不同基因型及培養(yǎng)基在燕麥花藥組織培養(yǎng)中的應用
2021-07-01 09:24:52田娟張曼孫墨可董玉迪王春龍郭來春魏黎明任長忠
江蘇農(nóng)業(yè)科學 2021年9期
田娟 張曼 孫墨可 董玉迪 王春龍 郭來春 魏黎明 任長忠
摘要:以JA3、M44、19A647燕麥資源為試驗材料,采用花藥離體培養(yǎng)方法,將3種材料的花藥接種于C17、W14培養(yǎng)基上,研究3種基因型在2種培養(yǎng)基上反應率、出愈率、轉(zhuǎn)移率和再生率的差異,以期篩選出適合燕麥花藥培養(yǎng)的基因型和誘導培養(yǎng)基。結(jié)果表明,在3個基因型中,JA3的反應率、出愈率、轉(zhuǎn)移率都是最高的,分別達到36.29%、2695%、86.93%,只有JA3獲得了再生植株,其綠苗率為0.10%,可見JA3適合用于花藥組織培養(yǎng),可以作為燕麥花藥培養(yǎng)的橋梁材料;C17培養(yǎng)基中的愈傷組織比較緊湊,質(zhì)量較好,W14培養(yǎng)基中的愈傷組織不易聚集長大,質(zhì)量較差,從W14培養(yǎng)基中共獲得6株白化苗,從C17培養(yǎng)基中獲得2株白化苗、2株綠苗。由研究結(jié)果可以看出,燕麥花藥愈傷組織誘導和再分化在基因型之間的差異顯著,JA3適合作為燕麥花藥培養(yǎng)的橋梁品種,相較于W14培養(yǎng)基,C17為燕麥花藥培養(yǎng)的最佳培養(yǎng)基。
關鍵詞:燕麥;基因型;培養(yǎng)基;花藥組織培養(yǎng);愈傷組織
中圖分類號: S339.4+1;S512.604.3 ?文獻標志碼: A ?文章編號:1002-1302(2021)09-0049-04
燕麥(Avena sativa L.)是禾本科燕麥屬一年生草本植物,是世界上主要的糧食作物之一[1],它抗旱[2]、耐寒[3]、耐鹽堿[4-5]。燕麥蛋白的氨基酸種類齊全,配比合理[6],具有極高的營養(yǎng)、飼用價值和保健功效[7]。燕麥新品種主要采用引進選育、系統(tǒng)育種、品種間雜交等方式育成[8]。花藥培養(yǎng)育種是將花粉培育成單倍體植株,再經(jīng)染色體自然或人工加倍得到純合二倍體的一種育種方法,這種由染色體加倍產(chǎn)生的純合二倍體,在遺傳上非常穩(wěn)定,不發(fā)生性狀分離,因此,花培育種能夠縮短育種年限[9]。