一種非洲豬瘟病毒假病毒細胞感染模型的建立及應用
2021-06-16 10:40:32唐嘉怡蔡艷星王欣玲姜世勃陸路
微生物與感染 2021年5期
唐嘉怡,蔡艷星,王欣玲,姜世勃,陸路
復旦大學基礎醫學院教育部/衛健委/醫科院醫學分子病毒學重點實驗室,上海 200032
非洲豬瘟病毒(African swine fever, ASFV)是一種具有包膜結構的核質大DNA病毒(nucleocytoplasmic large DNA virus, NCLDV),隸屬于非洲豬瘟病毒科(Asfarviridae),是該病毒家族的唯一成員,也是迄今為止發現僅有的蟲媒DNA病毒[1]。ASFV可以通過直接接觸、糞-口途徑或者是天然宿主蜱蟲在家豬或野豬之間快速傳播[2-3],部分家豬感染后會出現出血熱等癥狀,甚至引發急性死亡[4],此病原體引發的疾病被稱為非洲豬瘟(African swine fever, ASF)。該動物疾病傳染性極強,家豬或野豬感染烈性毒株后的死亡率接近100%[5-6],因此被世界動物衛生組織(World Organization for Animal Health, OIE)列為法定報告動物疫病(www.oie.int/),也是我國重點防范的動物疫情之一(http://www.moa.gov.cn/)。
2018年,中國出現了第一例非洲豬瘟病毒感染家豬的病例[7]。由于缺乏疫苗和抗病毒藥物,能夠控制疫情的方式主要是對疫區進行封鎖,對疫區內的生豬進行撲殺[8]。此次的非洲豬瘟疫情對我國的國民生活質量、食品安全以及農業產業經濟發展都造成了較大的危害。因此,開展對ASFV的感染機制、疫苗和抗病毒藥物等研究具有重要意義。
ASFV能夠在被污染的豬肉以及富含營養的環境中長久存在,具有極強的傳播性[9],因此在我國對于ASFV活病毒的研究主要在生物安全三級(biosafety level 3,BSL-3)或四級(biosafety level 4,BSL-4)實驗室中進行(http://www.moa.gov.cn/)。這在一定程度上限制了對于ASFV的研究,包括中和抗體等病毒入侵抑制劑的篩選。對于需要在BSL-3或BSL-4實驗室中進行活病毒研究的病原體,如H7N9亞型禽流感病毒、尼帕病毒和中東呼吸綜合征冠狀病毒等,均已建立了成熟的假病毒(pseudovirus)細胞感染模型[10-12]。使用假病毒細胞感染模型可在生物安全等級較低的實驗室中開展一些研究工作[13],包括病毒的進入機制、中和抗體評價以及入侵抑制劑的活性檢測等[14-17]。而目前對于ASFV假病毒細胞感染模型的相關研究極少。
假病毒是一種重組的病毒顆粒,其核心骨架和包膜蛋白來源于不同的病毒[18]。通常會對假病毒的核心骨架基因進行突變或修飾,這種改變使得假病毒無法產生包膜或表面蛋白,須加入另外的可表達病毒包膜蛋白或表面蛋白(可介導病毒進入靶細胞的功能蛋白)的質粒,從而產生只可單輪感染的假病毒[19]。
目前常用的假病毒包裝體系主要包括:I型人類免疫缺陷病毒(human immunodeficiency virus, HIV-1)包裝體系、猴免疫缺陷病毒(simian immunodeficiency virus, SIV)包裝體系和水皰口炎病毒(vesicular stomatitis virus, VSV)包裝體系等[20]。假病毒包裝系統中的核心骨架通常帶有報告基因,如熒光素酶報告基因或熒光蛋白基因,假病毒感染細胞的程度與所帶的報告基因表達成正比,因此相較于對真病毒(authentic virus),假病毒的感染定量分析更加簡單、易于操作[21]。基于上述原理,假病毒細胞感染模型被廣泛用于病毒的細胞嗜性[22]、進入機制、受體的發現以及中和抗體等入侵抑制劑的評價等研究。
