不同生境果園梨小食心蟲(chóng)各代成蟲(chóng)高峰期差異
2021-06-29 01:21:36沈志杰房明華洪文英劉濤沃林峰孫媛藍(lán)陳儀航鄧建宇
植物保護(hù) 2021年3期
沈志杰 房明華 洪文英 劉濤 沃林峰 孫媛 藍(lán)陳儀航 鄧建宇

摘要 :梨小食心蟲(chóng)Grapholita molesta (Busck)是桃、梨果園中最重要的食心害蟲(chóng)之一,僅依據(jù)往年的發(fā)生期和田間為害特征來(lái)指導(dǎo)防治具有滯后性。而性信息素具有高效、專一性強(qiáng)的特點(diǎn),可用于準(zhǔn)確監(jiān)測(cè)其成蟲(chóng)發(fā)生情況,以此推測(cè)成蟲(chóng)的卵高峰期和幼蟲(chóng)初孵期,科學(xué)地指導(dǎo)防治。本研究在3個(gè)不同生境果園中,利用性信息素監(jiān)測(cè)梨小食心蟲(chóng)成蟲(chóng)周年發(fā)生動(dòng)態(tài)。結(jié)果表明:2019年在杭州余杭地區(qū)3個(gè)不同生境果園,包括越冬代在內(nèi),平地桃園和平地梨園的梨小食心蟲(chóng)成蟲(chóng)發(fā)生7代,而坡地桃園只有6代。成蟲(chóng)始見(jiàn)于3月中旬,終見(jiàn)于10月下旬;3個(gè)果園由于生境不同,同一世代梨小食心蟲(chóng)高峰期發(fā)生時(shí)間和各世代高峰期間隔時(shí)間不盡相同。因此在不同生境果園利用性信息素監(jiān)測(cè)梨小食心蟲(chóng)成蟲(chóng)發(fā)生動(dòng)態(tài),有助于指導(dǎo)果園的適期防治。
關(guān)鍵詞 :梨小食心蟲(chóng); 監(jiān)測(cè); 不同生境果園; 種群動(dòng)態(tài); 高峰期
中圖分類號(hào):
S 436.612.22, S 433.4
文獻(xiàn)標(biāo)識(shí)碼: A
DOI: 10.16688/j.zwbh.2020044
Differences of adult peak periods of each generation of male oriental fruit moth,
Grapholita molesta (Busck) in different habitat orchards
SHEN Zhijie1, FANG Minghua2, HONG Wenying3, LIU Tao2, WO Linfeng2, SUN Yuan1,
LAN Chenyihang1, DENG Jianyu1*
(1.College of Agriculture and Food Science, Zhejiang A & F University, Hangzhou 311300, China;
2. Hangzhou Foundation Seed Farm of Zhejiang Province, Hangzhou 311115, China;
3. Hangzhou Plant Centre for Agricultural Technology Extension, Zhejiang Province, Hangzhou 310020, China)
Abstract
Grapholita molesta (Busck) is one of the most important fruit-boring pests in peach and pear orchards. Merely depending on calendars and the insect-damaged symptom in the field, the fruit farmers could not apply pesticide in time. Sex pheromone can be applied in monitoring the occurrence of male G.molesta adults because of its high efficiency and strong selectivity, hence peaks of egg laying and periods of newly hatched larvae can be predicted. Sex pheromones were used to monitor the population dynamic of male G.molesta adults in three orchards with different habitats in Yuhang district, Hangzhou in 2019. The results showed that there were seven generations of G.molesta adults both in a flat ground peach orchard and a flat ground pear orchard, and six generations in a sloping ground peach orchard in 2019. The adults were first trapped in mid of March and disappeared in late October. The peak period of G.molesta in the same generation and the peak interval of different generations were not the same. Therefore, it was of great significance to set up the pheromone-based monitoring of G.molesta for the pest control in different habitat orchards.
