黃土高原雨養區不同種植年限紫花苜蓿土壤真菌群落的分布特征
2021-06-28 02:50:16羅珠珠張耀全牛伊寧李玲玲蔡立群蔡雪梅劉家鶴
干旱地區農業研究 2021年3期
馬 欣,羅珠珠,張耀全,牛伊寧,李玲玲,蔡立群,蔡雪梅,劉家鶴
(1.甘肅農業大學資源與環境學院,甘肅 蘭州 730070;2.甘肅省干旱生境作物學重點實驗室,甘肅 蘭州 730070;3.延安市農業科學研究所,陜西 延安 716000)
紫花苜蓿(Medicagosativa)有著高產、耐瘠、保土等優良特性,在西部黃土高原地區廣泛種植,是當地退耕還林的主要草種[1],亦是糧改飼、發展草牧業的優質牧草,種植苜蓿可促進當地草食畜牧業健康發展[2]。苜蓿根瘤可在根瘤菌的作用下固定游離氮,地上部枯枝落葉可經微生物分解增加土壤有機碳含量,改善土壤質量[3]。土壤微生物是土壤生態系統的重要組成部分,可調節土壤養分轉化和系統的穩定性,提高其抗干擾能力[4]。一般情況下,多年生植物通過根系分泌物對土壤微生物的群落結構和分布產生影響,從而改變微生物的豐富度和多樣性[5]。紫花苜蓿屬于軸根型牧草,根系發達,多年種植紫花苜蓿會導致土壤容重減小、深層水分虧缺,土壤質量及牧草品質逐步退化[6-7],亦會對土壤微生物結構和功能產生影響。
真菌是土壤微生物中數量最多的類群之一[8],可通過參與土壤中碳素轉化、物質循環影響腐殖質的形成,穩定土壤結構和理化性質[9],是維持生態系統功能的基礎和生態系統健康的指示物[10]。Yang[11]、錢雅麗[12]等對黃土高原半干旱區不同土地利用類型真菌群落的研究發現種植紫花苜蓿有利于土壤真菌的生長和繁殖,從而提高真菌豐富度和多樣性,特別是通過影響土壤顆粒機械組成、養分含量、pH,進而影響土壤子囊菌和擔子菌等優勢菌群的分布格局[13]。研究表明,連作抑制土壤中細菌及固氮菌的生長,促進真菌群落的繁殖,導致土壤由高肥力‘細菌型’向低肥力‘真菌型’轉換[14-15]。耿德洲[16]等在寧南黃土高原的研究表明,隨著苜蓿種植年限的增加真菌群落多樣性降低,群落組成從農田逐漸向天然草地方向演變。麻冬梅[6]在寧南山區的研究表明,不同種植年限苜蓿地土壤微生物群落均以細菌為主,其豐度隨苜蓿種植年限的增加而增加,真菌的變化趨勢則不明顯,數量一直在0.10~0.14萬個·g-1范圍徘徊;邰繼承[17]對內蒙古灰色草甸土的研究發現,種植苜蓿能夠有效促進微生物群落的發展,種植5 a的紫花苜蓿土壤細菌數量是種植2 a苜蓿地細菌數量的1.86倍,但真菌數量無明顯變化。
目前,研究黃土高原雨養農業區紫花苜蓿的研究多集中在水分利用效率[18-19]、土壤理化性質[20-21]以及溫室氣體排放[22-23]等方面,對于土壤微生物群落的研究較少。而研究土壤真菌群落結構和多樣性可為農田土壤的可持續發展提供理論依據。因此,本研究以農田為對照,通過Illumina MiSeq高通量測序研究黃土高原地區不同種植年限苜蓿土壤真菌群落組成及多樣性特征,以探明不同種植年限苜蓿土壤真菌群落演變規律及優勢菌屬分布特征。研究結果在理論上有助于從分子生物學角度揭示苜蓿種植年限對土壤真菌群落特征的影響,在實踐上可為區域苜蓿適宜種植年限的選擇提供指導意義。