為了能夠包裝獲得可高效感染的ASFV假病毒細胞感染模型,本研究選取多種ASFV的包膜蛋白,包括p12、CD2v、p30、p54和pE248R,用于假病毒的包裝及感染模型的建立。基于這5種包膜蛋白在病毒感染過程中的關鍵作用,構建表達5種包膜蛋白的真核表達質粒,應用較為廣泛和高效的HIV-1包裝體系和VSV假病毒包裝體系嘗試制備多種ASFV假病毒。通過假病毒感染細胞能力的檢測,在VSV包裝體系上獲得1種感染能力較強的ASFV假病毒。以此為基礎,建立能夠穩定感染部分敏感細胞的ASFV假病毒細胞感染模型,并且用已報道[23]的能夠在ASFV進入階段發揮抗病毒作用的芫花素(Genkwanin)驗證此假病毒細胞感染模型的可靠性。該模型有望用于通過檢測所攜帶熒光基因的表達程度篩選藥物對ASFV假病毒感染的抑制活性,為在安全級別要求較低的實驗室中開展對ASFV生命周期中進入階段的研究以及病毒入侵抑制劑研發提供候選模型。
1 材料與方法
1.1 實驗材料
1.1.1 細胞系HEK 293T細胞(人源胚胎腎細胞)、Huh7細胞(人肝癌細胞系)、MDCK細胞(犬腎臟細胞系)均購買于中國科學院細胞庫,Vero細胞(非洲綠猴腎細胞)和BHK21細胞(倉鼠腎細胞)購于American type culture collection(ATCC),RD細胞(人橫紋肌瘤細胞)由上海市公共衛生臨床中心提供。以上細胞均使用含10%(體積分數)胎牛血清(fetal bovine serum, FBS)的DMEM培養基(Dulbecco’s modified eagle medium)培養。
1.1.2 質粒真核表達非洲豬瘟包膜蛋白p30、CD2v、pE248R、p54、p12的質粒pcDNA3.1-p30、pcDNA3.1-CD2v、pcDNA3.1-pE248R、pcDNA3.1-p54、pcDNA3.1-p12購買于南京金斯瑞生物科技有限公司(所選取包膜蛋白來源毒株African swine fever virus isolate Pig/HLJ/2018,GENBANK NO. MK333180),質粒pNL4-3.Luc.R-E-[貨號pNL4-3-Luc-R-E-(HIV-Luc)]購于ATCC,質粒pVKD-NA和pVKDA-HA由上海市金山公共衛生臨床中心提供。
1.1.3 菌株與假病毒Top10感受態細胞由本實驗室保存,Stable 3感受態細胞購自北京全式金生物技術有限公司,VSV-ΔG rLuc假病毒顆粒由Professor Benhur Lee惠贈。
1.2 實驗方法
1.2.1 HIV-1包裝體系制備非洲豬瘟病毒假病毒HIV-1包裝體系常用于制備包膜病毒的假病毒。參考流感病毒、冠狀病毒等假病毒的制備方式[10,12],在六孔板中使用含10%(體積分數)FBS的DMEM培養293T(3×105個/孔)細胞,待細胞生長密度達到60%時進行轉染。使用不同的包膜蛋白質粒和pNL4-3.Luc.R-E-骨架質粒共同轉染細胞,8 h后更換DMEM,細胞在37 ℃,5%(體積分數) CO2條件下繼續培養48 h。收集上清液,并使用0.45 μm過濾膜(購買于Millipore)過濾,將假病毒分裝于1.5 mL EP管中,-80 ℃條件保存。
1.2.2 VSV包裝體系制備非洲豬瘟病毒假病毒參考尼帕病毒的假病毒制備方式[24-25],利用VSV體系包裝ASFV假病毒。在六孔板中接種293T細胞(3×105個/孔),使用10%(體積分數)FBS的DMEM過夜培養。使用不同包膜蛋白質粒轉染細胞,繼續培養6 h。加入含有VSV包膜蛋白G蛋白的VSV-△G rLuc假病毒感染細胞,2 h后使用磷酸鹽緩沖液(phosphate buffered saline,PBS)潤洗細胞3次,棄去PBS加入10% FBS的DMEM培養基,繼續培養48 h。收集假病毒后,-80 ℃保藏。
1.2.3 熒光素酶檢測(luciferase assay)檢測假病毒感染水平制備假病毒所使用的HIV-1骨架質粒pNL4-3.Luc.R-E-含有螢火蟲熒光素酶報告基因,VSV-△G rLuc假病毒中含有海腎熒光素酶報告基因。感染細胞后,使用熒光素酶檢測試劑盒(luciferase assay system,購自美國Promega)或海腎熒光素酶檢測試劑盒(Renilla luciferase assay system,購自美國Promega)評價假病毒的感染水平。提前12~16 h在96孔板中培養Vero細胞或BHK21細胞(1×104個/孔),將保藏于-80 ℃冰箱中ASFV假病毒取出融化,震蕩混勻。向細胞中加入100 μL假病毒液,37 ℃培養12 h。棄去上清液,每孔加入含體積分數1% FBS的DMEM培養基,繼續培養48 h。根據熒光素酶檢測試劑盒的說明書要求,檢測相對發光單位(relative light unit,RLU)值。
