黑龍港苜蓿根際土壤真菌群落結(jié)構(gòu)及多樣性分析
2021-07-01 10:13:06劉震徐玉鵬王秀領(lǐng)黃素芳趙忠祥閻旭東
江蘇農(nóng)業(yè)科學(xué) 2021年10期
關(guān)鍵詞:群落結(jié)構(gòu)
劉震 徐玉鵬 王秀領(lǐng) 黃素芳 趙忠祥 閻旭東
摘要:為明確黑龍港地區(qū)苜蓿土壤微生物多樣性,利用高通量基因測序技術(shù)對多年生苜蓿土壤及對照土壤微生物真菌ITS區(qū)進行測定,并分析了不同處理間土壤微生物群落特點。結(jié)果表明,對照處理獲得真菌OTUs 523個,四年生苜蓿處理OTUs 742個。物種分類顯示對照處理隸屬12門22綱56目107科146屬,四年生苜蓿處理隸屬10門28綱60目126科186屬。各處理水平下均以子囊菌門(Ascomycota)為優(yōu)勢真菌種群,其相對豐度分別為54.69%和61.81%。苜蓿多年建植后可以增加土壤中腐生菌類數(shù)量,加快腐殖化進程,有利于提高土壤肥力及有機質(zhì)含量,同時也可以降低擔(dān)子菌門等致病菌的相對豐度,對減少植物土傳病害起到積極作用。
關(guān)鍵詞:黑龍港;苜蓿;土壤微生物;真菌;群落結(jié)構(gòu);多樣性分析
中圖分類號:S154.3 ? 文獻標(biāo)志碼: A ?文章編號:1002-1302(2021)10-0197-04
苜蓿是我國北方重要的飼料作物,擁有高蛋白、生物固氮能力強及適應(yīng)能力強等特性,近年來已成為北方調(diào)整農(nóng)業(yè)種植結(jié)構(gòu)的重要產(chǎn)業(yè)[1]。同時,常年種植苜蓿可以顯著改善土壤物理結(jié)構(gòu),Eynard等研究表明,同耕地土壤相比,草地土壤團聚體穩(wěn)定性約高17%[2]。 Barber通過多年試驗得出結(jié)論,長期種植苜蓿可提高土壤水穩(wěn)性團聚體指數(shù),該指數(shù)同苜蓿種植年限呈正相關(guān)關(guān)系[3]。不僅如此,種植苜蓿還會影響土壤養(yǎng)分組成,羅珠珠等研究發(fā)現(xiàn),苜蓿生長達一定年限后,可以顯著提高土壤表層輕組有機碳[4];黃祥豐等研究表明,種植苜蓿可以提高0~60 cm各土層有機質(zhì)、全氮、堿解氮含量[5]。然而,關(guān)于苜蓿對土壤的影響主要集中于土壤養(yǎng)分時間及空間變化,對土壤微生物研究較少,吳建峰研究發(fā)現(xiàn)土壤微生物可以促進植物根系吸收養(yǎng)分[6],同時土壤微生物也是土壤生態(tài)功能的重要指示物[7]。因此,研究微生物組成及其特性有重要意義。