Key words
Grapholita molesta; monitoring; orchards with different habitats; population dynamic; peak period
梨小食心蟲(chóng)Grapholita molesta (Busck)屬鱗翅目,卷蛾科,小食心蟲(chóng)屬,是一種重要的食心害蟲(chóng),其寄主范圍較廣,包括桃、梨、櫻桃、李、杏、蘋果等許多果樹(shù),主要以幼蟲(chóng)蛀入嫩梢和果實(shí),形成蛀梢、蛀果,造成減產(chǎn)。梨小食心蟲(chóng)是浙江桃園最主要的蛀果害蟲(chóng),占比達(dá)到95%以上[1]。我國(guó)除西藏外,梨小食心蟲(chóng)廣泛分布于各水果產(chǎn)區(qū),北至黑龍江,南至廣東,1年發(fā)生3~7代不等[2],并且在日本、美國(guó)東西部、歐洲中西部、澳大利亞南部等地區(qū)水果產(chǎn)區(qū)均有為害[3]。梨小食心蟲(chóng)孵化不久后,便蛀入嫩梢或果中[45],不利于有效防治。
根據(jù)前期調(diào)查發(fā)現(xiàn),化學(xué)防治是浙江余杭地區(qū)防控梨小食心蟲(chóng)的主要措施,但果農(nóng)對(duì)梨小食心蟲(chóng)發(fā)生規(guī)律缺乏認(rèn)知,且沒(méi)有相應(yīng)的措施及時(shí)了解其種群動(dòng)態(tài),很難掌握最佳的施藥時(shí)間,從而經(jīng)常使用“保險(xiǎn)藥”,以保證防治效果,但這種施藥方式不利于保障果品安全和保護(hù)生態(tài)環(huán)境。性信息素具有高效、綠色、專一性強(qiáng)的特點(diǎn),可用于梨小食心蟲(chóng)監(jiān)測(cè)、誘殺和迷向。杜娟等報(bào)道梨小食心蟲(chóng)百果卵量和卵果率均與誘蛾量顯著相關(guān),可利用梨小食心蟲(chóng)誘蛾量推測(cè)梨果著卵量,當(dāng)以卵果率為1%或百果卵量1粒為防治指標(biāo)時(shí),性誘防治指標(biāo)分別為2.27頭和232頭[6]。陳香梅等也認(rèn)為可利用性誘劑測(cè)報(bào)成蟲(chóng)發(fā)生期,當(dāng)成蟲(chóng)高峰期后5~10 d是卵高峰期和幼蟲(chóng)初孵期[7],以此依據(jù)指導(dǎo)防治。為了探明余杭地區(qū)梨小食心蟲(chóng)種群發(fā)生規(guī)律以及在小范圍內(nèi)不同生境果園梨小食心蟲(chóng)的發(fā)生情況是否一致,本文利用性信息素監(jiān)測(cè)3個(gè)不同生境果園梨小食心蟲(chóng)成蟲(chóng)周年發(fā)生動(dòng)態(tài),以期明確其在當(dāng)?shù)氐陌l(fā)生規(guī)律,為科學(xué)防治提供理論依據(jù),同時(shí)為不同生境果園合理使用性信息素監(jiān)測(cè)提供參考。