1.2.4 ASFV假病毒抑制實驗藥物對于ASFV假病毒抑制活性的檢測參考某些假病毒的抑制實驗[17]。在96孔板上過夜培養Vero細胞(1×104個/孔)備用,將藥物進行倍比稀釋,并加入等體積的ASFV假病毒,37 ℃條件下共同孵育30 min。將藥物與假病毒的混合液加入細胞中,將該孔記為藥物孔,繼續培養12 h。同時,將只加入等體積假病毒的孔,記為病毒孔;只含有細胞的孔記為細胞孔。棄去所有上清液,更換為含1% (體積分數)FBS的DMEM培養基,繼續在37 ℃,5%(體積分數)CO2條件下培養48 h。利用luciferase assay檢測病毒感染情況(RLU值),并計算藥物的抑制率。計算公式:藥物抑制率RDI(%)=(RLU病毒孔-RLU藥物孔)/(RLU病毒孔-RLU細胞孔)×100%。在GraphPad Prism 6.0中通過不同濃度下藥物抑制率生成函數曲線,并計算藥物半數抑制濃度(half maximal inhibitory concentration, IC50)。
1.2.5 統計學方法所有結果均進行3次重復獨立實驗驗證。數據分析使用GraphPad Prism 6.0,雙總體t檢驗,P<0.05表明差異具有統計學意義。
2 結果
2.1 HIV-1包裝體系制備ASFV假病毒
為了制備ASFV假病毒,本研究選取應用較為成熟和廣泛的HIV-1包裝體系,進行嘗試。假病毒的制備方法,參考中東呼吸綜合征冠狀病毒(Middle East respiratory syndrome coronavirus, MERS-CoV)的假病毒包裝方式[12]。首先,嘗試使用不同的包膜蛋白質粒pcDNA3.1-p30、pcDNA3.1-CD2v、pcDNA3.1-pE248R、pcDNA3.1-p54以及pcDNA3.1-p12分別與HIV骨架質粒共同轉染293T細胞,在已報道的可以被ASFV活病毒感染的Vero細胞上檢測假病毒的感染能力。如圖1A所示,使用不同包膜蛋白質粒制備的ASFV假病毒與細胞本底的熒光強度接近,而作為陽性對照的H7N9假病毒能夠高效地感染靶細胞,表明所使用的HIV-1包裝體系穩定,所制備的ASFV假病毒在所測條件下未表現顯著的感染能力。提示使用單個包膜蛋白質粒和HIV-1包裝體系較難制備出能夠高效感染的ASFV假病毒。
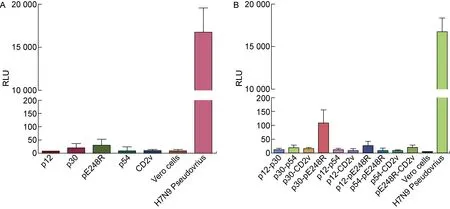
A: The HIV-1 packaging system with single plasmid encoding envelope protein for generating ASFV pseudovirus. To produce ASFV pseudovirus, single plasmid encoding different envelope proteins was transferred into 293T cells with pNL4-3.Luc.R-E-. The RLU corresponding to the infectivity of ASFV pseudovirus in Vero cells was measured. B: The HIV-1 packaging system with double plasmids encoding envelope protein for generating ASFV pseudovirus. To produce ASFV pseudovirus, dual plasmids encoding different envelope proteins was transferred into 293T cells with pNL4-3.Luc.R-E-. The RLU corresponding to the infectivity of ASFV pseudovirus in vero cells was measured.
