕食者(烏鱧和胡子鯰同時(shí)存在)暴露組三個(gè)捕食者處理組,每組30尾,分別放入規(guī)格相同的水槽(圖1;1m×0.6m×0.5m)中飼養(yǎng),每個(gè)水槽10尾鯽幼魚。水槽內(nèi)水深約30cm,光照強(qiáng)度約300lx。挑選大小接近的胡子鲇(200—300g;口裂寬1.5—2.0cm;口裂深0.5—1cm)和烏鱧(200—300g;口裂寬3—3.8cm;口裂深2.8—3.7cm)作為捕食者,分別加入對(duì)應(yīng)的捕食處理組的水槽之中(N=2,雙捕食者暴露組中為1尾胡子鯰和1尾烏鱧),但將捕食者與鯽幼魚用鐵絲網(wǎng)分隔開,并在捕食者的一側(cè)(鐵絲網(wǎng)外側(cè))投放數(shù)尾不用于實(shí)驗(yàn)測(cè)定的鯽幼魚作為餌料魚,使得鐵絲網(wǎng)內(nèi)側(cè)的鯽幼魚雖然不會(huì)被捕食,但可以通過視覺和嗅覺的方式感受捕食信息。對(duì)照組鐵絲網(wǎng)外側(cè)不放置捕食者,其它環(huán)境條件與處理組完全相同。另外每個(gè)水槽中鯽幼魚一側(cè)均放置有一不透明的PVC管(長(zhǎng)20cm,直徑10cm),用以給鯽幼魚提供隱蔽場(chǎng)所。處理期間其他養(yǎng)殖條件與馴養(yǎng)期間保持一致,連續(xù)處理60d后,測(cè)定鯽幼魚的體重、體長(zhǎng)(表1)。隨后挑選身體健康、大小接近的鯽幼魚分別進(jìn)行行為、快速啟動(dòng)游泳能力、臨界游泳速度和形態(tài)的測(cè)定(表2)。

表2 不同實(shí)驗(yàn)測(cè)定(行為、快速啟動(dòng)游泳能力、臨界游泳速度和形態(tài))中所使用鯽幼魚的樣本量、體重和體長(zhǎng) (Mean±SE)
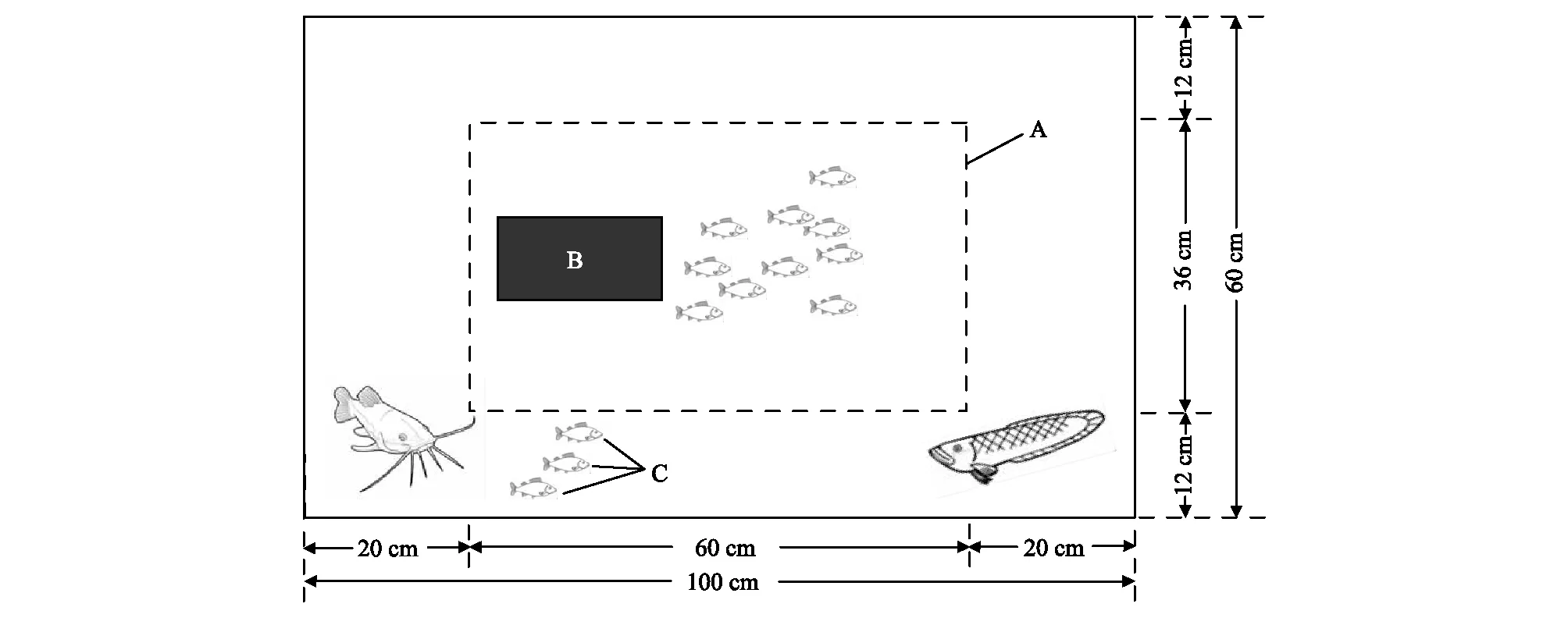
圖1 捕食者暴露處理水槽示意圖(俯視)Fig.1 A experimental tank was used for predator exposure (top view)圖中A為鐵絲網(wǎng);B為隱蔽場(chǎng)所(不透明PVC管;長(zhǎng)度20 cm,直徑10 cm);C為不用于實(shí)驗(yàn)測(cè)定的餌料魚;根據(jù)組別的不同,鐵絲網(wǎng)外側(cè)分別飼養(yǎng)不同的捕食者;對(duì)照組:無捕食者;胡子鯰暴露組:兩尾胡子鲇;烏鱧暴露組:兩尾烏鱧;雙捕食者暴露組:一尾胡子鲇和一尾烏鱧
1.3 實(shí)驗(yàn)參數(shù)的測(cè)定
1.3.1生長(zhǎng)能力的測(cè)定
鯽幼魚分組完成后開始生長(zhǎng)實(shí)驗(yàn)。每天上午9:00,鯽幼魚以商業(yè)顆粒飼料飽足投喂一次,烏鱧和胡子鯰每隔兩天投喂一次鰱魚肉片搭配商業(yè)餌料至飽足。分別在第0d和第60d測(cè)定4組鯽幼魚的體重、體長(zhǎng)用以計(jì)算生長(zhǎng)能力(表1)。稱重前,實(shí)驗(yàn)魚禁食24h。生長(zhǎng)能力使用下列公式進(jìn)行計(jì)算:特殊生長(zhǎng)率(special growth rate, SGR; %/d)=(lnM2-lnM1)/T×100。式中M1和M2分別為第0d和第60d每個(gè)水槽中10尾實(shí)驗(yàn)魚體重的平均值(g);T為生長(zhǎng)時(shí)間60d。
1.3.2行為特征的測(cè)定