但是在實際育種中,燕麥花培育種方法應用得很少,主要是因為花藥組織培養(yǎng)的誘導率不高。誘導率不高的原因,主要是材料的基因型、培養(yǎng)基的種類和成分、預處理和培養(yǎng)條件等因素在不同程度上制約了燕麥花藥培養(yǎng)的效果[10]。由于燕麥選育受資源與技術(shù)的雙重制約,使得燕麥育種效率低下,品種更新?lián)Q代較慢,產(chǎn)業(yè)化發(fā)展滯后[11]。為了解決我國燕麥育種面臨的育種技術(shù)單一、方法陳舊、周期長、優(yōu)良品種少等問題,攻克燕麥花藥單倍體育種技術(shù)屏障勢在必行。關于燕麥花藥組織培養(yǎng)的報道已經(jīng)有很多[12-24],本試驗選用JA3、M44、19A647燕麥品種(系)作為試驗材料,比較C17、W14這2種誘導培養(yǎng)基在燕麥花藥培養(yǎng)過程中的反應率、出愈率、轉(zhuǎn)移率和再生率差異,以期篩選出適合燕麥花藥培養(yǎng)的基因型和誘導培養(yǎng)基,為提高燕麥花藥培養(yǎng)育種效率提供理論依據(jù)。
1 材料與方法
1.1 試驗材料
以編號為JA3、M44、19A647的燕麥品種(系)作為試驗對象,于2019年4月1日播種于吉林省白城市農(nóng)業(yè)科學院內(nèi)。在孕穗期采集幼穗后置于4 ℃冰箱中進行暗、冷處理7 d后取出,用75%乙醇消毒,選取小孢子為單核中晚期的小穗(小孢子時期通過醋酸楊紅染色鏡檢來確定),剝離出花藥放在直徑為3.5 cm的培養(yǎng)皿中,每個培養(yǎng)皿中放置30個花藥,于32 ℃處理5 d后轉(zhuǎn)移到28 ℃條件下進行暗培養(yǎng)。
1.2 愈傷組織的誘導
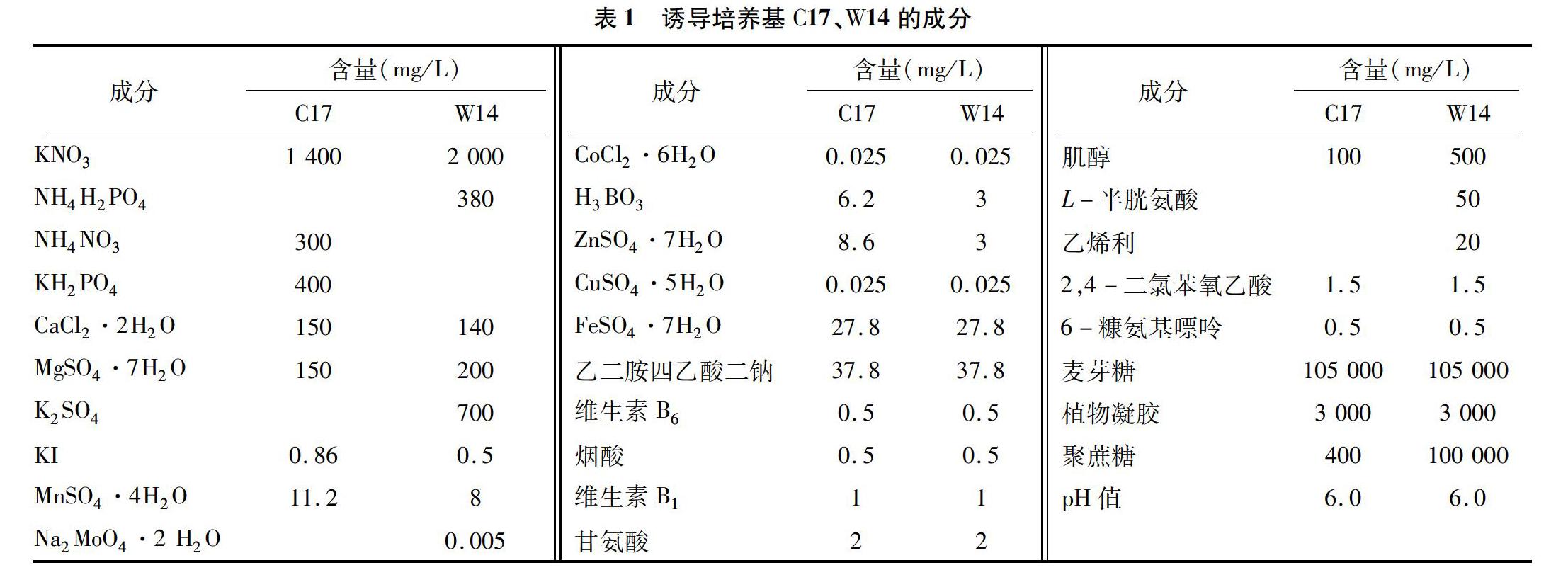
脫分化使用C17、W14這2種培養(yǎng)基,其中C17為固體培養(yǎng)基,W14為固液雙層培養(yǎng)基。W14上層是液體培養(yǎng)基,加入100 g/L聚蔗糖400、105 g/L麥芽糖等,培養(yǎng)基的具體成分見表1。
1.3 分化和生根培養(yǎng)
將誘導的愈傷組織轉(zhuǎn)移到再生培養(yǎng)基上,再生培養(yǎng)基是固體W14,另加2%蔗糖、0.3%植物凝膠,定容后調(diào)節(jié)pH值為6.0,于121 ℃滅菌20 min。將植物激素2 mg/L NAA、0.5 mg/L KT過濾消毒后加入再生培養(yǎng)基中。再生溫度設為22 ℃,采用16 h/d弱光光照。生根采用MS培養(yǎng)基[21],輔以0.2 mg/L NAA、2%蔗糖和0.3%植物凝膠。生根溫度為 25 ℃,光照時間為16 h/d。