1 材料與方法
1.1 研究區概況
試驗設在甘肅省定西市安定區李家堡鎮的甘肅農業大學旱作農業綜合試驗站(104°44′E,35°28′N)進行,該區屬于典型的黃土高原半干旱雨養農業區,平均海拔約為2 000 m,年均太陽輻射592.9 kJ·cm-2,平均日照時數2 476.6 h,年均氣溫6.4℃,0~10℃積溫范圍為2 239.1~2 933.5℃,無霜期為141 d,年均降雨量400 mm,年蒸發量1 531 mm,干燥度2.53。土壤類型為黃綿土,土壤容重1.19 g·cm-3,pH 值8.36,土壤有機質12.01 g·kg-1,全氮0.76 g·kg-1,全磷0.79 g·kg-1。
1.2 試驗設計
試驗以玉米田為對照,2003年(L2003)、2005年(L2005)、2012年(L2012)建植的紫花苜蓿人工草地為研究對象,小區面積均為3 m×7 m=21 m2,隨機區組排列,3次重復。苜蓿品種為當地傳統品種隴東苜蓿,整個生育期無人為干預,各處理田間管理保持一致,每年刈割兩次(6月和10月)。苜蓿人工草地在2003、2005和2012年建植時施純氮105 kg·hm-2,純P2O5105 kg·hm-2,之后在苜蓿生長期間未進行施肥、灌水。玉米品種為‘先玉335’,2013年開始種植玉米,持續至今,每年播前人工撒施純N 200 kg·hm-2,純P2O5105 kg·hm-2,生育期不再追肥,收獲后期玉米根系不還田。
1.3 土樣采集
在每個采樣小區中,于2018年苜蓿頭茬花期按照五點采樣法用土鉆采集0~30 cm和30~60 cm土層土壤樣品,去除樣品中的植物根系、石塊等雜物混合均勻后過2 mm篩,裝入滅菌自封袋,干冰保存,帶回實驗室置于超低溫冰箱中用于土壤真菌群落結構和多樣性的測定。
1.4 真菌ITS高通量測序
將用于測定的土樣送至上海美吉生物醫藥科技有限公司進行真菌 ITS 高通量測序,引物如表1所示。從土壤樣品中抽提基因組DNA后,利用1%瓊脂糖凝膠電泳檢測抽提的基因組DNA,然后對樣品的ITS區進行PCR擴增(ABI GeneAmp? 9700型,Thermo Fisher Scientific,USA),將同一樣本的PCR產物混合后用2%瓊脂糖凝膠電泳檢測,使用AxyPrepDNA凝膠回收試劑盒(AXYGEN公司,USA)切膠回收PCR產物,Tris_HCl洗脫, 2%瓊脂糖電泳檢測。參照電泳初步定量結果,將PCR產物用QuantiFluorTM~ST藍色熒光定量系統(Promega公司,USA)進行檢測定量,之后按照每個樣本的測序量要求,進行相應比例的混合,隨后構建Miseq文庫,用Illumina平臺Miseq 300測序儀測序[24-25]。
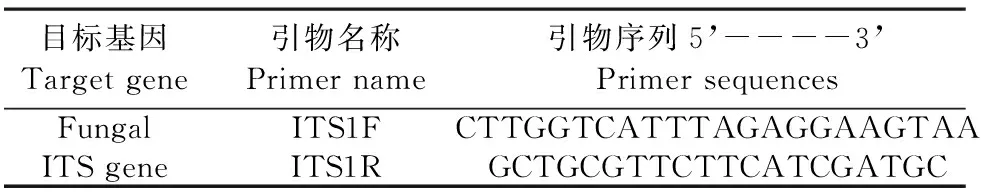