鯽幼魚禁食24h后,首先進(jìn)行自發(fā)運(yùn)動(dòng)的測(cè)定。將單尾實(shí)驗(yàn)魚轉(zhuǎn)移至圓形的行為觀察裝置[14](120 L)中馴化1h。觀察裝置水深12 cm,用以降低實(shí)驗(yàn)魚的垂直移動(dòng)[24],裝置正上方安裝有攝像頭(羅技,Pro 9000),用于實(shí)驗(yàn)魚的行為拍攝。1h后對(duì)實(shí)驗(yàn)魚進(jìn)行20 min的自發(fā)運(yùn)動(dòng)拍攝。所有的視頻均使用行為學(xué)軟件EthoVision(EthoVision XT 9,荷蘭Nodus公司)進(jìn)行分析。分析過程中,使用“zones”工具將圓形的觀察水面(直徑38 cm)劃分為兩個(gè)不同的區(qū)域:中間的圓形區(qū)域(直徑26 cm)和周圍的環(huán)形區(qū)域(寬度6 cm,約1倍實(shí)驗(yàn)魚體長(zhǎng))。中間的圓形區(qū)域作為開放水體定義為風(fēng)險(xiǎn)區(qū)域,周圍的環(huán)形區(qū)域靠近觀察裝置的側(cè)壁,可能會(huì)給實(shí)驗(yàn)魚提供隱蔽[9],因此本研究中將其定義為安全區(qū)域[14]。自發(fā)運(yùn)動(dòng)評(píng)價(jià)指標(biāo)包括:實(shí)驗(yàn)魚在風(fēng)險(xiǎn)區(qū)域停留時(shí)間百分比(%),風(fēng)險(xiǎn)區(qū)域停留頻次百分比(%),運(yùn)動(dòng)時(shí)間比(實(shí)驗(yàn)魚處于運(yùn)動(dòng)狀態(tài)的累計(jì)時(shí)間占總時(shí)間的百分比,%)和平均游泳速度(cm/s)。
自發(fā)運(yùn)動(dòng)測(cè)定完成后,在觀察裝置中放入一根PVC管(與馴化期間規(guī)格相同)作為隱蔽場(chǎng)所。2min后對(duì)實(shí)驗(yàn)魚進(jìn)行20 min的隱匿行為拍攝。所有的視頻均使用行為學(xué)軟件EthoVision進(jìn)行分析。實(shí)驗(yàn)魚在隱蔽場(chǎng)所中停留的累計(jì)時(shí)間百分比即為隱蔽場(chǎng)所利用率(%)。
鯽幼魚的隱匿行為測(cè)定完成后,將單尾實(shí)驗(yàn)魚轉(zhuǎn)移至勇敢性測(cè)定裝置[14]中進(jìn)行勇敢性的測(cè)定。測(cè)定裝置(60 cm × 20 cm × 20 cm)由透明的有機(jī)玻璃構(gòu)成。四周貼有不透明的白紙用以降低外界對(duì)實(shí)驗(yàn)魚的干擾。由一塊帶有小門的白板將整個(gè)區(qū)域分為隱蔽場(chǎng)所(上方有蓋板,可提供隱蔽;20 cm × 20 cm × 20 cm)和開放水域(40 cm × 20 cm × 20 cm),小門為實(shí)驗(yàn)魚進(jìn)出隱蔽場(chǎng)所的通道。水深保持在12 cm。將單尾實(shí)驗(yàn)魚轉(zhuǎn)移置至開放區(qū)域中適應(yīng)1h,1h后輕輕拉開小門,隨后將一個(gè)小型抄網(wǎng)快速下墜至開放水域中(模擬一次捕食者襲擊),將實(shí)驗(yàn)魚趕入隱蔽場(chǎng)所之中,隨后使用連接電腦的攝像頭紀(jì)錄實(shí)驗(yàn)魚首次從隱蔽場(chǎng)所游出的時(shí)間[20, 25]。實(shí)驗(yàn)魚遭遇一次捕食者襲擊(模擬)后首次從安全的隱蔽場(chǎng)所游出進(jìn)入開放環(huán)境的時(shí)間即勇敢性(s)[25]。