1.4 數(shù)據(jù)統(tǒng)計與分析
使用WPS 2019軟件對試驗數(shù)據(jù)進行整理和統(tǒng)計分析,用IBM SPSS Statistics 19.0軟件進行單因素方差分析。
反應時間指從花藥接種到培養(yǎng)基中至花藥表面出現(xiàn)小泡的時間,出愈時間指從花藥接種到培養(yǎng)基中至出現(xiàn)愈傷組織的時間,其他計算公式如下:
反應率=反應的花藥數(shù)/接種的花藥數(shù)×100%;
出愈率=出現(xiàn)愈傷組織的花藥數(shù)/接種的花藥數(shù)×100%;
轉(zhuǎn)移率=能夠轉(zhuǎn)移的愈傷數(shù)/出現(xiàn)愈傷組織的花藥數(shù)×100%;
綠苗率=出現(xiàn)的綠苗數(shù)/接種的花藥數(shù)×100%;
白化苗率=出現(xiàn)的白化苗數(shù)/接種的花藥數(shù)×100%。
2 結(jié)果與分析
2.1 基因型對燕麥花藥愈傷組織誘導和再分化的影響
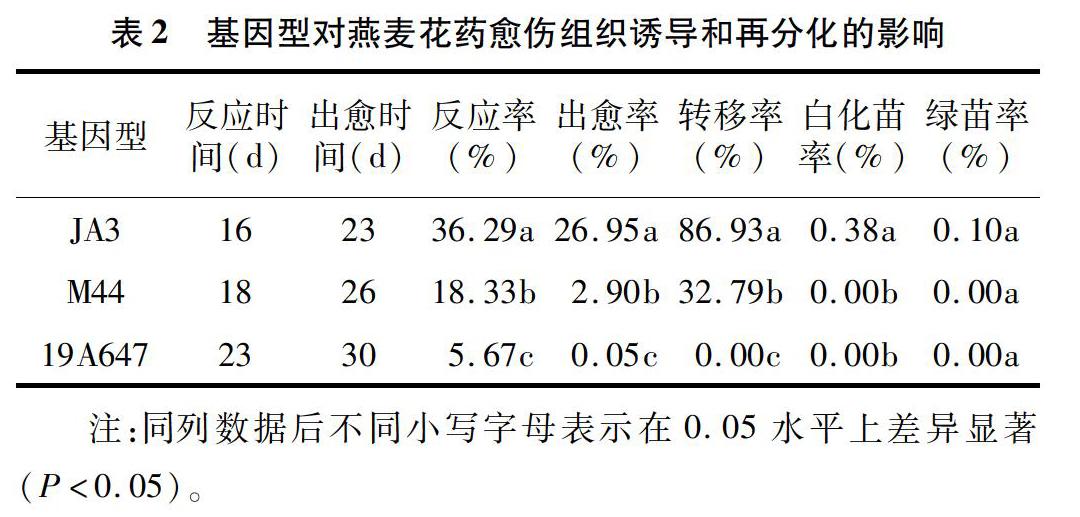
以JA3、M44、19A647燕麥品種(系)為材料,研究不同基因型對燕麥花藥愈傷組織誘導和再分化的影響。由表2可以看出,不同基因型燕麥的反應時間和出愈時間有所不同,但是3種基因型燕麥從反應到出愈約用1周時間,出愈時間為23~30 d不等。在相同培養(yǎng)條件下,燕麥花藥愈傷組織誘導和再分化在不同基因型之間的差異顯著。在供試的3個材料中,JA3的反應率、出愈率均最高,分別能達到36.29%、26.95%,而且JA3的轉(zhuǎn)移率能達到8693%,說明其愈傷組織活力較高,增殖速度快。在3個燕麥基因型中,只有JA3獲得了再生植株,其綠苗率為0.10%。以上結(jié)果說明,JA3適合用于花藥組織培養(yǎng),可以作為燕麥花藥培養(yǎng)的橋梁材料。19A647的反應率、出愈率都較低,沒有能夠轉(zhuǎn)移的愈傷組織,說明19A647不適合用于花藥培養(yǎng)。
2.2 培養(yǎng)基對燕麥花藥愈傷組織誘導和再分化的影響
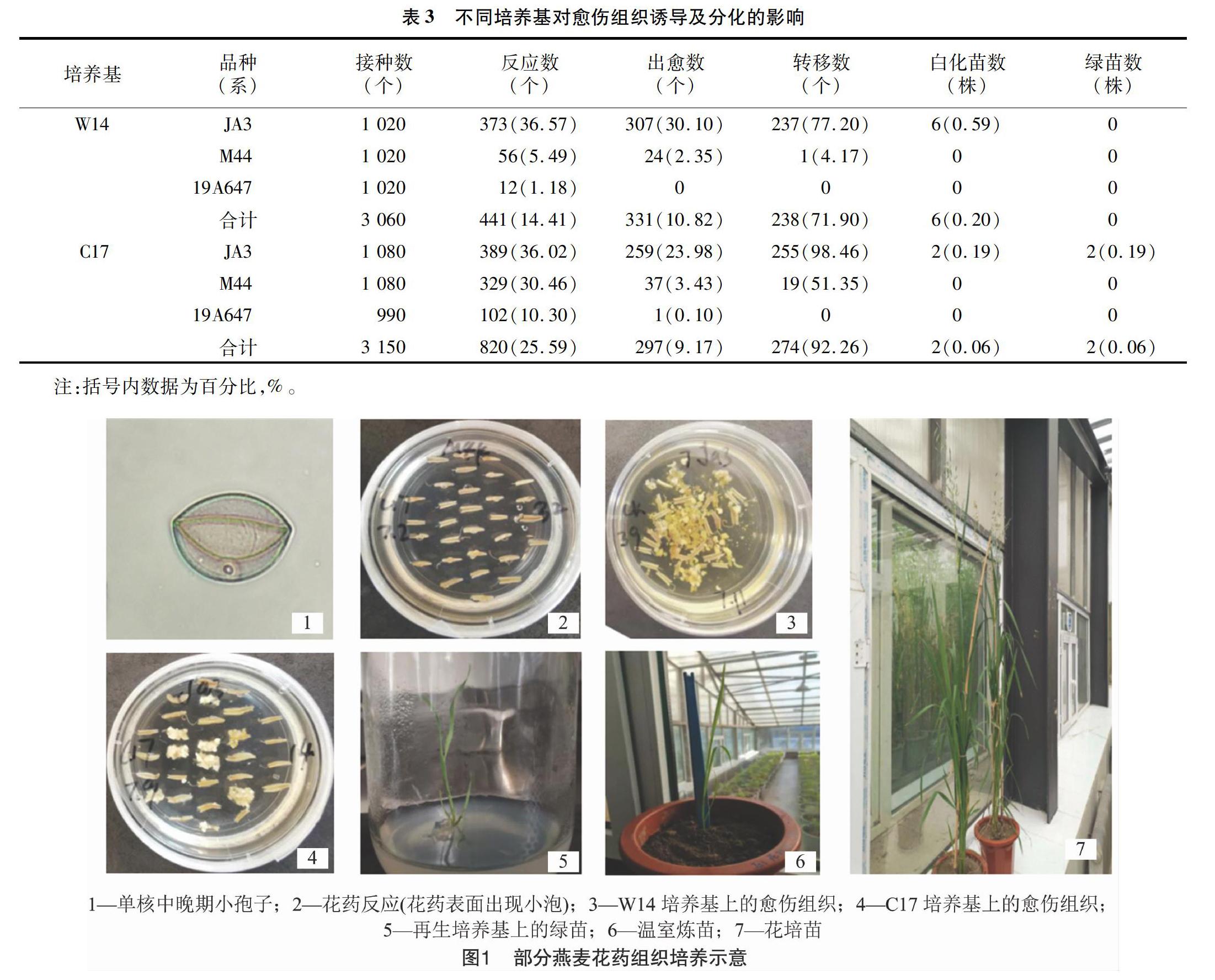