隨后,參考H7N9亞型禽流感病毒(Avian Influenza virus, AIV)的假病毒制備方式,嘗試使用2種包膜蛋白質粒的不同組合與HIV-1骨架質粒共同轉染293T細胞,從而產生ASFV假病毒。如圖1B所示,使用表達p30和pE248R包膜蛋白的質粒所制備的ASFV假病毒的熒光強度雖略高于其余質粒組合所制備的假病毒,但在統計學上并沒有顯著差異,并且感染水平遠低于H7N9假病毒(RLU>10 000)。現有的假病毒細胞感染模型中,RLU值大多能達到103以上[12,26],故使用以上方法制備的ASFV假病毒滴度較低,用于細胞感染或病毒抑制實驗等可能出現不穩定的情況。然而,這個結果提示ASFV的包膜蛋白p30以及pE248R可能在ASFV感染Vero細胞過程中發揮著一定的作用。
2.2 VSV包裝體系制備ASFV假病毒
VSV包裝體系是最常見的假病毒體系之一,如尼帕病毒(Nipah virus, NiV)、埃博拉病毒(Ebola virus, EBOV)等可以在VSV包裝體系上制備假病毒[11,27]。本研究參考NiV的假病毒包裝方式[24-25],分別使用以上5種ASFV包膜蛋白質粒制備假病毒。含有VSV包膜蛋白G蛋白的VSV-△G rLuc感染細胞后,可能會摻入收獲的假病毒中,在檢測所制備的ASFV假病毒熒光強度時可能出現假陽性。因此,在轉染細胞時設置轉染空載質粒(pcDNA3.1)的陰性對照。如圖2A所示,使用p30、pE248R和p54包膜蛋白分別轉染細胞所獲得的ASFV假病毒熒光強度略高于陰性對照,但與該背景RLU值差距不足3倍。在以前的研究中,使用VSV假病毒包裝體系制備不同病毒的假病毒時,對照組與產生的假病毒RLU值相差均大于3倍[28],因此使用這3種包膜蛋白質粒單獨包裝的ASFV假病毒也難以感染細胞。此外,p12和CD2v兩種包膜蛋白質粒分別制備的ASFV假病毒的熒光強度與陰性對照接近,可能是殘留的VSV-△G rLuc假病毒所造成的假陽性。隨后,選取5種包膜蛋白質粒中的2種進行組合,共同轉染293T細胞以獲得ASFV假病毒。如圖2B所示,其中利用p30-p54、p30-pE248R和p54-pE248R這3種組合制備的假病毒感染Vero細胞后的RLU值均達到104以上。其中,p30和pE248R包膜蛋白質粒組合包裝的假病毒在Vero細胞上的感染水平高于其他2組(P<0.001),表明利用此種方法可能獲得更高效感染細胞的ASFV假病毒,可在此假病毒的基礎上建立較為高效的ASFV細胞感染模型。隨后,基于p30和pE248R質粒組合進行優化, 通過增加不同的包膜蛋白質粒共同轉染細胞,嘗試獲得更高滴度的ASFV假病毒。如圖2 C所示,使用3種包膜蛋白質粒制備的ASFV假病毒,滴度反而低于原先制備的ASFV假病毒(P<0.01或P<0.000 1)。以上結果顯示,使用VSV包裝體系以及p30、pE248R包膜蛋白質粒的組合制備方法,所包裝出的假病毒可能具有較優的感染活性,適合用于建立細胞感染模型。