1 材料與方法
1.1 試驗(yàn)地點(diǎn)
本研究地點(diǎn)位于浙江省杭州市余杭區(qū)瓶窯鎮(zhèn)毛元嶺村,是余杭區(qū)桃主產(chǎn)區(qū)之一。
3個(gè)果園分別為:平地桃園(30°24′26″N,119°56′25″E),株行距3 m×4 m,樹(shù)齡5年,面積約3 300 m2,品種為‘白麗,成熟期7月,海拔高度0 m,周邊種植水稻、玉米、蔬菜等作物,100 m外有連片大面積桃園。坡地桃園(30°25′17″N,119°56′34″E),株行距3 m×4 m,樹(shù)齡7年,面積約10 000 m2,品種為‘大觀1號(hào),成熟期5月,海拔高度100 m,位于北坡,東側(cè)緊靠一個(gè)面積約4 000 m2的桃園,其余周邊均為荒山。平地梨園(30°24′40″N,119°56′35″E),株行距3 m×3 m,樹(shù)齡7年,面積約3 300 m2,品種為‘翠冠,成熟期7月,海拔高度0 m,南側(cè)緊靠一個(gè)面積約2 000 m2的桃園,其余周邊種植水稻、玉米、蔬菜等作物。平地桃園距平地梨園約800 m、距坡地桃園約1 700 m,平地梨園和坡地桃園之間相隔約2 000 m。3個(gè)果園周年由果農(nóng)自行管理。
1.2 試驗(yàn)材料
誘捕器為三角形誘捕器配套白色黏蟲(chóng)板,購(gòu)于北京中捷四方生物科技股份有限公司,由1塊白色塑料板折疊成三棱柱形狀,長(zhǎng)24 cm,截面三角形寬18 cm、高18 cm,水平底面內(nèi)側(cè)固定黏蟲(chóng)板[8]。
梨小食心蟲(chóng)誘芯為實(shí)驗(yàn)室自制誘芯:將順-8-十二碳烯醇乙酸酯、反-8-十二碳烯醇乙酸酯、順-8-十二碳烯醇、反-8,反-10-十二碳二烯醇用正己烷溶解,配制成一定比例的梨小食心蟲(chóng)性信息素溶液,定量加入性信息素載體(“黑色橡膠帽”構(gòu)造)中,待正己烷揮發(fā)后,裝入密封袋,置于4℃冰箱中保存,田間持效期可達(dá)30 d。
1.3 試驗(yàn)方法