由表3可以看出,C17培養(yǎng)基中的花藥反應數(shù)較W14培養(yǎng)基高,并且每個基因型中反應數(shù)均超過10%。在出愈數(shù)方面,雖然W14培養(yǎng)基比C17培養(yǎng)基高,但是在轉(zhuǎn)移數(shù)方面,W14培養(yǎng)基不如C17培養(yǎng)基。由圖1可以看出,W14培養(yǎng)基出現(xiàn)的愈傷組織比較分散,不能達到轉(zhuǎn)移所需的大小。而C17培養(yǎng)基中的愈傷組織比較緊湊,容易長大,質(zhì)量比較好。從W14培養(yǎng)基中獲得了6株白化苗,從C17培養(yǎng)基中獲得了2株白化苗和2株綠苗,且每100個花藥的綠苗數(shù)達到0.06株。綜合愈傷組織質(zhì)量和獲得的綠苗數(shù),在本研究選用JA3、M44、19A647這3個燕麥材料進行花藥組織培養(yǎng)的過程中,C17培養(yǎng)基優(yōu)于W14培養(yǎng)基。
2.3 誘導培養(yǎng)基C17、W14成分的比較
培養(yǎng)基各成分的配比與優(yōu)化直接影響了燕麥花藥培養(yǎng)的出愈率。從表1可以看出,在C17、W14的培養(yǎng)基成分中,鐵鹽、有機成分、植物激素等含量完全相同。C17、W14中大量元素的種類不同,C17中的大量元素由KNO3、NH4NO3、KH2PO4、CaCl2·2H2O、MgSO4·7H2O提供,W14中的大量元素由KNO3、NH4H2PO4、CaCl2·2H2O、MgSO4·7H2O、K2SO4提供,相較于C17,W14中的微量元素(由KI、MnSO4·4H2O、H3BO3、ZnSO4·7H2O等化合物提供)含量都有不同程度的減少。C17是固體培養(yǎng)基,碳源只來自蔗糖,W14是固液雙層培養(yǎng)基,碳源來自蔗糖、麥芽糖,W14培養(yǎng)基中增加了L-半胱氨酸、乙烯利等。
3 討論與結(jié)論
3.1 基因型
誘導花粉形成愈傷組織是誘導花粉植株的第一步,在同樣的培養(yǎng)條件下,不同品種或雜種燕麥花藥產(chǎn)生花粉愈傷組織的頻率差異極大,有的高達48.5%,但有的則不產(chǎn)生花粉愈傷組織[22]。這種差異顯然是由花藥與花粉內(nèi)部的因素引起的,供體植株的基因型是影響花藥、花粉培養(yǎng)的關鍵因素,不同基因型的植株對培養(yǎng)的反應不同。