高通量測序技術(shù)作為新一代測序技術(shù),通過基因序列對土壤微生物進行分析,在分析微生物群落結(jié)構(gòu)時有巨大優(yōu)勢,能更好地揭示土壤中微生物群落及構(gòu)成[8]。本研究通過高通量測序方法對滄州市農(nóng)林科學(xué)院試驗基地土壤中的真菌群落構(gòu)成及豐度進行了分析,為苜蓿改良土壤生態(tài)環(huán)境、提高土壤肥力提供了理論依據(jù)。
1 材料與方法
1.1 試驗材料
2018年7月15日在河北省滄州市獻縣北張白村滄州市農(nóng)林科學(xué)院試驗基地(116.36E,38.31N)選擇四年生苜蓿根際土壤(Y4)作為研究對象,同時選取該研究對象周邊未耕作土壤作為對照(CK)。采用五點法取樣,每個處理按照梅花形確定5個采樣點。每個采樣點內(nèi)選取生長良好的苜蓿樣品,去除其周邊及表層雜草后,采用抖落法取水平深度 0~20 cm苜蓿根際土壤約10 g為1個采樣點,待5個采樣點全部取完后充分混勻,采用四分法選取混合均勻的土壤樣品保存為1份土壤待測樣品。將待測樣品裝入無菌密封袋,放入裝有干冰的泡沫箱中保存。待全部樣品采集完成后運輸回試驗室,超低溫冰箱-80 ℃保存。
1.2 試驗方法
1.2.1 土壤樣品DNA提取及測序 將土壤提取基因組后采用NanoDrop-ND1000分光光度儀測定總DNA濃度,并利用1%瓊脂糖凝膠電泳檢測總DNA提取質(zhì)量。選擇質(zhì)量合格的DNA樣品進行真菌ITS1區(qū)測序,測序工作由北京百邁客科技有限公司完成,使用Illumina-HiSeq平臺進行測序。
1.2.2 DNA質(zhì)量評價 去除低質(zhì)量Reads,根據(jù)PE數(shù)據(jù)之間Overlap關(guān)系將成對的Reads拼接成一條序列。去除Tags兩端的Barcode序列及引物序列,去除嵌合體及其短序列等后得到高質(zhì)量的Clean Tags。然后對拼接好的序列進行進一步的去除嵌合體和段序列得到優(yōu)質(zhì)序列。
1.2.3 生物信息分析 使用QIIME 1.8.0[9]軟件中的 UCLUST[10]對Tags在97%相似度水平下獲得OTU,并基于真菌分類學(xué)數(shù)據(jù)庫對OTU進行分類學(xué)注釋,統(tǒng)計各樣品OTU中的豐度信息。