1.3.3快速啟動(dòng)游泳能力的測(cè)定
實(shí)驗(yàn)魚禁食24h后,采用實(shí)驗(yàn)室自行研制的快速啟動(dòng)測(cè)定系統(tǒng)對(duì)實(shí)驗(yàn)魚進(jìn)行快速啟動(dòng)游泳能力的測(cè)定[26]。將一小顆白色的泡沫球(直徑約1mm,質(zhì)量< 0.001g)用少量膠粘附在實(shí)驗(yàn)魚背部的質(zhì)心位置。隨后將實(shí)驗(yàn)魚轉(zhuǎn)移至快速啟動(dòng)測(cè)定水槽(40 × 40 × 15 cm)中進(jìn)行1h的恢復(fù)和適應(yīng)。測(cè)定水槽水深10cm,水槽底部為刻有方形網(wǎng)格的白色有機(jī)玻璃板,用于后續(xù)圖像分析過程中的距離校正。水槽四周不透明,用以減少外界對(duì)實(shí)驗(yàn)魚的干擾。適應(yīng)結(jié)束后待實(shí)驗(yàn)魚游至水槽中央時(shí),打開高速相機(jī)(A504K,德國(guó)Basler公司;500幀/s),同時(shí)給予實(shí)驗(yàn)魚一次電刺激(電場(chǎng)強(qiáng)度0.55 V/cm,持續(xù)時(shí)間50 ms)[27],電刺激裝置連接有發(fā)光二極管作為指示燈。高速相機(jī)紀(jì)錄下電刺激引起的實(shí)驗(yàn)魚逃逸反應(yīng)的全過程,拍攝的序列照片隨后使用TpsUtil和TpsDig軟件來分析整個(gè)逃逸過程中實(shí)驗(yàn)魚質(zhì)心的移動(dòng)軌跡。實(shí)驗(yàn)魚質(zhì)心的原始軌跡進(jìn)行5點(diǎn)3次平滑化處理[2]。后續(xù)快速啟動(dòng)參數(shù)計(jì)算:最大線速度(Vmax, m/s), 最大線加速度(Amax,m/s2),120ms內(nèi)的移動(dòng)距離(S120 ms,cm)和反應(yīng)時(shí)滯(T,ms)。其中Vmax和Amax分別為實(shí)驗(yàn)魚逃逸過程中質(zhì)心移動(dòng)的最大線性速度和加速度。S120 ms為實(shí)驗(yàn)魚逃逸過程中電刺激開始后前120ms實(shí)驗(yàn)魚質(zhì)心的移動(dòng)總路程。參數(shù)T是實(shí)驗(yàn)魚從電刺激開始(LED燈亮)到逃逸反應(yīng)開始的時(shí)間間隔[28]。
雪螢一怔:“江老師這不是帶給你了嗎?”狗子說:“他今天才給我的。以前那么久,他都沒給我。你一定是才買的。”雪螢哭笑不得,說:“是是是,阿姨太忙,忘了,對(duì)不起。”
1.3.4臨界游泳速度和代謝率的測(cè)定
將實(shí)驗(yàn)魚禁食24h后轉(zhuǎn)移至實(shí)驗(yàn)室自行研制的Blazka式游泳代謝測(cè)定儀[29]中進(jìn)行臨界游泳速度Ucrit和代謝率的測(cè)定,儀器的原理結(jié)構(gòu)詳見已發(fā)表論文[26]。實(shí)驗(yàn)魚轉(zhuǎn)移至游泳管之后進(jìn)行2 h 的適應(yīng),適應(yīng)期間游泳管密封蓋打開用于換水,水流速度為6 cm/s。適應(yīng)結(jié)束后,以6 cm/s作為起始流速,每隔20 min 水流速度增加6 cm/s,直至實(shí)驗(yàn)魚力竭。隨后取出實(shí)驗(yàn)魚測(cè)量體重、體長(zhǎng)。實(shí)驗(yàn)魚力竭的判斷標(biāo)準(zhǔn)為實(shí)驗(yàn)魚不能在游泳管中保持位置而被沖到篩板上持續(xù)20 s[30]。Ucrit的計(jì)算使用以下公式[31]:
Ucrit=V+(t/T)ΔV
其中V是實(shí)驗(yàn)魚完整游完的最大速度(力竭前一檔速度,cm/s);ΔV為每一檔的水流速度增量(6 cm/s);T是每一檔的設(shè)定游泳時(shí)間(20 min);t是實(shí)驗(yàn)魚在最大速度檔(最后一檔)的實(shí)際游泳時(shí)間(min)。
游泳管分開放和封閉兩種模式,開放模式用于換水,封閉模式用于測(cè)定實(shí)驗(yàn)魚的代謝率。封閉模式狀態(tài)下,每2 min紀(jì)錄一次管中溶氧水平,用于計(jì)算實(shí)驗(yàn)魚的代謝率,計(jì)算公式如下[32]:
Mo2=60(St-S0)v/m
其中Mo2為實(shí)驗(yàn)魚的運(yùn)動(dòng)代謝率(mgO2kg-1h-1);St和S0(斜率,mgO2L-1min-1)分別為游泳管內(nèi)有魚和無魚時(shí)溶氧水平的下降率(用以扣除細(xì)菌耗氧),通過氧含量(mg/L)與時(shí)間(min)的線性回歸計(jì)算而來;v是游泳管的總體積(3L);m為實(shí)驗(yàn)魚的體重(kg)。測(cè)定過程中的最大Mo2定義為活躍代謝率MO2active,作為實(shí)驗(yàn)魚代謝能力的指標(biāo)。
1.3.5形態(tài)的測(cè)定
從每組中分別選取13尾大小接近的實(shí)驗(yàn)魚,使用過量MS- 222麻醉致死,隨后測(cè)量相關(guān)形態(tài)學(xué)參數(shù)(表3)。測(cè)定時(shí)將實(shí)驗(yàn)魚的背鰭、尾鰭、臀鰭等依次展開后固定在放有刻度尺的泡沫板上,采用數(shù)碼相機(jī)拍攝左側(cè)照片[33]。參照閆冠杰等人[33-34]的形態(tài)學(xué)分析方法,使用TpsDig2軟件對(duì)實(shí)驗(yàn)魚照片進(jìn)行處理,得到包含16個(gè)特征點(diǎn)的圖像。通過上述每尾實(shí)驗(yàn)魚16個(gè)特征點(diǎn)的坐標(biāo)計(jì)算相關(guān)形態(tài)學(xué)參數(shù),包括叉長(zhǎng)、腹鰭前長(zhǎng)、頭長(zhǎng)、頭高、體高、尾柄高1和尾柄高2。另外通過TpsDig 2軟件計(jì)算尾柄側(cè)面積和尾鰭表面積。上述參數(shù)的單位均通過圖像距離與實(shí)際距離(從照片中的刻度尺獲得)之間的轉(zhuǎn)換關(guān)系換算為mm或mm2。

表3 不同捕食者暴露對(duì)鯽幼魚形態(tài)特征的影響
1.4 數(shù)據(jù)統(tǒng)計(jì)與分析
實(shí)驗(yàn)數(shù)據(jù)采用Excel 2016進(jìn)行常規(guī)計(jì)算,使用統(tǒng)計(jì)學(xué)軟件SPSS 17.0進(jìn)行統(tǒng)計(jì)分析。不同組之間生長(zhǎng)、行為、游泳能力和形態(tài)參數(shù)之間的差異使用單因素方差分析。所有數(shù)據(jù)均以平均值±標(biāo)準(zhǔn)誤(Mean±SE)表示,顯著性水平為P<0.05。
