硝氮對狐尾藻附著生物膜細菌群落的影響
2021-06-09 07:32:50張松賀張子秋
河海大學學報(自然科學版) 2021年3期
張松賀,袁 強,陳 潔,張子秋
(河海大學環境學院,江蘇 南京 210098)

濕地作為一種經濟實用、環保有效的措施廣泛應用于污水處理中。水生植物吸收及微生物反硝化是濕地去除硝氮的主要途徑[2]。與挺水和浮葉植物相比,沉水植物幾乎全株生長在水面下,可直接從水體中積累養分[3]。生物膜是一種由細菌、真菌、藻類以及其他非生物成分組成的復雜微孔生物反應器[4],沉水植物可為其附著生物膜的生長提供合適的附著面以及穩定的有機碳源[2]。生物膜中的微生物在硝化反硝化等營養循環中起著關鍵作用[5]。然而,關于硝氮負荷下沉水植物附著微生物群落結構及相關反硝化微生物的響應機制仍不清楚。
本文以針狀葉、根系不發達的沉水植物狐尾藻(MyriophyllumVerticillatum)為對象,研究不同硝氮濃度條件下狐尾藻葉片及其附著微生物的形態特征變化,利用高通量測序和定量PCR分析技術探討微生物群落結構和反硝化基因豐度的變化規律,系統分析生物膜在硝態氮轉化中的潛在作用,以期為沉水植物生態修復以及水環境保護提供參考。
1 材料與方法
1.1 試驗方法
狐尾藻從南京高淳水生植物基地購買。試驗裝置使用140 L(700 mm×500 mm×400 mm)的水箱,添加7 cm混合均勻的沉積物(取自南京烏龍潭湖)以及30 cm的自來水。穩定一周后,每個水箱種植60株株高約30 cm且形態相近的狐尾藻(種植密度約300株/m2,種植深度為5 cm)。待植物馴化2周后,分別控制水體硝氮濃度為2 mg/L(M2)、8 mg/L(M8)、20 mg/L(M20)和40 mg/L(M40),并設置空白對照組M0。每2 d監測1次硝氮濃度,添加硝酸鹽化合物(w(NaNO3)∶w(KNO3)=1∶1)以維持設定濃度。每組試驗設置3個平行樣,于河海大學淺水湖泊綜合治理與資源開發教育部重點試驗室溫室內進行。
1.2 樣品測定
1.2.1 水質及理化指標測定
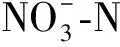
1.2.2 掃描電鏡
試驗第14天,分別采集M0和M20組的狐尾藻葉片。將植物切片后(5 mm×5 mm),依次使用30%、50%、70%、80%、90%濃度的酒精脫水15 min,再用100%濃度的酒精脫水2次,每次15 min,冷凍干燥12 h后備用。最終使用日立S-4800場發射掃描電鏡進行觀察。
1.2.3 生物膜采集
在水面下10~20 cm處采集約50 g的狐尾藻葉片,采集過程中注意避免沉積物的污染。將采集后的樣品轉移到含有500 ml磷酸鹽緩沖液(PBS,pH為7.4)的聚乙烯瓶中,超聲處理(150 W,40 kHz)1 min,225 r/min轉速下振蕩30 min后,通過50 μm的篩子去除植物碎屑,最后離心10 min(8 000 rpm)。離心后的生物膜保存在-80 ℃用于提取DNA。
1.2.4 PCR擴增
使用特定引物[2],利用定量聚合酶鏈反應(qPCR)方法測定生物膜DNA樣品中反硝化基因的豐度。采用熒光染料SYBR-Green法,使用MyiQ2實時PCR檢測系統(Bio-Rad)在20 μL反應混合物中進行qPCR擴增,反應物包括supermix(10 μL)、上下游引物(0.5 μL)、DNA模板(1 μL)和超純水(8 μL)。qPCR擴增共包括40個循環。
1.3 數據分析
硝氮去除率及反硝化基因豐度使用Origin 8.0處理制圖;利用SPSS(2000版)進行Pearson相關性指數分析和單因素方差分析;使用PAST軟件(Version 3.10)基于Bray-Curtis距離在OTU水平上對細菌群落結構聚類分析;βSOR(S?rensen差異性指數)、βSIM和βNES使用R語言(Version 3.5.3)中的betapate軟件包計算;生物膜門水平上的細菌群落分布使用R語言中的cirlize軟件包分析;利用Heml軟件繪制術細菌屬水平熱圖;利用CANOCO(Version 4.5, USA)進行冗余分析;網絡分析使用R語言中的psych包,基于Spearman相關系數(|r|>0.6,P<0.05)分析后,使用Gephi軟件可視化。
2 結果與討論
2.1 不同處理組對硝氮去除率及水體理化指標的變化
本研究監測了試驗14 d內狐尾藻對硝酸鹽的去除效果(圖1),結果表明:各處理組前3 d對NO3-N的去除率高達92.11%~94.17%,但隨著時間的累積均顯著下降,在試驗末期低于40%。初期階段氮的去除可能與狐尾藻對硝氮的吸收吸附作用有關[6]。處理3 d后,M2對硝氮的去除率在13.09%~43.88%之間,但在同一取樣時間點都始終大于其他3組的去除效果(2.96%~27.38%)。已有研究表明,沉水植物在過量氮負荷下會吸收比它們營養所需的更多氮,但這種額外的吸收會隨著時間推移接近飽和而減少[7]。Li等[8]研究表明,暴露于硝氮濃度為10 mg/L的水體14 d后,沉水植物苦草的生理活性受到顯著抑制。這些結果說明高硝氮可能對狐尾藻產生了脅迫。