2 結(jié)果與分析
2.1 序列統(tǒng)計和微生物多樣性
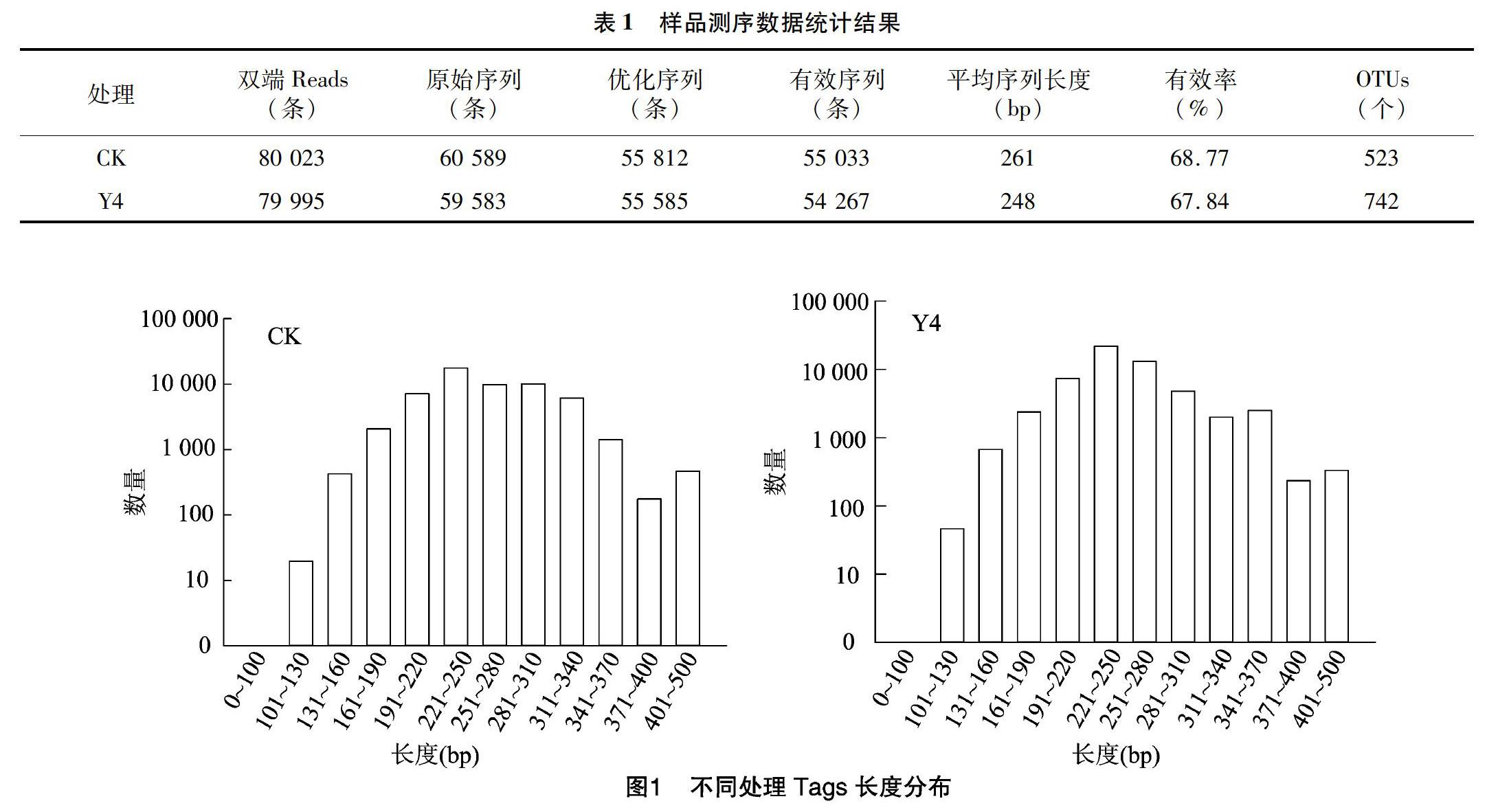
經(jīng)提取后,四年生苜蓿土壤(Y4)DNA濃度為2.5 ng/μL,D260 nm/280 nm值為4.18,D260 nm/230 nm值為0.03,對照土壤(CK)DNA濃度為1.9 ng/μL,D260 nm/280 nm值為2.88,D260 nm/230 nm值為0.08。各處理土壤樣品符合PCR檢測標(biāo)準(zhǔn),進一步以行擴增,其產(chǎn)物經(jīng)過高通量測序,樣品測序數(shù)據(jù)統(tǒng)計結(jié)果見表1,Tags長度分布見圖1。
2.2 對照土壤真菌群落組成
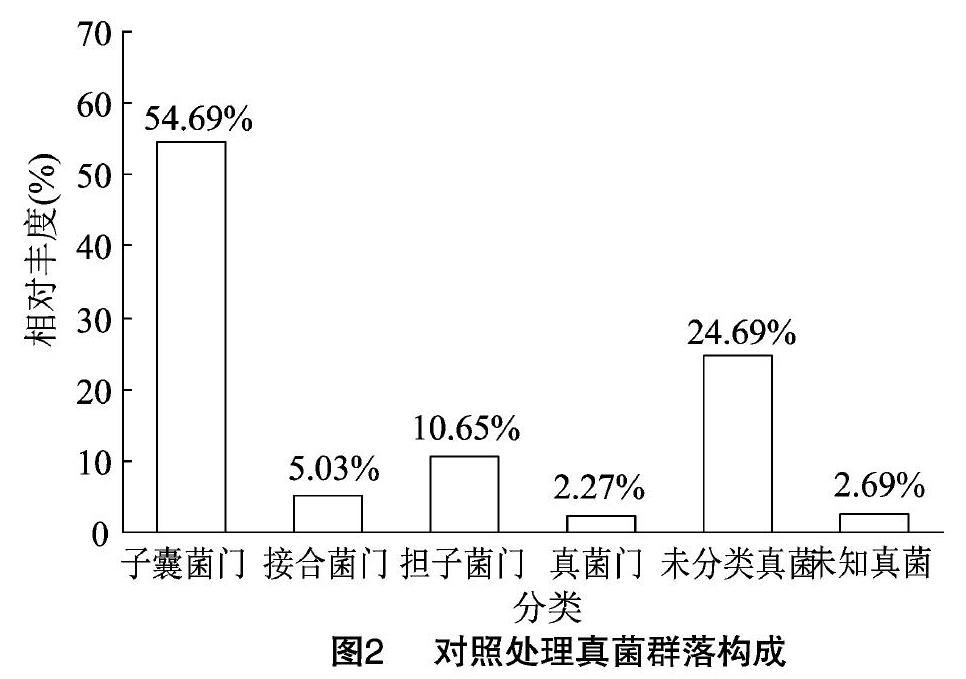
通過對照土壤真菌OTUs分析結(jié)果可以看出,其真菌群落隸屬于12門22綱56目107科146屬。由圖2可見,在門水平分類上至少隸屬于12個不同的真菌門,其中,相對豐度≥2%的有子囊菌門、接合菌門、擔(dān)子菌門、真菌門,其相對豐度分別為54.69%、5.03%、10.65%、2.27%;另外還有未能分類的真菌和未知真菌,其相對豐度分別為24.69%和2.69%。
在綱水平上至少隸屬于22個綱,相對豐度≥1%的有12個,其名稱分別為糞殼菌綱(30.57%)、座囊菌綱(12.84%)、散囊菌綱(8.09%)、酵母菌(2.16%)、錘舌菌綱(1.03%)、新囊菌綱(2.27%)、孢霉綱(3.20%)、毛霉綱(1.83%)、傘菌綱(9.36%)、節(jié)擔(dān)菌綱(1.29%)、未分類真菌(24.69%)和未知真菌(2.69%)。
在目分類水平上隸屬于56目,相對豐度≥1%的有12個,分別是肉座菌目(18.23%)、散囊菌目(7.13%)、糞殼菌目(5.85%)、格孢腔菌目(6.05%)、煤炱目(4.36%)、牛肝菌目(3.37%)、被孢霉目(3.20%)、新麗鞭毛菌目(2.27%)、球團目(2.11%)、毛霉目(1.83%)、未分類真菌(27.18%)、未知真菌(18.43%)。
2.3 四年生苜蓿處理土壤真菌生物多樣性
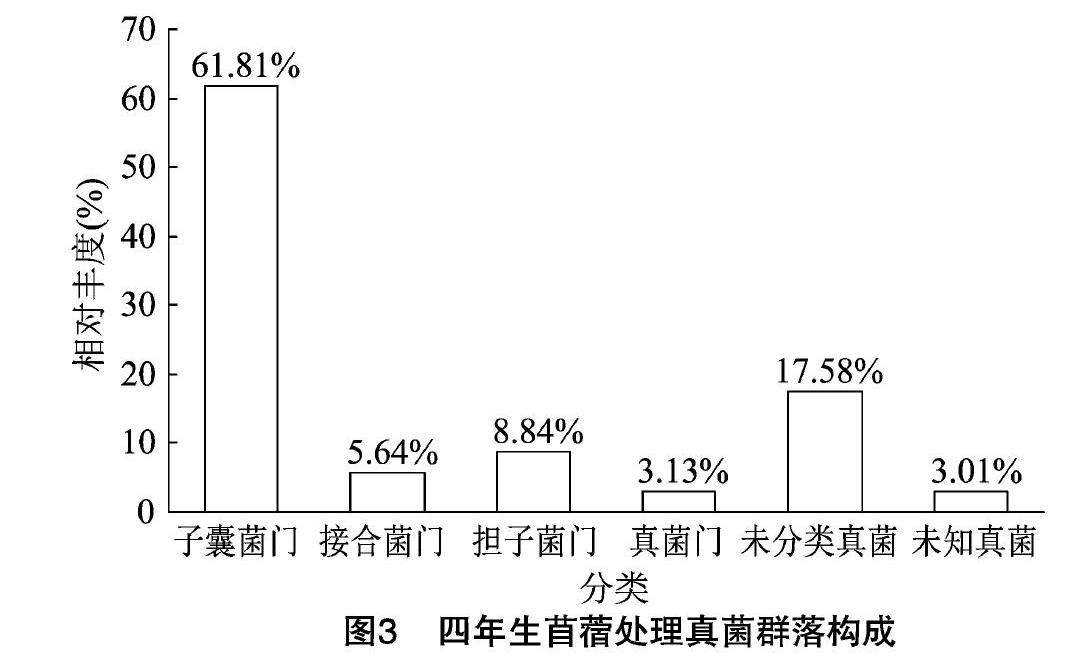