在平地桃園中設(shè)置3個(gè)梨小食心蟲(chóng)性信息素監(jiān)測(cè)誘捕器,相鄰誘捕器均間隔約30 m,懸掛高度約1.8 m。每周調(diào)查1次誘捕器中梨小食心蟲(chóng)蟲(chóng)量,每月更換1次誘芯,黏板覆蓋率超過(guò)50%或使用1月未達(dá)到50%覆蓋率的進(jìn)行更換。試驗(yàn)于2019年3月4日至11月5日進(jìn)行。坡地桃園設(shè)置梨小食心蟲(chóng)監(jiān)測(cè)誘捕器4個(gè),試驗(yàn)于2019年3月26日至11月5日進(jìn)行,其余試驗(yàn)方法同平地桃園。平地梨園試驗(yàn)方法同平地桃園。
1.4 數(shù)據(jù)處理
試驗(yàn)數(shù)據(jù)采用Excel 2010和SPSS 22.0軟件處理。誘蛾量=每次調(diào)查蟲(chóng)口總數(shù)/(誘捕器數(shù)×調(diào)查間隔天數(shù))。
2 結(jié)果與分析
2.1 不同桃園梨小食心蟲(chóng)種群動(dòng)態(tài)監(jiān)測(cè)
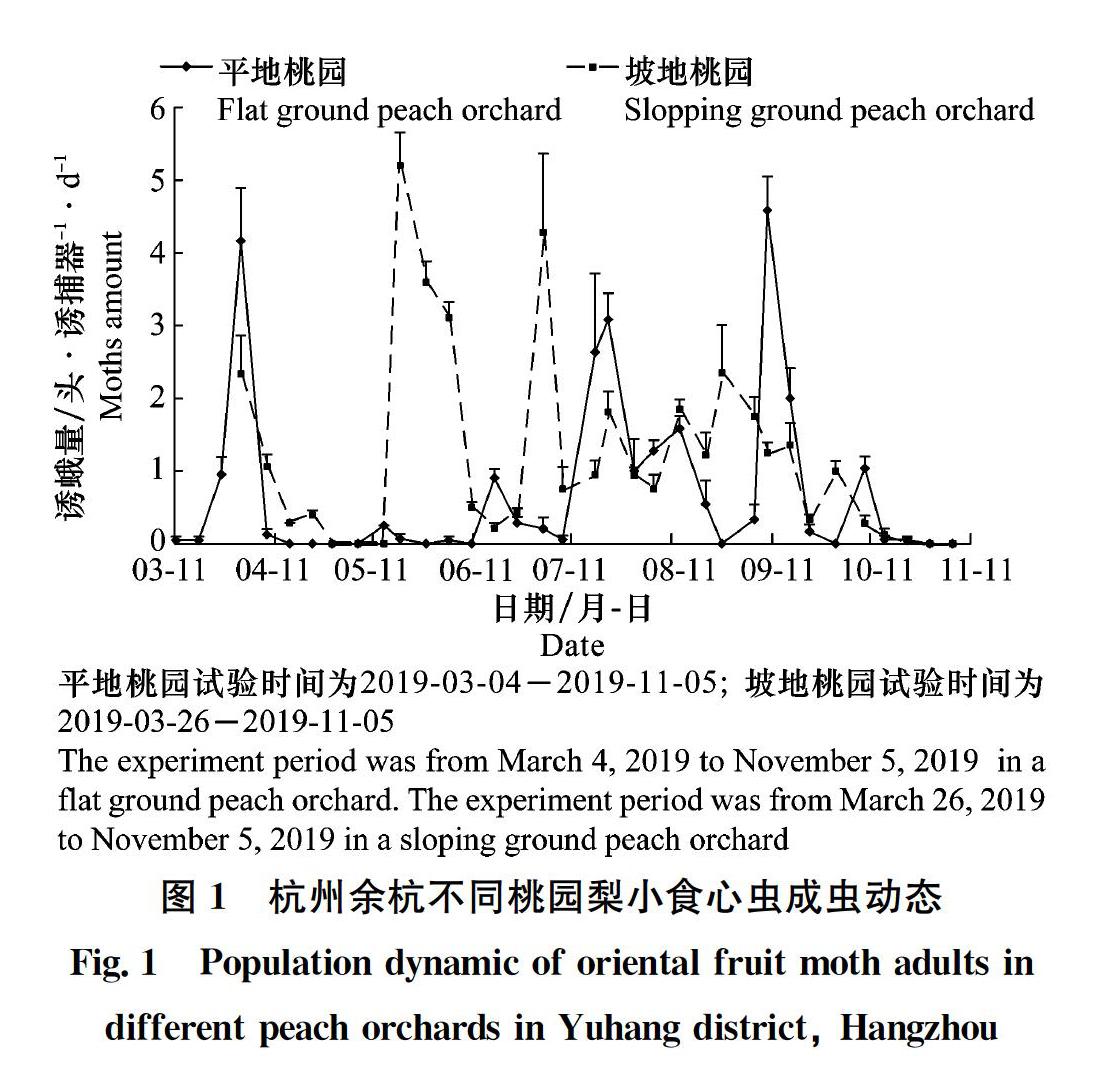
從圖1可看出,平地桃園2019年梨小食心蟲(chóng)雄成蟲(chóng)發(fā)生7代,坡地桃園發(fā)生6代。平地桃園成蟲(chóng)始見(jiàn)于3月中旬,越冬代雄成蟲(chóng)高峰期發(fā)生于3月底,誘蛾量較高;坡地桃園越冬代雄成蟲(chóng)在3月已發(fā)生,但未監(jiān)測(cè)到高峰期。平地桃園1~5代雄成蟲(chóng)高峰期均早于坡地桃園;1代、2代和4代誘蛾量均低于坡地桃園;而3代和5代誘蛾量則高于坡地桃園。成蟲(chóng)均終見(jiàn)于10月下旬。
2.2 平地梨園梨小食心蟲(chóng)種群動(dòng)態(tài)監(jiān)測(cè)
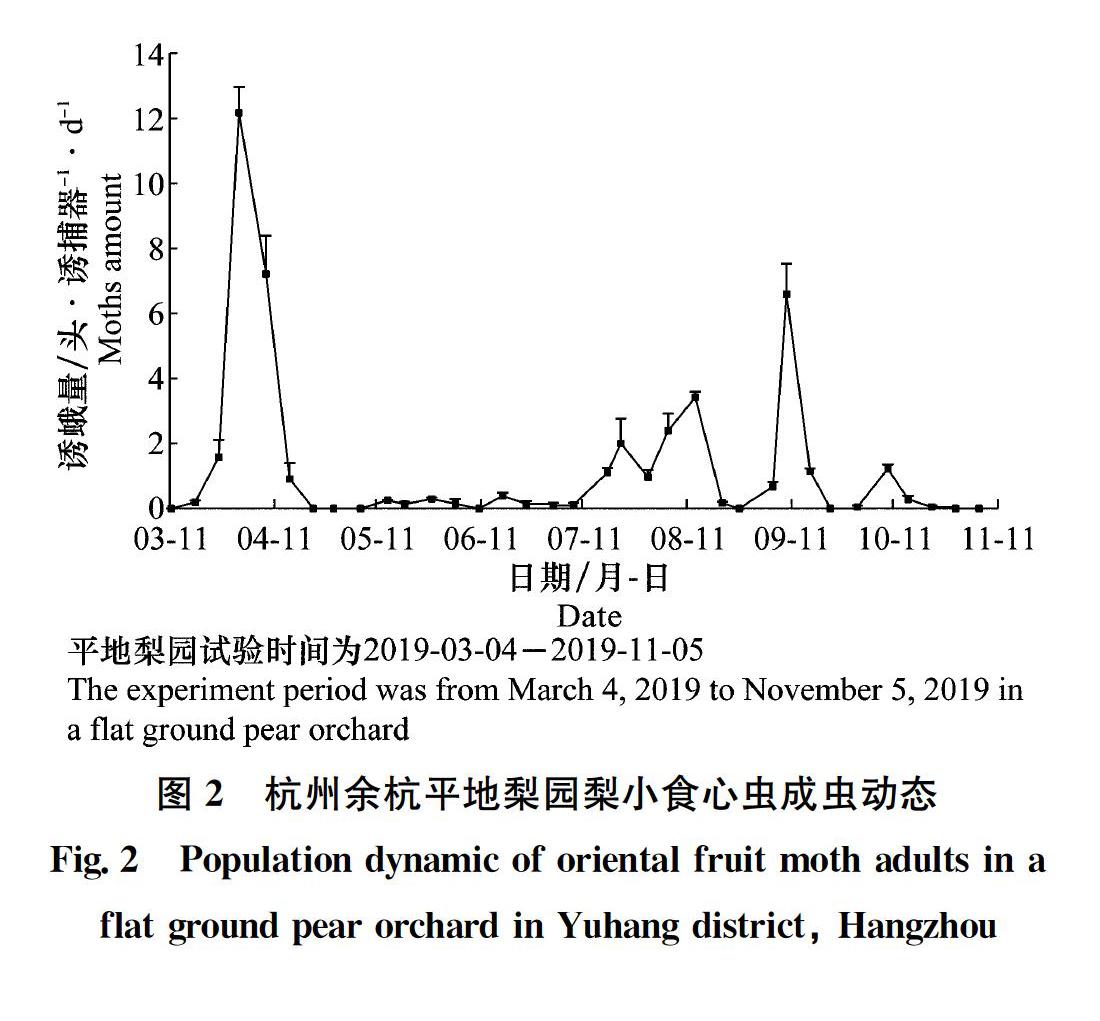
從圖2可看出,平地梨園2019年梨小食心蟲(chóng)雄成蟲(chóng)發(fā)生7代,梨小食心蟲(chóng)成蟲(chóng)始見(jiàn)于3月中旬,越冬代雄成蟲(chóng)高峰期為3月底,誘蛾量高于其他世代,越冬代成蟲(chóng)持續(xù)27 d;1代雄成蟲(chóng)高峰期為5月中下旬,2代雄成蟲(chóng)高峰期為6月中旬,誘蛾量均較低;3代雄成蟲(chóng)高峰期于7月中下旬,誘蛾量高于前2代;4代雄成蟲(chóng)高峰期為8月上中旬,誘蛾量高于3代;5代雄成蟲(chóng)高峰期為9月上旬,誘蛾量高于4代;6代雄成蟲(chóng)高峰期為10月上旬,誘蛾量處于較低水平。成蟲(chóng)終見(jiàn)于10月下旬。
2.3 梨小食心蟲(chóng)各代雄成蟲(chóng)高峰期和誘蛾量