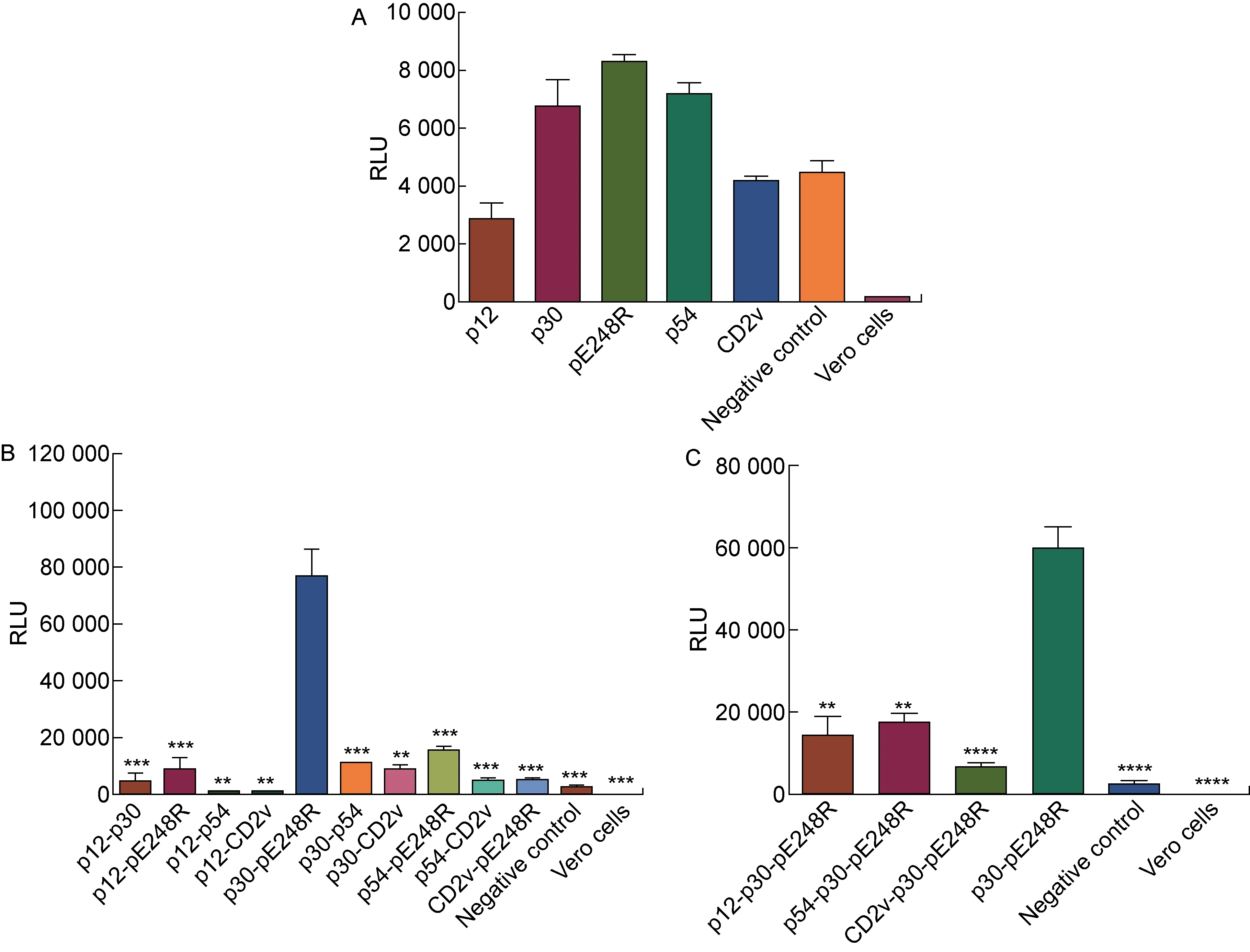
A: The VSV packaging system with single plasmid encoding envelope protein for generating ASFV pseudovirus. To produce ASFV pseudovirus, single plasmid encoding different envelope proteins was transferred into 293T cells, followed by the infection of VSV-G rLuc. The RLU corresponding to the infectivity of ASFV pseudovirus in Vero cells was detected. B: The VSV packaging system with double plasmids encoding envelope protein for generating ASFV pseudovirus. To produce ASFV pseudovirus, dual plasmids encoding different envelope proteins was transferred into 293T cells, followed by the infection of VSV-G rLuc. The RLU corresponding to the infectivity of ASFV pseudovirus in Vero cells was detected. C: The VSV packaging system with triple plasmids encoding envelope protein for generating ASFV pseudovirus. To produce ASFV pseudovirus, triple plasmids encoding different envelope proteins was transferred into 293T cells, followed by the infection of VSV-G rLuc. The RLU corresponding to the infectivity of ASFV pseudovirus in Vero cells was detected.