通過對四年生苜蓿土壤真菌OTUs分析結(jié)果可以看出,其真菌群落隸屬于10門28綱60目126科186屬。由圖3可見,在門水平分類上至少隸屬于10個不同的真菌門,其中,相對豐度≥3%的有子囊菌門、接合菌門、擔(dān)子菌門、真菌門,其相對豐度分別為61.81%、5.64%、8.84%、3.13%;另外還有未能分類的真菌和未知真菌,其相對豐度分別為24.69%和2.69%。
在綱水平上至少隸屬于28個綱,其中,相對豐度≥1%的有12個,其名稱分別為糞殼菌綱(37.59%)、座囊菌綱(11.11%)、散囊菌綱(9.73%)、酵母菌(2.03%)、錘舌菌綱(1.35%)、新囊菌綱(3.13%)、孢霉綱(3.20%)、毛霉綱(2.44%)、傘菌綱(7.32%)、節(jié)擔(dān)菌綱(1.52%)、未分類真菌(17.58%)和未知真菌(3.01%)。
在目分類水平上隸屬于60目,其中,相對豐度≥1% 的有12個,其名稱分別是肉座菌目(22.23%)、散囊菌目(8.99%)、糞殼菌目(8.08%)、格孢腔菌目(5.05%)、煤炱目(4.47%)、牛肝菌目(4.08%)、被孢霉目(3.20%)、新麗鞭毛菌目(3.13%)、球團目(2.27%)、毛霉目(2.44%)、未分類真菌(20.73%)、未知真菌(15.35%)。
2.4 Alpha和Beta多樣性分析
Alpha多樣性(表2)分析可以看出,Y4處理OUT數(shù)量為710,較CK處理OUT數(shù)量提高19.13%,ACE指數(shù)和Chao1指數(shù)也分別提高了19.21%和18.78%,說明Y4土壤中真菌微生物種類及豐度均大于CK土壤。Shannon指數(shù)值越大,說明樣品中物種多樣性越高,結(jié)合Simpson和Shannon指數(shù)可以看出,Y4土壤中真菌的種類也高于CK土壤,結(jié)果同OUT等指數(shù)得出的結(jié)論相似。
而由圖4可見,PCA分析可以代表89.70%的主要信息,其中可以明顯看出CK與Y4處理距離較遠,差異性較大;而通過Beta多樣性組間差異分析也可以直觀反映出CK與Y4處理組間差異較大,且達到極顯著水平。
3 結(jié)論與討論
土壤中含有大量組成結(jié)構(gòu)復(fù)雜的微生物群落[8],這些群落在地球化學(xué)循環(huán)中起重要的作用[11-13], 不僅影響土壤有機質(zhì)分解、土壤養(yǎng)分轉(zhuǎn)化