從表1可看出,不同生境果園梨小食心蟲(chóng)雄成蟲(chóng)各代高峰期和誘蛾量不盡相同。平地桃園與平地梨園的高峰期較為一致,僅1代和4代高峰期不一致,但與坡地桃園1~5代高峰期均不一致。平地梨園與坡地桃園僅1代高峰期一致,2~5代均不一致。且同一果園不同世代高峰期間隔時(shí)間也不一致。平地梨園越冬代誘蛾量最高,達(dá)12.2頭;1代和2代高峰期誘蛾量坡地桃園高于平地桃園和梨園,分別為5.2頭和4.3頭;3代則是平地桃園較高,達(dá)3.1頭;而4代和5代平地梨園較高,為3.4頭和6.6頭;平地桃園和平地梨園的6代高峰期誘蛾量差異不大。
3 結(jié)論與討論
本研究利用性信息素對(duì)杭州余杭不同生境桃園、梨園中梨小食心蟲(chóng)進(jìn)行監(jiān)測(cè)發(fā)現(xiàn),2019年余杭地區(qū)發(fā)生6~7代梨小食心蟲(chóng)成蟲(chóng),與沈群超等[9]和馮帆[10]報(bào)道的浙江地區(qū)梨小食心蟲(chóng)發(fā)生6代結(jié)果不一致,平地桃園和平地梨園較往年其他地區(qū)發(fā)生代數(shù)多1代。越冬代高峰期至梨小食心蟲(chóng)發(fā)生期結(jié)束共持續(xù)205 d。根據(jù)有效積溫預(yù)測(cè)法,參考杭州余杭地區(qū)2019年4月至10月平均溫度255℃以及杜娟等和李文亮等報(bào)道的梨小食心蟲(chóng)發(fā)育起點(diǎn)溫度和有效積溫[1112],分別計(jì)算得到完成6個(gè)梨小食心蟲(chóng)世代需要199.1 d和171.2 d,小于205 d,理論上在平地桃園和平地梨園可完成7代,與實(shí)際結(jié)果吻合。
梨小食心蟲(chóng)的田間發(fā)生情況受溫度、濕度、食物、天敵等多種生態(tài)因子調(diào)控[7]。其中溫度對(duì)梨小食心蟲(chóng)發(fā)育歷期影響較大,杜娟等報(bào)道在20、23、26、29、32℃等5種不同溫度條件下飼養(yǎng)梨小食心蟲(chóng)的發(fā)育歷期分別為49.72、38.99、29.78、25.81、27.18 d,各溫度下發(fā)育歷期達(dá)顯著差異(P<0.05)[11]。而徐龍祥則認(rèn)為在19~31℃范圍內(nèi),梨小食心蟲(chóng)成蟲(chóng)前期的發(fā)育歷期滿足線性關(guān)系y=-0.0133 9+0.00168 3x(R2=0.993 5),溫度越高發(fā)育歷期越短[13]。2019年杭州余杭地區(qū)梨小食心蟲(chóng)成蟲(chóng)共發(fā)生226 d,時(shí)間跨度大,各世代之間溫度差異較大,并且南北坡溫差也是不可忽略的因素,因此不同生境果園同一世代梨小食心蟲(chóng)高峰期發(fā)生時(shí)間和各世代高峰期間隔時(shí)間不盡相同。李曉軍等報(bào)道2008年至2012年,山東泰安肥城桃產(chǎn)區(qū)梨小食心蟲(chóng)各世代發(fā)生時(shí)間、高峰期和發(fā)生量均存在差異[14],說(shuō)明梨小食心蟲(chóng)在不同年份的發(fā)生情況也是不一致的。