2 結(jié)果
2.1 不同捕食者暴露對(duì)鯽幼魚生長(zhǎng)的影響
經(jīng)過60d的生長(zhǎng),相比初始值,所有組實(shí)驗(yàn)魚的體重、體長(zhǎng)均顯著提高(P<0.05;表1)。然而無論是在捕食暴露前、后,相比對(duì)照組,三個(gè)捕食者暴露組的鯽幼魚的體重、體長(zhǎng)均未表現(xiàn)出顯著差異(表1)。此外,不同組之間生長(zhǎng)能力SGR也未表現(xiàn)出顯著差異(F=0.746,P=0.554;表1)。
2.2 不同捕食者暴露對(duì)鯽幼魚形態(tài)的影響
經(jīng)過60d的捕食者暴露,烏鱧暴露組的鯽幼魚體高(BD)顯著高于對(duì)照組(P<0.001)和鯰魚暴露組(P<0.001),但與雙捕食者暴露組無顯著差異(P=0.561;表3)。其他所有形態(tài)學(xué)參數(shù)在捕食暴露組與對(duì)照組之間均未表現(xiàn)出顯著性差異(表3)。
2.3 不同捕食者暴露對(duì)鯽幼魚行為特征的影響
經(jīng)過60d的捕食者暴露,鯽幼魚自發(fā)運(yùn)動(dòng)的四個(gè)指標(biāo),風(fēng)險(xiǎn)區(qū)域停留時(shí)間百分比(F=0.512,P=0.676)、風(fēng)險(xiǎn)區(qū)域停留頻次百分比(F=0.747,P=0.529)、運(yùn)動(dòng)時(shí)間比(F=1.119,P=0.348)和平均速度(F=1.034,P=0.384)在四個(gè)組之間均未表現(xiàn)出顯著性差異(圖2)。
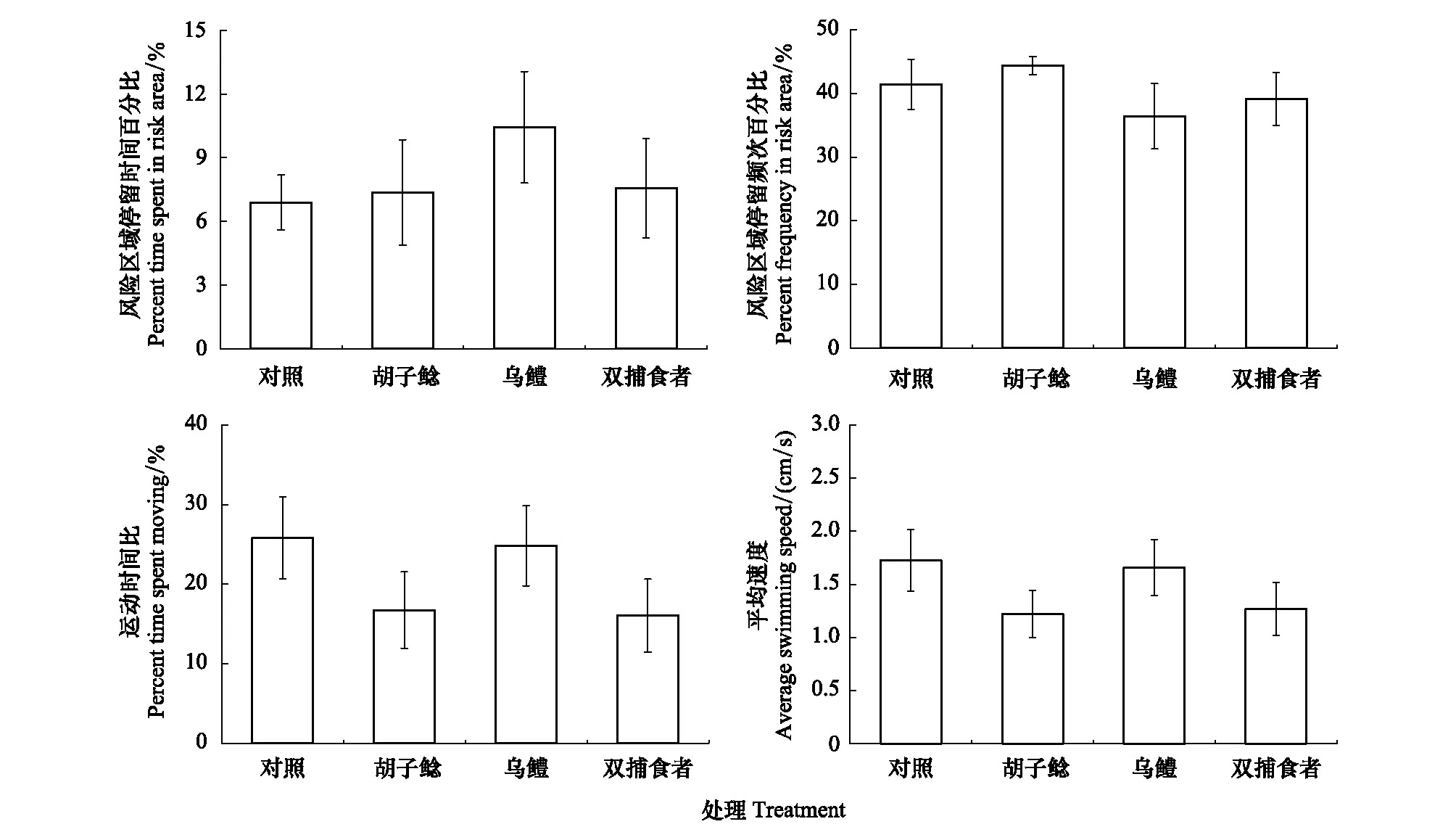
圖2 不同捕食者暴露對(duì)鯽幼魚自發(fā)運(yùn)動(dòng)的影響Fig.2 The effects of different predator exposure on spontaneous activity in juvenile crucian carp
此外,鯽幼魚的隱蔽場(chǎng)所利用率(F=1.083,P=0.363)以及勇敢性(F=0.851,P=0.473)在四個(gè)組之間同樣未表現(xiàn)出顯著性差異(圖3)。
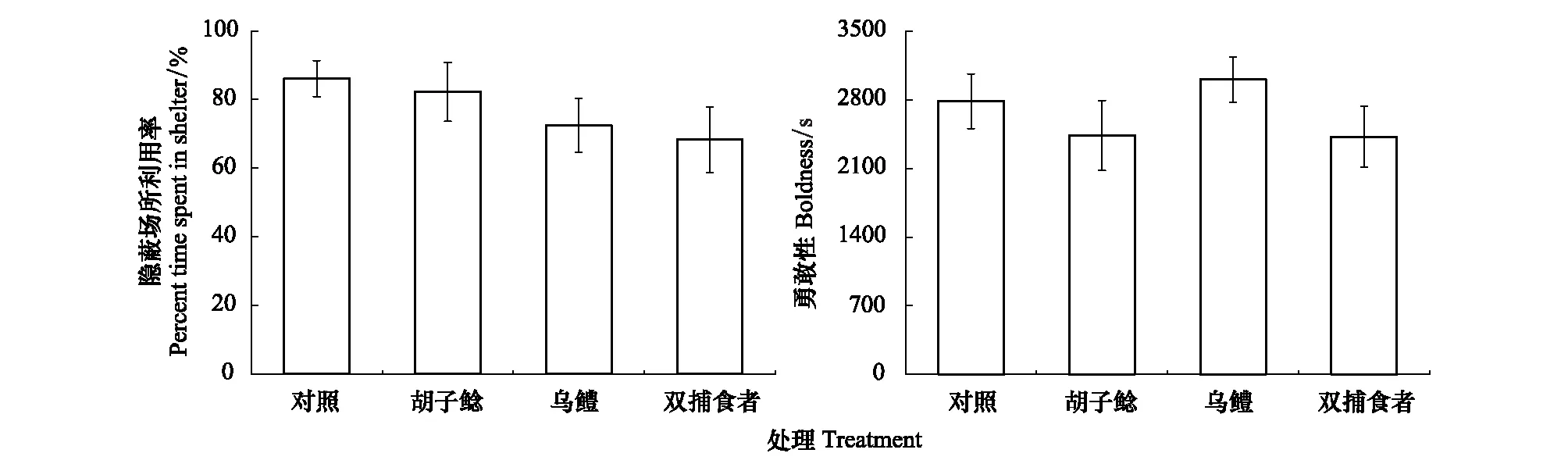
圖3 不同捕食者暴露對(duì)鯽幼魚隱蔽場(chǎng)所利用率和勇敢性的影響Fig.3 The effects of different predator exposure on percent time spent in shelter and boldness in juvenile crucian carp
2.4 不同捕食者暴露對(duì)鯽幼魚快速啟動(dòng)游泳能力的影響
經(jīng)過捕食者暴露,鯽幼魚的快速啟動(dòng)最大速度(F=1.415,P=0.253)和最大加速度(F=0.647,P=0.590)在四個(gè)組之間未表現(xiàn)出顯著差異(圖4);而快速啟動(dòng)120ms移動(dòng)距離烏鱧暴露組顯著高于對(duì)照組(P=0.016;圖4);另外反應(yīng)時(shí)滯胡子鯰暴露組(P=0.047)、烏鱧暴露組(P=0.035)和雙捕食者暴露組(P=0.023)均顯著低于對(duì)照組(圖4)。
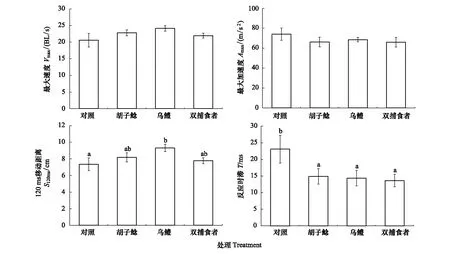