及運移,同時在土壤腐殖質(zhì)形成、各類土壤污染物的分解以及植物對土壤養(yǎng)分的吸收過程中也發(fā)揮一定的功能。通常可以用土壤微生物群落構(gòu)成來反映土壤微生物多樣性,也在一定程度上體現(xiàn)了土壤生態(tài)系統(tǒng)的穩(wěn)定性[14]。一般來說,微生物群落組成的復(fù)雜程度與土壤抵御不良生態(tài)環(huán)境的能力成正比。本試驗對多年生苜蓿土壤及對照土壤中真菌群落組成進行了分析,發(fā)現(xiàn)常年種植苜蓿在門水平真菌分類相對明確,但在綱水平及目水平下,仍有較大比例真菌不能準(zhǔn)確分類。
測序結(jié)果表明,對照及四年生苜蓿土壤中均檢測到子囊菌門、接合菌門、擔(dān)子菌門和真菌門,群落組成相對簡單。子囊菌門為土壤中的優(yōu)勢群落,對照和四年生苜蓿處理子囊菌門相對豐度分別為54.69%和61.81%,四年生苜蓿的提高7.12百分點;結(jié)核菌門、真菌門及未知真菌無明顯變化;擔(dān)子菌門相對豐度分別為10.65%和8.84%,四年生苜蓿的下降了1.81百分點。同對照相比,四年生苜蓿中有部分未分類真菌被檢測出,導(dǎo)致該處理中未分類真菌相對豐度下降了7.11百分點。
本試驗研究結(jié)果表明,鹽堿條件并不是子囊菌門真菌生存的限制因素,子囊菌門真菌群落可以在鹽堿條件下生存,甚至進一步擴大群落[15-16],該結(jié)果同李巖等的研究結(jié)果[17-18]相似。子囊菌門為土壤中優(yōu)勢真菌群落,其群落主要由腐生菌構(gòu)成。腐生菌可以通過分泌多種酶以體外方式分解土壤中的有機質(zhì)及動植物尸體,形成二氧化碳、無機鹽和簡單的有機物,為農(nóng)田土壤中的植物提供養(yǎng)分,是土壤養(yǎng)分循環(huán)過程中的重要真菌,說明在黑龍港鹽堿地區(qū)種植苜蓿可以提高子囊菌門真菌群落相對豐度,進一步改善土壤養(yǎng)分環(huán)境。同時,本研究還發(fā)現(xiàn)土壤中擔(dān)子菌門相對豐度降低,擔(dān)子菌門中一些微生物可以引起作物黑穗病、銹病以及一些其他森林和植物的病害,該門類真菌相對豐度的降低對減少作物病害也起到了一定積極的作用。本研究還發(fā)現(xiàn),在多年生苜蓿土壤中肉座菌目及糞殼科目菌類相對豐度有顯著提高,分別提高了4.00百分點和2.23百分點,這2類真菌的主要作用是降解腐爛有機質(zhì)及纖維素[19]。