2.3 驗證包膜蛋白在所制備ASFV假病毒上的表達
為驗證所制備的ASFV假病毒是否能夠表達包膜蛋白,選擇其中1個包膜蛋白為代表,使用western blot(WB)檢測其在假病毒上的表達情況。構建編碼pE248R蛋白全長基因和His標簽的質粒pcDNA3.1-pE248R-His。使用該質粒替代原有的pE248R包膜蛋白質粒,和p30包膜蛋白質粒共同轉染細胞,在VSV包裝體系上制備帶有His標簽的ASFV假病毒,并進行WB檢測。如圖3所示,使用抗His標簽的抗體,在生成的ASFV假病毒中出現了對應pE248R蛋白的清晰條帶。相反,在作為陰性對照的VSV-G 假病毒中未檢測到pE248R蛋白的表達。這些結果提示,ASFV的包膜蛋白pE248R被有效整合到VSV-ΔG rLuc顆粒中,并包裝出ASFV假病毒。
2.4 ASFV假病毒細胞感染模型的特異性
為進一步驗證,所制備的ASFV假病毒(p30-pE248R-ASFV-PsV)可用于細胞感染模型的建立。在不同的細胞系上檢測了該假病毒的感染水平,其中包括已報道[29-32]能被ASFV感染的細胞系:Vero細胞和BHK21細胞。如圖4A所示, 在VSV包裝體系上使用p30和pE248R包膜蛋白質粒所制備的p30-pE248R-ASFV-PsV能夠感染已報道的非洲豬瘟敏感細胞系Vero細胞和BHK21細胞,且不感染RD細胞,具有一定的感染特異性(P<0.01或P<0.001)。如圖4B所示,作為陽性對照的VSV-G假病毒能夠感染所有用于檢測的細胞系。此外,只轉染了pcDNA3.1空載的陰性對照在感染不同細胞系后只有微弱的熒光素酶表達(見圖4C),排除了多余的VSV-G假病毒對實驗結果造成的影響。
MW: Relative molecular weight.
2.5 ASFV假病毒細胞感染模型用于藥物抗病毒活性的評價
為了進一步驗證該假病毒細胞感染模型是否可用于篩選具有抗ASFV活性的病毒入侵抑制劑,選擇已報道[23]的陽性藥物——芫花素(Genkwanin,該藥物能夠在非洲豬瘟活病毒感染Vero細胞的早期發揮抑制作用),檢測其對所建立的ASFV假病毒(p30-pE248R-ASFV-PsV)的抑制活性。結果如圖5所示,芫花素能夠濃度依賴性地抑制ASFV假病毒感染Vero細胞,其半數抑制濃度(half maximal inhibitory concentration, IC50)為4.05±0.88 μmol/L。設置VSV-G假病毒作為陰性對照,芫花素無法抑制VSV-G假病毒感染Vero細胞,也反映出該ASFV假病毒細胞感染模型在ASFV入侵抑制劑活性評價和篩選中的可行性。
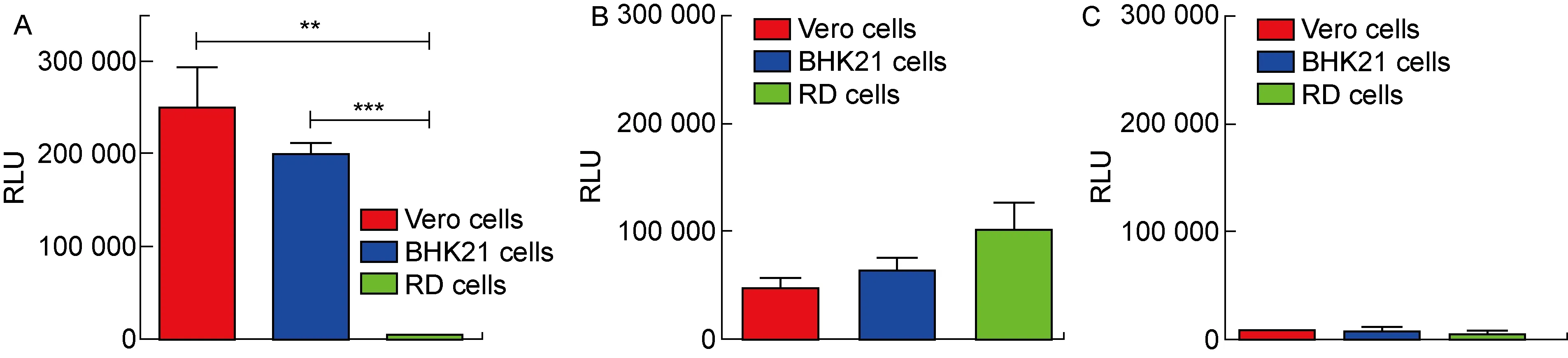