圖4 不同捕食者暴露對(duì)鯽幼魚快速啟動(dòng)游泳能力的影響Fig.4 The effects of different predator exposure on fast-start swimming performance in juvenile crucian carp
2.5 不同捕食者暴露對(duì)鯽幼魚臨界游泳速度的影響
經(jīng)過60d的捕食者暴露,鯽幼魚的臨界游泳速度(F=0.280,P=0.840)和活躍代謝率(F=1.269,P=0.302)在四個(gè)組之間均未表現(xiàn)出顯著差異(圖5)。
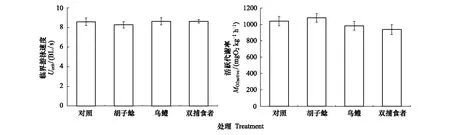
圖5 不同捕食者暴露對(duì)鯽幼魚臨界游泳速度和活躍代謝率的影響Fig.5 The effects of different predator exposure on critical swimming speed and active metabolic rate in juvenile crucian carp
3 討論
3.1 不同捕食者暴露對(duì)鯽幼魚行為特征的影響
魚類的行為特征中自發(fā)運(yùn)動(dòng)、隱蔽場(chǎng)所利用率以及勇敢性等均與其逃避捕食者密切相關(guān)[14]。魚類的自發(fā)運(yùn)動(dòng)越低,其在水中留下的化學(xué)以及水動(dòng)力學(xué)的信號(hào)就越少,那么其被捕食者發(fā)現(xiàn)的概率就越低[35]。此外,較低的自發(fā)運(yùn)動(dòng)也能降低獵物魚與捕食者直接相遇的幾率。因此多數(shù)研究者認(rèn)為,在有捕食脅迫的情況下魚類可能會(huì)降低其自發(fā)運(yùn)動(dòng)[7, 36-37]。另外增加隱蔽場(chǎng)所的使用,以及降低勇敢性同樣使得獵物魚更難被捕食者發(fā)現(xiàn),而隱蔽場(chǎng)所的使用使得獵物魚即使被捕食者發(fā)現(xiàn),仍能藏身于隱蔽場(chǎng)所內(nèi)拒止大型的捕食者[9, 37]。
然而,在本研究中,鯽幼魚自發(fā)運(yùn)動(dòng)的4個(gè)指標(biāo)在不同組之間均未表現(xiàn)出顯著性差異。另外,不同組鯽幼魚的隱蔽場(chǎng)所利用率以及勇敢性也未表現(xiàn)出顯著性差異。這可能像本研究之前預(yù)期的那樣,當(dāng)一種或幾種反捕食策略足夠應(yīng)對(duì)捕食脅迫時(shí),其他的反捕食策略可能會(huì)趨于保守,因?yàn)榻^大多數(shù)的反捕食策略都是有代價(jià)的[38-39]。相比運(yùn)動(dòng)能力與形態(tài),魚類反捕食行為的代價(jià)可能更為嚴(yán)重:降低自發(fā)運(yùn)動(dòng)、提高隱蔽場(chǎng)所利用率以及降低勇敢性均會(huì)使其獲取食物、占有領(lǐng)地的幾率大大降低[14],這對(duì)其長(zhǎng)期生存是非常不利的。此外,在本研究中,捕食者與獵物魚是通過鐵絲網(wǎng)隔開的,捕食者對(duì)鯽幼魚而言并非致命性的威脅,可能正是因?yàn)檫@個(gè)原因,鯽幼魚的反捕食行為未出現(xiàn)顯著變化。對(duì)應(yīng)的,經(jīng)過2個(gè)月的處理,不同組之間鯽幼魚的體重、體長(zhǎng)和SGR均未表現(xiàn)出顯著性差異也進(jìn)一步佐證了這一點(diǎn)。