綜上所述,本試驗通過高通量測序技術(shù)分析了長期種植苜蓿土壤中真菌群落結(jié)構(gòu)多樣性及結(jié)構(gòu)性,發(fā)現(xiàn)多年種植苜蓿后,土壤真菌群落結(jié)構(gòu)發(fā)生了改變,腐殖真菌群落相對豐度提高,腐殖化進程加快,可利于提高土壤肥力及有機質(zhì)。但是由于根際土壤微生物結(jié)構(gòu)復(fù)雜,受土壤物理結(jié)構(gòu)、施肥方式等影響較大,不同作物根系分泌物對微生物影響也不盡相同[20-24]。因此,苜蓿種植對土壤微生物真菌影響的主要因素和機制尚未完全明了,還需開展進一步研究。
參考文獻:
[1]趙海明,謝 楠,劉貴波,等. 紫花苜蓿品種在河北低平原區(qū)的引進篩選試驗研究[J]. 中國草地學(xué)報,2006,28(3):16-20.
[2]Eynard A,Schumacher T E,Lindstrom M J,et al. Aggregate sizes and stability in cultivated South Dakota Prairie Ustolls and Usterts[J]. Soil Science Society of America Journal,2004,68(4):1360-1365.
[3]Barber S A. The influence of alfalfa,bromegrass,and corn on soil aggregation and crop yield[J]. Soil Science Society of America Journal,1959,23(4):258-259.
[4]羅珠珠,李玲玲,牛伊寧,等. 土壤團聚體穩(wěn)定性及有機碳組分對苜蓿種植年限的響應(yīng)[J]. 草業(yè)學(xué)報,2016,25(10):40-47.
[5]黃祥豐,向地奎,賈黎明,等. 間作苜蓿對毛白楊紙漿林生長和地力的影響[J]. 南京林業(yè)大學(xué)學(xué)報(自然科學(xué)版),2013,37(4):39-44.
[6]吳建峰,林先貴. 土壤微生物在促進植物生長方面的作用[J]. 土壤,2003,35(1):18-21.
[7]王繼萍,龐碧琳,于亞軍. 不同復(fù)墾年限煤矸山土壤微生物群落特征及其指示作用[J]. 干旱區(qū)資源與環(huán)境,2017,31(9):140-144.
[8]牛世全,龍 洋,李海云,等. 應(yīng)用IlluminaMiSeq高通量測序技術(shù)分析河西走廊地區(qū)鹽堿土壤微生物多樣性[J]. 微生物學(xué)通報,2017(9):2067-2078.