另外,王怡等報(bào)道用不同成熟度的桃、蘋果、梨飼養(yǎng)各世代梨小食心蟲(chóng)發(fā)現(xiàn),寄主植物的種類和果實(shí)成熟度均可影響梨小食心蟲(chóng)幼蟲(chóng)齡數(shù)[15]。王沖等認(rèn)為不同成熟度果實(shí)可影響梨小食心蟲(chóng)各蟲(chóng)態(tài)的發(fā)育歷期[16]。楊小凡報(bào)道用杏等6種不同寄主植物飼養(yǎng)梨小食心蟲(chóng),除卵期外,對(duì)梨小食心蟲(chóng)各蟲(chóng)態(tài)發(fā)育歷期均有明顯影響[17]。焦旭東等報(bào)道梨小食心蟲(chóng)在李、杏、梨3種不同寄主植物上的高峰期時(shí)間和發(fā)生量存在差異[18]。不同寄主植物對(duì)梨小食心蟲(chóng)各蟲(chóng)態(tài)發(fā)育歷期和發(fā)生量有一定影響,推測(cè)本研究中不同果園梨小食心蟲(chóng)各代成蟲(chóng)高峰期發(fā)生時(shí)間和發(fā)生量不一致與寄主植物種類不同有關(guān)。
2019年杭州余杭地區(qū)梨小食心蟲(chóng)成蟲(chóng)發(fā)生周期長(zhǎng),達(dá)226 d。不同生境果園梨小食心蟲(chóng)發(fā)生情況不一致,僅依據(jù)往年的發(fā)生期和田間為害癥狀,難以準(zhǔn)確把握梨小食心蟲(chóng)發(fā)生動(dòng)態(tài),致使藥劑防治效果不佳。因此不同生境果園需獨(dú)立設(shè)置性信息素監(jiān)測(cè),測(cè)報(bào)梨小食心蟲(chóng)發(fā)生時(shí)間和高峰期,并依據(jù)杜娟等報(bào)道的基于性誘劑監(jiān)測(cè)的梨小食心蟲(chóng)數(shù)量動(dòng)態(tài)確定防治指標(biāo)[6],這對(duì)果園適期防控梨小食心蟲(chóng)有重要意義,并有助于推廣梨小食心蟲(chóng)監(jiān)測(cè)技術(shù)。
參考文獻(xiàn)
[1] 熊彩珍,王志龍,趙敏,等.桃蛀果害蟲(chóng)的發(fā)生特點(diǎn)與動(dòng)態(tài)研究[J].現(xiàn)代農(nóng)業(yè)科技,2018(8):122.