表1 目標基因的引物名稱及引物序列
1.5 Alpha多樣性指數計算方法
微生物α多樣性[26]:
式中,Chao1表示菌群豐度;Sobs為實際觀測到的OTU數;n1為只含有一條序列的OTU數目;n2為只含有兩條序列的OTU數目。
Simpson和Shannon代表微生物多樣性:
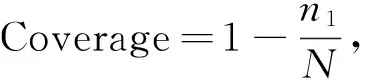
1.6 數據處理
使用Mothur指數分析,基于指數評估的OTU相似水平97%得出土壤真菌Alpha多樣性指數(Shannon和Simpson)、菌群豐富度指數(Chao1)。土壤微生物群落結構和多樣性指數均采用SPSS 22.0和Excel數據處理軟件,差異顯著性分析利用單因素方差分析(ANOVA)和多重比較法(Duncan)。維恩圖(Venn)通過I-Sanger云平臺(美吉公司,上海)進行處理,主成分分析使用Canoco 5,AI軟件進行圖表修飾。
2 結果與分析
2.1 土壤真菌群落OTU特異性分析
通過對不同種植年限土壤樣品中真菌群落Venn圖(圖1)的比較分析發現,隨種植年限增加,耕層0~30 cm苜蓿土壤真菌群落物種數在增加,L2003、L2005、L2012分別獲得OTUs 731、746、691個,農田獲得OTUs 752個,各處理特有物種分別為108、125、116和216,占各處理物種總數的14.77%、16.76%、16.79%%和28.72%;共有物種OTUs 303個,占物種總數量的10.38%。L2003和L2005共有物種數(506),高于L2003和L2012(468)、L2005和L2012(474)物種共有數。30~60 cm土層中L2003、L2005、L2012和農田分別獲得OTUs 678、675、631、697個,共有275個,占物種總數的10.26%;L2003特有OTUs 114個,L2005特有OTUs 139個,L2012特有OTUs 95個,農田特有OTUs 186個。這表明隨苜蓿種植年限延長土壤真菌物種數增加,但不同種植年限苜蓿土壤真菌特有物種數均少于農田土壤。
2.2 土壤真菌群落 Alpha多樣性
Miseq高通量測序結果優化后共獲得389693條有效序列,對所有樣品進行抽平得到38560條序列后進行后續分析。由表2看出,樣品測序覆蓋度均在0.99以上,說明測序數據量合理,基本能真實反映土壤微生物的群落組成。與農田相比,耕層0~30 cm種植苜蓿對Shannon指數無明顯影響,但其隨著苜蓿種植年限的延長顯著增加(P<0.05),表現為L2003和L2005處理顯著高于L2012,分別增加18.47%和21.34%。Chao1和Simpson指數在農田和不同種植年限苜蓿間均無顯著差異。耕層以下30~60 cm土壤剖面真菌群落的Chao1指數、Shannon指數和Simpson指數隨著種植年限的變化呈現出與耕層0~30 cm類似的趨勢,但處理間均無顯著差異。