[9]Caporaso J G,Kuczynski J,Stombaugh J,et al. QIIME allows analysis of high-throughput community sequencing data[J]. Nature Methods,2010,7(5):335-336.
[10]Edgar R C. Search and clustering orders of magnitude faster than BLAST[J]. Bioinformatics,2010,26(19):2460-2461.
[11]van der Heijden M G A,Bardgett R D,van Straalen N M. The unseen majority:soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems[J]. Ecology Letters,2008,11(3):296-310.
[12]Fuhrman J A,F(xiàn)uhrman J A. Microbial community structure and its functional implications[J]. Nature,2009,459(7244):193-199.
[13]張 晟,張徐潔,趙 遠,等. 不同溫度制備的水稻秸稈生物炭對稻田土壤固碳減排及微生物群落結(jié)構(gòu)的影響[J]. 江蘇農(nóng)業(yè)學(xué)報,2019,35(5):1102-1111.
[14]張紅娟,陳 麗,趙 芳,等. 山西臨汾小麥—玉米輪作田土壤微生物多樣性分析[J]. 中國植保導(dǎo)刊,2018,38(11):9-15.
[15]韓世忠,高 人,馬紅亮,等. 中亞熱帶森林土壤真菌多樣性的案例研究[J]. 熱帶亞熱帶植物學(xué)報,2015,23(3):343-352.
[16]韓世忠,高 人,馬紅亮,等. 建甌萬木林自然保護區(qū)兩種森林類型土壤真菌多樣性[J]. 生態(tài)學(xué)雜志,2015,34(9):2613-2620.
[17]李 巖,何學(xué)敏,楊曉東,等. 不同生境黑果枸杞根際與非根際土壤微生物群落多樣性[J]. 生態(tài)學(xué)報,2018,38(17):5983-5995.
[18]王艷云,郭篤發(fā). 黃河三角洲鹽堿地土壤真菌多樣性[J]. 北方園藝,2016(18):185-189.
[19]張 玥,胡雲(yún)飛,王樹茂,等. 茶園年限對根際土壤真菌群落結(jié)構(gòu)及多樣性的影響[J]. 應(yīng)用與環(huán)境生物學(xué)報,2018,24(5):972-977.
[20]Berendsen R L,Pieterse C M,Bakker P A. The rhizosphere microbiome and plant health[J]. Trends in Plant Science,2012,17(8):478-486.
[21]張 翔,宋水山,黃亞麗,等. 基于高通量測序分析河北省中南部地區(qū)耕地土壤細菌多樣性[J]. 華北農(nóng)學(xué)報,2018,33(4):196-203.
[22]張萌萌,敖 紅,張景云,等. 建植年限對紫花苜蓿根際土壤微生物群落功能多樣性的影響[J]. 草業(yè)科學(xué),2014,31(5):787-796.
[23]張萌萌,敖 紅,李 鑫,等. 桑樹/苜蓿間作對根際土壤酶活性和微生物群落多樣性的影響[J]. 草地學(xué)報,2015,23(2):302-309.
[24]張 超,周 旭,張 海,等. 蘋果專用肥對旱地果園土壤酶活性以及微生物多樣性的影響[J]. 生態(tài)學(xué)雜志,2017,36(12):3485-3492.
猜你喜歡
熱帶農(nóng)業(yè)科學(xué)(2016年11期)2017-01-21 14:59:21
熱帶農(nóng)業(yè)科學(xué)(2016年11期)2017-01-21 14:57:08
河北漁業(yè)(2015年12期)2015-12-21 14:37:31
河北漁業(yè)(2015年11期)2015-11-18 16:39:54
河北漁業(yè)(2015年11期)2015-11-18 16:37:49
河北漁業(yè)(2015年10期)2015-10-15 11:14:57
湖北農(nóng)業(yè)科學(xué)(2015年18期)2015-10-13 21:34:01
河北漁業(yè)(2015年7期)2015-07-21 11:06:52
河北漁業(yè)(2015年6期)2015-06-24 11:04:37
安徽農(nóng)學(xué)通報(2015年5期)2015-03-23 23:51:20