本試驗結(jié)果表明,在相同培養(yǎng)條件下,燕麥花藥愈傷組織誘導和再分化在基因型之間的差異顯著,主要表現(xiàn)為出現(xiàn)愈傷組織的時間不同以及出愈率、綠苗分化率和白化苗分化率不同等方面。本研究結(jié)果與Kiviharju等的相關研究結(jié)果[23-25]一致。本研究所選的JA3燕麥在W14培養(yǎng)基上的出愈率達到30.10%,適合作為燕麥花藥培養(yǎng)的橋梁品種。
3.2 培養(yǎng)基
培養(yǎng)基的組成在胚胎發(fā)生中起著主要作用。燕麥花藥培養(yǎng)效率的提高,在某些程度上依賴于培養(yǎng)基成分、激素配比等。C17、W14培養(yǎng)基是廣泛用于燕麥花藥培養(yǎng)的2種培養(yǎng)基[13,26-29],這2種培養(yǎng)基的成分與激素含量存在一定差異。本研究對比了2種培養(yǎng)基誘導愈傷組織和綠苗分化的能力,并綜合愈傷組織的質(zhì)量、獲得的綠苗數(shù)得出,C17為最佳培養(yǎng)基,與Ponitka等的研究結(jié)果[30]一致,但是與Warchol等的研究結(jié)果[31]不一致。
本試驗中培養(yǎng)基差異很大,大量元素來源不同,微量元素含量不同,并且與C17相比,W14中添加了L-半胱氨酸、乙烯利等激素,分為固液2層,在液體層中加入了麥芽糖而不是蔗糖。這些成分的改變并沒有提高燕麥花藥愈傷組織的誘導率和質(zhì)量,但是可以看出,雙層培養(yǎng)基的優(yōu)點是花藥在培養(yǎng)早期可以從活性高的液體培養(yǎng)基中汲取營養(yǎng),從而提高花粉反應率,但是并沒有提高愈傷組織誘導率,而且由于花藥直接放在液體培養(yǎng)基上,出現(xiàn)的愈傷組織在培養(yǎng)基上來回游動,不能聚集長大,反而降低了愈傷組織質(zhì)量。由于本研究的培養(yǎng)基差異很大,究竟是大量元素、微量元素、糖源還是 L-半胱氨酸和乙烯利等激素對愈傷組織的影響,還有待進一步探討。
本試驗從多種角度發(fā)現(xiàn),在相同培養(yǎng)條件下,不同基因型植株的再生情況差異明顯。這種差異在很大程度上由基因型自身的再生能力決定,優(yōu)化再生體系僅能在一定程度上提高某些基因型的培養(yǎng)力,但并不能從根本上改變這種差距。對于不同培養(yǎng)基而言,相同基因型植株的出愈率差別不大。
參考文獻:
[1]鄭殿升.中國燕麥的多樣性[J]. 植物遺傳資源學報,2010,11(3):249-252.