表2 不同處理真菌Alpha多樣性指數
2.3 門水平土壤真菌群落組成特征
本研究所有供試黃綿土樣品中共檢測到真菌6門24綱73目156科313屬,土壤真菌群落門水平主要包括4個類群(圖2),占真菌總數的99%以上,分別為:子囊菌門(Ascomycota)、接合菌門(Zygomycota)、擔子菌門(Basidiomycota)和unclassified_k__Fungi。黃綿土子囊菌門相對豐度隨土層深度增加而降低,接合菌門和擔子菌門相對豐度則表現為相反趨勢。
與農田相比,耕層0~30 cm苜蓿土壤子囊菌門相對豐度顯著增加(P<0.05),且隨苜蓿種植年限延長先降低后升高,不同種植年限苜蓿土壤比農田土壤提高17.50%~32.08%。接合菌門、擔子菌門和unclassified_k__Fungi相對豐度表現為苜蓿土壤顯著低于農田土壤(P<0.05),且隨種植年限延長先升高后降低。其中接合菌門相對豐度降低62.36%~73.54%,擔子菌門豐度降低60.08%~78.84%,unclassified_k__Fungi相對豐度降低8.16%~55.05%。不同種植年限苜蓿土壤優勢門豐度亦存在差異,L2003子囊菌門相對豐度顯著高于L2005處理12.42%(P<0.05),接合菌門相對豐度顯著低于L2005處理153.54%(P<0.05)。耕層以下30~60 cm真菌優勢門群落相對豐度隨著苜蓿種植年限的變化呈現出與耕層0~30 cm類似的趨勢。農田土壤子囊菌門和擔子菌門相對豐度顯著高于苜蓿土壤(P<0.05);接合菌門相對豐度表現出農田和L2005處理顯著高于L2003和L2012處理(P<0.05);unclassified_k__Fungi相對豐度在農田和不同種植年限苜蓿土壤間無明顯差異。
2.4 屬水平土壤真菌群落組成特征
土壤真菌在屬水平種類繁雜,各個門科下屬分類地位不明確的菌屬占總序列的10.98%~32.71%,耕層0~30 cm土壤真菌相對豐度>1%的菌屬分布見圖3。農田土壤前3種優勢真菌屬分別為被孢霉屬(Mortierella,19.63%)、綠僵菌屬(Metarhizium,13.67%)和腐質霉屬(Humicola,11.63%),且表現為農田土壤顯著高于苜蓿土壤(P<0.05),其中被孢霉屬相對豐度提高62.38%~73.48%,綠僵菌屬豐度提高64.83%~85.55%,腐質霉屬豐度提高88.43%~93.53%。苜蓿土壤共有優勢真菌屬為刺孢屬(Phaeomycocentrospora, 10.49%~29.54%)、白僵菌屬(Beauveria,10.10%~13.02%)、被孢霉屬(5.21%~13.19%)和綠僵菌屬(1.98%~4.81%)。隨苜蓿種植時間延長,刺孢屬、赤霉菌屬(Gibberella)相對豐度呈先降低后升高趨勢,被孢霉屬、Tetracladium屬和隱球菌屬相對豐度呈相反趨勢先升高后降低,白僵菌屬、綠僵菌屬和Lectera屬相對豐度逐漸升高。