A: The ability of p30-pE248R-ASFV-PsV to infect different cell lines were determined. B: The ability of VSV-G pseudovirus to infect different cell lines. Different cell lines were infected by VSV-G as the positive control. C: The ability of negative control to infect different cell lines. Different cell lines were infected by the negative control which were generated by pcDNA3.1 and VSV packaging system. The difference between the ability of p30-pE248R-ASFV-PsV and other pseudoviruses to infect different cell lines was statistically analyzed by two-tailed unpaired Student’s t-test using GraphPad Prism, version 6.0. *** and ** mean P<0.001 and P<0.01, respectively. Values with P<0.05 was considered statistically significant.
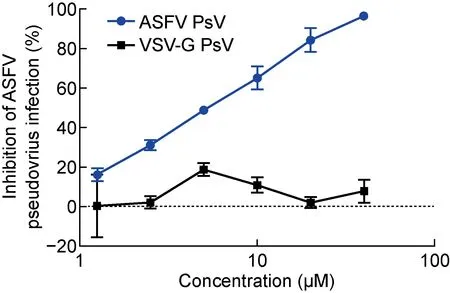
圖5 芫花素對ASFV假病毒感染的抑制活性檢測
3 討論
自2018年亞洲地區暴發非洲豬瘟疫情,我國持續有疫情感染的病例報道(1)非洲豬瘟疫情通報在中華人民共和國農業農村部官網http://www.moa.gov.cn/。。迄今為止,全球沒有獲批的非洲豬瘟疫苗上市;我國也未有針對ASFV的預防或治療藥物獲得批準。當前,從事ASFV活病毒的研究有嚴格的生物安全要求,須在生物安全三級及以上實驗室中進行。因此,疫苗和藥物的開發在一定程度上受到了限制。為了獲得可在低安全級別實驗室開展ASFV入侵機制和入侵抑制劑等藥物研究的模型,本研究選取多種ASFV的包膜蛋白,包括 p12、CD2v、p30、p54和pE248R,用于假病毒的包裝及感染模型的建立。這些蛋白在ASFV進入細胞的過程中發揮重要作用,例如位于外包膜上的p12蛋白能夠與靶細胞Vero細胞的細胞膜結合[33-34],而同樣位于外包膜上的CD2v蛋白能夠與紅細胞上的CD受體結合,從而完成病毒的吸附過程,并且增強ASFV在細胞間感染[35]。在研發非洲豬瘟疫苗的過程中發現,位于內包膜上的p30和p54蛋白能夠刺激機體產生體液免疫和細胞免疫,具有良好的抗原性[36-37]。同時,有文獻報道這兩種蛋白能夠與豬肺泡巨噬細胞結合,從而介導病毒的內化,因此推測p30和p54蛋白對于ASFV的感染可能是必須的[38]。ASFV使用內吞途徑進入細胞,隨后需要在內吞泡中完成病毒膜與內吞泡膜的融合,從而介導病毒的遺傳物質釋放于胞漿中,開啟復制過程[39]。近期一項研究證明,位于內包膜上的pE248R是介導病毒內包膜與內吞泡膜融合的關鍵因子,pE248R缺失的重組病毒無法完成感染過程以及啟動后續的基因組復制[40]。基于這5種包膜蛋白在病毒感染過程中的關鍵作用,本研究構建表達了包膜蛋白的真核表達質粒。利用ASFV的包膜蛋白質粒pcDNA3.1-p30、pcDNA3.1-pE248R和VSV包裝體系成功制備ASFV假病毒,并初步建立細胞感染模型,此細胞感染模型或可進行病毒入侵抑制劑的篩選和活性評價。