3.2 不同捕食者暴露對(duì)鯽幼魚形態(tài)和運(yùn)動(dòng)能力的影響
魚類的形態(tài)特征與運(yùn)動(dòng)能力密切關(guān)聯(lián)[12, 40]。例如較高的身體高度,較大的尾鰭面積有利于魚類在運(yùn)動(dòng)過程中產(chǎn)生更大的推進(jìn)力,進(jìn)而提高快速啟動(dòng)逃逸能力[2, 41]。本研究中,經(jīng)過2個(gè)月的處理,與對(duì)照組相比,僅烏鱧暴露組的鯽幼魚體高表現(xiàn)出顯著性提高。在運(yùn)動(dòng)能力方面,與對(duì)照組相比,鯽幼魚快速啟動(dòng)參數(shù)中S120ms僅烏鱧暴露組顯著提高,這與形態(tài)學(xué)的變化基本上是一致的,進(jìn)一步證明了魚類形態(tài)與運(yùn)動(dòng)能力之間的緊密關(guān)系:提高的體高有利于提高魚類的快速啟動(dòng)逃逸能力。此外提高的體高也使獵物魚更容易對(duì)捕食者形成口裂限制[42],這些運(yùn)動(dòng)能力與形態(tài)特征的變化均有利于獵物魚提高生存幾率[43]。另外,相比對(duì)照組,胡子鯰暴露組、烏鱧暴露組和雙捕食者暴露組實(shí)驗(yàn)魚的快速啟動(dòng)T均顯著縮短。這種在捕食脅迫下縮短快速啟動(dòng)T的策略在近期研究中已有發(fā)現(xiàn)[26, 44-45],本研究進(jìn)一步證實(shí)了T對(duì)捕食脅迫的高敏感性。盡管這種策略對(duì)于獵物逃避捕食非常關(guān)鍵[26, 44],但T在捕食脅迫下縮短的內(nèi)在機(jī)理仍有待于進(jìn)一步研究。
本研究對(duì)鯽幼魚的Ucrit和MO2active也進(jìn)行了測(cè)定,旨在驗(yàn)證魚類的臨界游泳能力對(duì)于其逃避捕食者是否同樣較為關(guān)鍵[45],尤其是在應(yīng)對(duì)捕食者連續(xù)尾隨和追擊的情況下是否更為重要。然而本研究發(fā)現(xiàn)相比對(duì)照組,三個(gè)捕食者暴露組的Ucrit和MO2active均未表現(xiàn)出顯著差異。這也表明:至少對(duì)于胡子鯰和烏鱧,在應(yīng)對(duì)這兩種捕食者脅迫時(shí)快速啟動(dòng)游泳能力依然扮演著更為重要的角色。
3.3 不同捕食者暴露與多捕食者效應(yīng)對(duì)鯽幼魚的整體影響
在自然界中,動(dòng)物的捕食者多種多樣,因此動(dòng)物常常需要分別或同時(shí)應(yīng)對(duì)不同捕食者所帶來的風(fēng)險(xiǎn)[46-47]。對(duì)此,獵物魚所做出的反捕食響應(yīng)可能是截然不同的[48]。這一觀點(diǎn)在蝌蚪(Ranatemporaria)[49]、蜻蜓(Pachydiplaxlongipennis)幼蟲[50]、沙鼠(Gerbillusandersoni)[51]和淡水蝸牛(Physagyrina;Physaheterostropha)[46]等動(dòng)物中已得到驗(yàn)證,但魚類的相關(guān)研究非常有限。本研究中,相比胡子鯰,鯽幼魚對(duì)烏鱧暴露的反應(yīng)更為敏感,可能由于相比胡子鯰,烏鱧口裂更大,對(duì)鯽幼魚的威脅更為嚴(yán)重。胡子鯰雖然覓食行為非常活躍,但其口裂較小,而鯽自身體高較高。在本研究中鯽的體高與胡子鲇口裂非常接近,因此胡子鯰對(duì)其脅迫相對(duì)有限[2]。