30~60 cm土層,農田土壤優勢屬為被孢霉屬(32.45%),相對豐度顯著高于苜蓿土壤28.11%~80.75%(P<0.05)(圖4)。刺孢屬(7.94%~17.95%)和被孢霉屬(6.25%~23.33%)為苜蓿土壤共有優勢菌屬。隨苜蓿種植時間延長,刺孢屬、被孢霉屬和Tetracladium屬相對豐度呈先升高后降低趨勢,白僵菌屬、綠僵菌屬、土赤殼屬和叢赤殼屬(Nectria)相對豐度呈先降低后升高趨勢,而鐮刀菌屬相對豐度顯著增加。統計分析表明,L2012刺孢屬相對豐度顯著低于L2003和L2005(P<0.05),降低45.95%和555.77%;L2012處理白僵菌屬和綠僵菌屬相對豐度顯著高于其余處理(P<0.05),增加84.73%~98.28%和51.22%~92.82%;L2003鐮刀菌屬相對豐度顯著高于L2005和L2012處理54.00%和64.04%(P<0.05)。
2.5 土壤真菌群落結構的主成分分析
主成分分析(圖5)表明,不同種植年限苜蓿土壤真菌屬在PC1軸和PC2軸上解釋度分別為66.90%和17.13%,農田和苜蓿草地土壤真菌群落顯著分開。農田土壤真菌群落主要分布在第一、四象限,苜蓿土壤真菌群落主要分布在第二、三象限。載荷因子反映真菌屬水平豐度與所提取主成分之間的相關性,載荷因子絕對值越高,表示該真菌屬對主成分的影響越大[27]。表3結果表明,叢赤殼屬、土赤殼屬和鐮刀菌屬與第一主成分(PC1)具有較高的相關性,Lectera菌屬、支頂孢屬和赤霉菌屬與第二主成分具有較高相關性,而這6類菌屬均屬子囊菌門肉座菌目類植物病原菌。與第一、二主成分相關的菌屬均與植物病害相關,說明長期種植苜蓿引起相關病原菌累積。
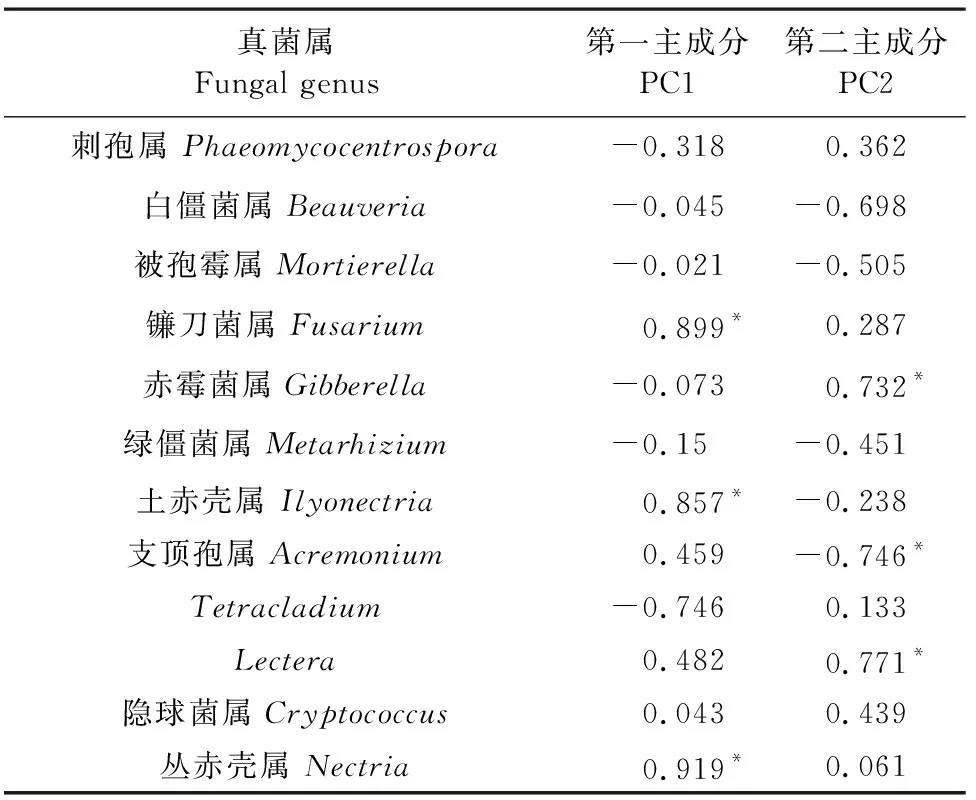
表3 真菌屬水平豐度的主成分載荷因子Table 3 Component loading factor of soil fungalgenera in relative abundance
3 討 論
3.1 苜蓿種植對黃綿土真菌多樣性的影響
土壤微生物群落結構組成及多樣性的增加能夠改變土壤微生態的穩定性,其在一定程度上可反映土壤健康狀況[28]。真菌參與土壤有機物質分解,是土壤生態系統中不可或缺的一部分[29]。趙彤等[30]對寧南山區不同植被類型土壤微生物群落進行比較時發現,苜蓿人工草地能夠顯著改善土壤微生物多樣性,這主要是因為不同作物根系分泌物存在異質性,釋放根系分泌物(氨基酸、分泌酶、粘液和細胞裂解物)會影響微生物多樣性,且凋落物經分解后釋放的有機物質成分和數量亦使土壤微生物群落結構組成和多樣性發生不同程度的變化[31-32]。已有研究表明,氮含量添加會降低真菌豐度及多樣性并改變真菌的群落結構;相對細菌來說,真菌有更強的吸收氮素和磷素的能力[33-35]。本研究發現,氮含量添加的農田土壤真菌群落多樣性Chao1、Simpson和Shannon指數與苜蓿土壤并無明顯差異,這說明黃綿土區不同作物根系分泌物引起的土壤微生物多樣性的差異可能主要存在于細菌群落而非真菌群落,本試驗后續還需進一步探索。