本研究在嘗試制備假病毒的過程中發現,使用HIV-1包裝體系較難產生具有高感染力的ASFV假病毒。此前嘗試制備漢坦病毒假病毒的研究中也出現類似現象:使用VSV包裝體系能夠產生較難通過HIV-1體系制備的漢坦病毒假病毒[41]。另外,在制備拉沙病毒假病毒的實驗中也發現,使用VSV包裝體系所產生的假病毒滴度高于HIV-1包裝體系[42]。因此,推測本研究使用的VSV包裝體系的包裝效率高于HIV-1包裝體系,這可能與HIV-1體系對包膜蛋白有較嚴格的選擇性有關[20]。不過,VSV體系包裝出的假病毒也不能認為可用于完全模擬活病毒的感染過程。有文獻報道,骨架病毒的形狀可能會影響構建相對應假病毒的適用性[41,43],VSV顆粒呈子彈狀,與ASFV并不相似,假病毒糖蛋白的構象和分布可能無法模擬野生型活病毒的天然狀態。因此,本研究中所制備的ASFV假病毒和細胞感染模型也可能具有局限性。與ASFV的其他包膜蛋白質粒相比,包膜蛋白質粒p30和pE248R可包裝出滴度較高的ASFV假病毒。既有研究已證明p30蛋白是ASFV內化所必需的[38],而pE248R蛋白介導病毒膜與內吞泡膜的融合[39]。因此,本文推測病毒的內化和膜融合對于ASFV的感染十分關鍵,而p30和pE248R蛋白在其中扮演著重要的角色,這也提示在對ASFV進入機制研究時應關注這兩個蛋白。
同時也發現,本研究所制備的假病毒對不同細胞系的感染能力存在差異。ASFV假病毒(p30-pE248R-ASFV-PsV)感染后,在Vero細胞上的熒光強度高于BHK21細胞。由于目前尚不清楚ASFV的細胞受體,無法確定該假病毒對于不同細胞系感染差異的原因。若能夠深入探究2種細胞系之間的差異,或許能夠為ASFV細胞受體的解析提供新思路。但是,本研究中所獲得的ASFV假病毒是否可對原代細胞產生有效感染,以及其感染特征是否與ASFV活病毒相似,這些受限于倫理及BSL-3實驗室等未開展研究。今后應做進一步研究,以明確該假病毒模型的局限性。
本研究對所建立的ASFV假病毒細胞感染模型,還檢測了芫花素的抑制活性,其IC50為4.05±0.88 μmol/L,這與文獻報道的芫花素對非洲豬瘟活病毒感染Vero細胞的抑制活性(IC50為2.9 μmol/L)相似[23]。這提示該ASFV假病毒細胞感染模型用于病毒入侵抑制劑抗ASFV活性的評價具有一定的可行性。但是否可用于抗ASFV藥物的高通量篩選,以及與活病毒感染模型相比是否具有較高的一致性,還需要進一步的研究。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中國外匯(2019年17期)2019-11-16 09:31:14
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
現代企業(2015年1期)2015-02-28 18:43:18
新高考·高一物理(2014年1期)2014-09-18 01:26:07
中國火炬(2010年7期)2010-07-25 10:26:09