可見,本研究中鯽幼魚針對(duì)不同捕食者做出不同的反捕食響應(yīng)。另外值得注意的是,綜合鯽幼魚的多種反捕食策略,該種魚的確存在僅體現(xiàn)形態(tài)和快速啟動(dòng)逃逸能力的可塑性響應(yīng)以確保安全,而放棄反捕食行為以保障生長(zhǎng)的現(xiàn)象,這種現(xiàn)象從長(zhǎng)遠(yuǎn)看對(duì)于其提高生存適合度是有幫助的。因?yàn)閷?duì)于魚類而已,通過持續(xù)生長(zhǎng)獲得的更大的體型會(huì)使其在應(yīng)對(duì)種內(nèi)、種間競(jìng)爭(zhēng),以及各種環(huán)境脅迫(包括捕食脅迫)時(shí)更為有利[52- 54]。
有研究者認(rèn)為,當(dāng)多種捕食脅迫同時(shí)存在時(shí),多重捕食者對(duì)獵物帶來的風(fēng)險(xiǎn)效應(yīng)可能有以下4種:(1)重疊(Additive,捕食者A與B同時(shí)存在時(shí)的風(fēng)險(xiǎn)與A或B單獨(dú)存在時(shí)相同)、(2)風(fēng)險(xiǎn)增強(qiáng)(Risk enhancement,捕食者A和B同時(shí)存在時(shí)的風(fēng)險(xiǎn)大于A或B單獨(dú)存在的情況)、(3)風(fēng)險(xiǎn)削弱(Risk reduction,捕食者A和B同時(shí)存在時(shí)風(fēng)險(xiǎn)小于A或B單獨(dú)存在的情況)、(4)替換(Substitutable,捕食者A+A與捕食者B+B、捕食者A+B的風(fēng)險(xiǎn)相同)[17]。本研究中,雙捕食者暴露組所體現(xiàn)的風(fēng)險(xiǎn)效應(yīng)甚至低于烏鱧暴露組,如果參照上述觀點(diǎn),本研究中的多捕食者效應(yīng)體現(xiàn)為第三種情況:風(fēng)險(xiǎn)削弱。這可能是由于胡子鯰和烏鱧相互干擾所致。這兩種捕食者大小接近,均不能吃掉對(duì)方,但在實(shí)驗(yàn)過程中經(jīng)常可以觀察到兩種魚之間的相互追逐,這就反而降低了兩種捕食者對(duì)鯽幼魚的捕食脅迫。
綜上所述,本研究中鯽幼魚表現(xiàn)出對(duì)烏鱧的反應(yīng)最為明顯,可能與烏鱧口裂較大,給鯽幼魚帶來的威脅更大有關(guān)。另外,應(yīng)對(duì)捕食脅迫,鯽幼魚僅在形態(tài)和快速啟動(dòng)逃逸能力上表現(xiàn)出反捕食響應(yīng),其反捕食行為的喪失可能是對(duì)生長(zhǎng)的妥協(xié),這對(duì)于其適應(yīng)生境中長(zhǎng)期存在的捕食脅迫可能至關(guān)重要。
猜你喜歡
作文·小學(xué)低年級(jí)(2025年2期)2025-02-13 00:00:00
小雪花·小學(xué)生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學(xué)低年級(jí)(2024年2期)2024-04-29 00:00:00
作文·小學(xué)低年級(jí)(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(bào)(2022年4期)2022-08-09 08:52:06
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