但是本研究發現,不同種植年限的紫花苜蓿草地土壤真菌多樣性發生了改變,隨著苜蓿種植年限延長,真菌群落的多樣性(Shannon指數)顯著增加,之后隨著苜蓿生長的衰敗又有所降低。這與耿德洲[16]在寧南山區研究的結果基本一致,但是最高值出現的年份有所不同,這可能與當地苜蓿適宜種植的年限有關,隴中黃土高原苜蓿適宜種植的年限可達10 a[36],遠遠長于寧南山區苜蓿6 a的適宜種植年限。多年生植物通過分泌根系分泌物對土壤微生物的群落結構和分布產生影響,進而改變微生物的豐富度和多樣性[5]。苜蓿為多年生草種,根系發達,根系分布可直接改變真菌群落結構多樣性[37];且隨著苜蓿種植時間的延長到達旺盛期后地表覆蓋物不斷增加,凋落物和根系的腐解程度增大,苜蓿根瘤菌的固氮作用增強,使土壤有機碳和養分含量增加,根系通過養分轉化和能量傳遞等過程提高了真菌豐富度[16,38-39]。隨著苜蓿草地慢慢退化,苜蓿自身較強的化感物質通過根系分泌物、植株殘體分解和地上部分淋洗等形式被釋放到土壤中,并呈現逐年富集的趨勢[40],這種自毒效應損害了根系的生長發育并降低根瘤菌固氮能力[41],引起土壤養分含量和微生物豐度與多樣性下降。
3.2 苜蓿種植對黃綿土真菌群落分布特征的影響
農田土壤真菌群落組成與苜蓿存在差異,說明作物類型、耕作和施肥等措施對土壤中微生物群落組成有著顯著影響。本研究發現農田和苜蓿土壤真菌均以子囊菌門為主,占比達到了69.15%~91.45%,其次為接合菌門和擔子菌門,這與肖禮和Wang[42-43]等在黃土高原地區土壤真菌的研究結果相似,進一步說明子囊菌門為黃綿土真菌優勢菌門,但由于受不同種植作物影響,不同生境中其相對豐度具有一定差異[42-43]。子囊菌門以腐生菌為主,可將土壤中的有機物質分解成易被作物吸收的養分,土壤中難以分解的纖維素和木質素主要依賴于子囊菌[44]。本研究中苜蓿土壤子囊菌門相對豐度顯著高于農田土壤,農田土壤雖然通過長期耕作提高了土壤輕組C和N含量,但連續種植苜蓿增加了土壤腐殖質含量,且苜蓿能與根瘤菌形成共生關系固氮,所以苜蓿能為子囊菌的生長提供良好的繁殖條件[45-46]。農田土壤擔子菌門相對豐度顯著高于苜蓿土壤,可能是因為傳統耕作措施種植玉米影響作物殘茬分解過程,使土壤中木質素含量升高,增加了擔子菌門相對豐度,而苜蓿土壤長期種植不翻耕,分解速率降低,導致擔子菌門豐度較低[47]。本研究耕層土壤接合菌門相對豐度隨著苜蓿種植年限的延長呈先升高后降低趨勢,長期種植苜蓿且未翻耕,植物殘體落葉等進入土壤轉化為腐殖質,使有機物質含量提高,為微生物的生長和繁殖提供了充足的底物[48],而接合菌門恰恰能很好地利用腐生環境[49]。苜蓿種植15 a以后地上生物量降低,養分含量降低與生長衰退同時發生[50],有機物質的減少導致接合菌門相對豐度隨之減小。