[2] 鄭錦城.梨小食心蟲(chóng)Grapholita molesta (Busck)地理種群耐熱性差異及其相關(guān)分子基礎(chǔ)[D].北京:中國(guó)農(nóng)業(yè)科學(xué)院,2015.
[3] NEVEN L G, KUMAR S, YEE W L, et al. Current and future potential risk of establishment of Grapholita molesta (Lepidoptera: Tortricidae) in Washington State [J].Environmental Entomology,2018,20(10):19.
[4] 林露菲,劉依華,黃邦侃.福建桃、梨果蛀蟲(chóng)及梨小食心蟲(chóng)初步研究[J].華東昆蟲(chóng)學(xué)報(bào),1998,7(2):3540.
[5] 康芝仙,胡全德,王樹(shù)君,等.梨小食心蟲(chóng)在吉林省高寒山區(qū)李樹(shù)上的發(fā)生與防治[J].昆蟲(chóng)知識(shí),1995,32(5):270272.
[6] 杜娟,劉彥飛,譚樹(shù)乾,等.基于性誘劑監(jiān)測(cè)的梨小食心蟲(chóng)防治指標(biāo)[J].植物保護(hù)學(xué)報(bào),2013,40(2):140144.
[7] 陳梅香,駱有慶,趙春江,等.梨小食心蟲(chóng)研究進(jìn)展[J].北方園藝,2009(8):144147.
[8] 趙玄,王宇磊,沈志杰,等.性信息素不同配方、劑量與誘捕器類型對(duì)引誘甜菜夜蛾效果的影響[J].環(huán)境昆蟲(chóng)學(xué)報(bào),2018,40(5):968973.
[9] 沈群超,吳華新,王立如,等.浙江慈溪桃園和梨園梨小食心蟲(chóng)發(fā)生規(guī)律研究[J].中國(guó)南方果樹(shù),2018,47(4):104106.
[10]馮帆.浙西北梨癭蚊、梨小食心蟲(chóng)的發(fā)生規(guī)律與物理防治技術(shù)研究[D].杭州:浙江農(nóng)林大學(xué),2014.
[11]杜娟,郭建挺,張亞素,等.溫度對(duì)梨小食心蟲(chóng)Grapholitha molesta Busck生長(zhǎng)發(fā)育及繁殖的影響[J].西北農(nóng)業(yè)學(xué)報(bào),2009,18(6):314318.
[12]李文亮,李定旭,董鈞鋒,等.梨小食心蟲(chóng)發(fā)育起點(diǎn)溫度和有效積溫研究[J].河南農(nóng)業(yè)科學(xué),2010,39(10):8082.
[13]徐龍祥.溫度變化對(duì)梨小食心蟲(chóng)生物學(xué)特性的影響[D].泰安:山東農(nóng)業(yè)大學(xué),2016.
[14]李曉軍,翟浩,王濤,等.山東泰安肥城桃產(chǎn)區(qū)梨小食心蟲(chóng)發(fā)生規(guī)律及預(yù)測(cè)預(yù)報(bào)模型研究[J].果樹(shù)學(xué)報(bào),2013,30(5):841847.
[15]王怡,孔維娜,郭永福,等.不同寄主植物對(duì)梨小食心蟲(chóng)幼蟲(chóng)齡數(shù)的影響[J].山西農(nóng)業(yè)科學(xué),2018(9):15151521.
[16]王沖,楊小凡,馮娜,等.不同成熟度的寄主果實(shí)對(duì)梨小食心蟲(chóng)生長(zhǎng)發(fā)育及繁殖的影響[J].應(yīng)用昆蟲(chóng)學(xué)報(bào),2017,54(1):100106.
[17]楊小凡.寄主因子對(duì)梨小食心蟲(chóng)的發(fā)生、生長(zhǎng)發(fā)育及其產(chǎn)卵選擇性的影響[D].保定:河北農(nóng)業(yè)大學(xué),2013.
[18]焦旭東,郭艷蘭,何彩,等.甘肅武威不同果園梨小食心蟲(chóng)發(fā)生規(guī)律及防治技術(shù)研究[J].中國(guó)果樹(shù),2017(3):6770.
(責(zé)任編輯:田 喆)