[2]李 威. PEG脅迫下6種裸燕麥品種種子萌發(fā)期的抗旱性研究[J]. 種子,2014,33(5):38-41.
[3]柏曉玲,周青平,陳有軍,等. 燕麥幼苗對低溫脅迫的響應[J]. 草業(yè)科學,2016,33(7):1375-1382.
[4]姜 瑛,周 萌,吳 越,等. 不同燕麥品種耐鹽性差異及其生理機制[J]. 草業(yè)科學,2018,35(12):2903-2914.
[5]付鸞鴻,于 崧,于立河,等. 不同基因型燕麥萌發(fā)期耐鹽堿性分析及其鑒定指標的篩選[J]. 作物雜志,2018(6):27-35,174.
[6]許英一,王 宇,林 巍. 燕麥蛋白理化性質(zhì)研究[J]. 安徽農(nóng)業(yè)大學學報,2018,45(3):385-388.
[7]石振興,朱瑩瑩,任貴興. 燕麥中減肥降脂的功能成分研究進展[J]. 食品安全質(zhì)量檢測學報,2018,9(7):1567-1571.
[8]任長忠,崔 林,楊 才,等. 我國燕麥高效育種技術(shù)體系創(chuàng)建與應用[J]. 中國農(nóng)業(yè)科技導報,2016,18(1):1-6.
[9]王 煒,葉春雷,楊隨莊,等. 花藥培養(yǎng)技術(shù)在小麥種質(zhì)資源創(chuàng)制及育種中的應用[J]. 中國種業(yè),2018(11):25-29.
[10]田 娟,張 曼,董玉迪,等. 燕麥花藥組織培養(yǎng)影響因子研究進展[J]. 安徽農(nóng)業(yè)科學,2018,46(35):11-13.
[11]王 娟,李蔭藩,梁秀芝,等. 北方主栽燕麥品種種質(zhì)資源形態(tài)多樣性分析[J]. 作物雜志,2017(4):27-32.
[12]Rines H W. Oat anther culture:genotype effect on callus initiation and the production of a haploid plant[J]. Crop Science,1983,23(2):268-272.
[13]Kiviharju E,Puolimatka M,Pehu E. Regeneration of anther-derived plants of Avena sterilis[J]. Plant Cell,Tissue & Organ Culture,1997,48(2):147-152.