本研究中被孢霉屬相對豐度表現為農田土壤顯著高于苜蓿土壤,隨苜蓿種植年限延長被孢霉屬相對豐度先增加后降低。被孢霉屬能促進植株生長,是土壤中的有益微生物,可補充氮素、溶磷,提高植物的抗病性[51-52];健康植株土壤中的相對豐度顯著高于發病植株土壤[53]。玉米根系分泌糖和氨基酸,能促進微生物活化難利用的磷[54],進而提高了被孢霉屬豐度。隨苜蓿種植時間延長,土壤養分和地上生物量均逐漸增加,為被孢霉屬的生長繁殖提供了充足的底物,而持續種植紫花苜蓿會導致土壤產生干層,當苜蓿種植年限達到一定的階段,土壤干燥化嚴重,此時土壤孔隙度降低、空氣含量下降,水分條件和氧氣含量成為了限制被孢霉屬生長繁殖的限制性因素,缺乏生存條件的被孢霉屬豐度又逐漸下降[7,45,50]。本研究農田土壤綠僵菌屬相對豐度顯著高于苜蓿土壤,且隨苜蓿種植年限延長逐漸下降。綠僵菌具有較強的耐高溫和耐旱特性,豐富的N、P可以促進菌絲的生長并能增加其菌苔厚度[55]。農田土壤由于連年施肥使得速效N、P含量較高,且覆膜使土壤溫度持續上升,提高了氮素轉化速率,增加了綠僵菌屬豐度;L2012處理的苜蓿處于生長旺盛期,土壤中可以被利用的營養物質多,能夠為綠僵菌的生長與繁殖提供適宜的環境[56]。隨著苜蓿種植年限的延長,土壤速效磷含量大幅降低[57],導致綠僵菌的豐度持續降低。本研究中農田土壤腐質霉屬相對豐度均顯著高于苜蓿土壤,可能是因為腐質霉屬營腐生,可在土壤內或病株殘體上越冬,農田玉米收獲后根系殘留在地表過冬,碳含量充足,加之覆膜保濕,為腐質霉屬的繁殖保證了水分和營養條件。
主成分分析發現,土壤鐮刀菌屬、叢赤殼屬和土赤殼屬均與第一主成分具有較高的相關性,鐮刀菌屬是重要的植物病原菌之一,對種子萌發、根系、根莖生長產生不利影響[58]。本研究苜蓿土壤鐮刀菌屬相對豐度隨種植時間延長隨之增長,由于苜蓿隨種植時間的增加可通過根系分泌物和植株殘體分解等形式釋放化感物質到土壤中,產生自毒效應,并呈現出逐年富集的趨勢[40],特別是對其根系的生長發育及根瘤菌的固氮能力造成損害[41],使病原真菌含量逐漸增長。叢赤殼屬和土赤殼屬作為子囊菌門肉座菌目的分生孢子在草本植物的根際大量存在,為腐生真菌,常引起植物根腐病等病害[59-60],本試驗中與第一主成分相關性較高的菌屬均為與植物病害有關的病原菌,這與苜蓿種植年限有一定的關系。
4 結 論
與農田相比,種植苜蓿對土壤真菌多樣性無明顯影響,但其表現為隨苜蓿種植年限的延長顯著增加,之后隨著苜蓿生長的衰敗有所降低。不同種植年限苜蓿地土壤真菌群落結構組成相似性高,但其優勢菌群分布受種植時間的影響。黃土高原雨養農業區農田和苜蓿土壤真菌優勢門為子囊菌門(56.46%~91.45%)、接合菌門(5.23%~32.83%)和擔子菌門(1.92%~9.06%);農田土壤已分類優勢菌屬為被孢霉屬(19.63%~32.45%),苜蓿土壤優勢菌屬為刺孢屬(7.94%~29.54%)、被孢霉屬(5.21%~23.33%)和白僵菌屬(3.13%~29.73),且表現為隨著種植年限延長,耕層土壤促生菌(白僵菌屬和綠僵菌屬)豐度下降,部分病原菌(鐮刀菌屬、赤霉菌屬)豐度增加。