[14]Warchol M,Skrzypek E,Nowakowska A,et al. The effect of auxin and genotype on the production of Avena sativa L. doubled haploid lines[J]. Plant Growth Regulation,2016,78(2):155-165.
[15]Kiviharju E,Pehu E. The effect of cold and heat pretreatments on anther culture response of Avena sativa and A. sterilis[J]. Plant Cell,Tissue and Organ Culture,1998,54(2):97-104.
[16]Kiviharju E M,Tauriainen A A. 2,4-dichlorophenoxyacetic acid and kinetin in anther culture of cultivated and wild oats and their interspecific crosses:plant regeneration from A. sativa L.[J]. Plant Cell Reports,1999,18(7):582-588.
[17]Rines H W, Rieralizarazu O, Nunez V M, et al. Oat haploids from anther culture and from wide hybridizations[M]//Jain S M, Sopory S K, Veilleux R E, et al. In vitro haploid production in higher plants. Dordrecht:Kluwer, 1997,4:205-221.
[18]Jingsan S,Tiegang L,Sōndahl M R,et al. Anther culture of naked oat and the establishment of its haploid suspension cell[J]. Journal of Integrative Plant Biology,1991,33(6):417-420.
[19]Rines H W. Oat haploids from wide hybridization[J]. Doubled Haploid Production in Crop Plants,2003:155-159.doi:10.1007/978-94-017-1293-4_24.
[20]Kiviharju E,Moisander S,Laurila J,et al. Improved green plant regeneration rates from oat anther culture and the agronomic performance of some DH lines[J]. Plant Cell,Tissue and Organ Culture,2005,81(1):1-9.
[21]Murashige T,Skoog F. A revised medium for rapid growth and bio assays with tobacco tissue cultures[J]. Physiologia Plantarum,1962,15(3):473-497.
[22]楊 才,王秀英. 采用花藥單倍體育種方法育成花中21號莜麥新品種[J]. 河北北方學院學報(自然科學版),2005,21(3):39-42.
[23]Kiviharju E,Puolimatka M,Saastamoinen M,et al. Extension of anther culture to several genotypes of cultivated oats[J]. Plant Cell Reports,2000,19(7):674-679.
[24]S'lusarkiewicz-Jarzina A,Ponitka A. The effect of physical medium state on anther culture response in Polish cultivated oat (Avena sativa L.)[J]. Acta Biologica Cracoviensia(Series Botanica),2007,49(2):27-31.
[25]Noga A,Skrzypek E,Warchol M,et al. Conversion of oat (Avena sativa L.) haploid embryos into plants in relation to embryo developmental stage and regeneration media[J]. In Vitro Cellular & Developmental Biology-Plant,2016,52(6):590-597.
[26]Kiviharju E,Puolimatka M,Saastamoinen M,et al. The effect of genotype on anther culture response of cultivated and wild oats[J]. Agricultural and Food Science in Finland,1998,7(3):409-422.
[27]Kiviharju E,Moisander S,Tanhuanpaa P,et al. Oat anther culture and use of DH-lines for genetic mapping[J]. Methods of Molecular Biology,2017,1536:71-93.
[28]Sidhu P K,Davies P A. Regeneration of fertile green plants from oat isolated microspore culture[J]. Plant Cell Reports,2009,28(4):571-577.
[29]Ferrie A M R,Irmen K I,Beattie A D,et al. Isolated microspore culture of oat (Avena sativa L.) for the production of doubled haploids:effect of pre-culture and post-culture conditions[J]. Plant Cell,Tissue and Organ Culture,2014,116(1):89-96.
[30]Ponitka A,S'lusarkiewicz-Jarzina A. Regeneration of oat androgenic plants in relation to induction media and culture conditions of embry-like structures[J]. Acta Societatis Botanicorum Poloniae,2009,78(3):209-213.
[31]Warchol M,Czyczylo-Mysza I,Marcińska I,et al. Factors inducing regeneration response in oat (Avena sativa L.) anther culture[J]. In Vitro Cellular & Developmental Biology-Plant,2019,55(5):595-604.