圖1 狐尾藻對硝酸鹽去除率的變化Fig.1 Changes of nitrate removal rate by Myriophyllum Verticillatum
試驗14 d內監測了各處理組水體理化參數,如表1所示。電導率Ec隨硝酸鹽濃度升高而升高。與對照M0相比,M2-M40處理組水體的pH、DO和ORP的均值均下降。高濃度硝氮負荷下水體DO和pH的降低與狐尾藻受抑制的光合系統有關,植物減少了對二氧化碳吸收及氧氣釋放[8];另外,生物膜形成過程中微生物的繁殖也會加大水體耗氧[9]。

表1 試驗14 d內對照(M0)及硝酸鹽處理組(M2-M40)水體的理化特征
2.2 狐尾藻葉片及表面附著生物膜形態變化
根據電鏡圖像,對照處理下狐尾藻葉片結構清晰可見,附著了少量微生物,存在有典型的桿菌群落(圖2(a));而20 mg/L硝氮處理14 d后,狐尾藻表面附著了大量的細菌(球菌、芽孢桿菌和其他細菌)、微生物聚集團和藻類等微生物,這些微生物通過自身分泌的物質粘連聚集在一起,形成了明顯的生物膜群落結構[10](圖2(b))。這表明硝氮施加刺激了沉水植物附著生物膜的增長[9],過量附著微生物會影響植物對光的接收,進而抑制沉水植物生長。

圖2 狐尾藻葉片表面掃描電鏡照片Fig.2 SEM photographs of Myriophyllum Verticillatum leaves surface
2.3 不同處理組狐尾藻附著生物膜細菌群落特征
根據高通量測序結果,對細菌多樣性指數進行了分析(表2)。與對照M0相比,M2-M40的細菌物種豐富度(OTU數量)、群落多樣性(shannon和simpson指數)和群落均勻度(simpson均勻度)均顯著降低(ANOVA,P<0.05)。這些結果說明硝酸鹽的富集顯著降低了狐尾藻附著生物膜細菌群落的生物多樣性。與本研究結果類似,Zhang等[2]發現水體中的硝氮負荷降低了金魚藻附生生物膜的微生物多樣性。

表2 生物膜樣品細菌測序α多樣性指數
聚類分析將5個生物膜樣品分為2組:硝氮處理樣品M2-M40聚集為組Ⅰ,而對照M0單獨在組Ⅱ內(圖3(a)),這表明硝氮的施加顯著改變了狐尾藻附著生物膜的細菌群落結構。βSOR多樣性可用來描述微生物群落間的差異程度。根據物種的存在與否,βSOR可將生物多樣性過程歸結于兩種生物現象:βNES(生物因死亡或遷移造成的物種損失)和βSIM(物種更替)[11]。研究中βSOR多樣性結果表明,硝酸鹽脅迫下細菌群落多樣性構建過程中(βSOR= 0.60),物種更替現象(βSIM= 0.44)大于物種損失(βNES= 0.16)。

圖3 細菌群落OTU水平聚類和主要門/變形菌綱相對豐度Fig.3 OTU level cluster analysis of bacterial community, and relative abundance of major phyla/Proteobacteria
在5個附著生物膜樣品中共有12個細菌門占優勢(至少在一個生物膜樣品中排前10)(圖3(b))。Proteobacteria(53.57%~71.92%)是最主要的門,其次是Planctomycetes(9.60%~20.1%)、Cyanobacteria(1.86%~15.12%)、Bacteroidetes(2.37%~5.13%)、Actinobacteria(3.66%~4.80%)、Acidobacteria(1.22%~3.74%)、Firmicutes(0.28%~1.34%)、Verrucomicrobia(1.12%~2.76%)、Chloroflexi(0.04%~1.69%)和Nitrospirae(0.03%~0.70%)等。在Proteobacteria內,5個生物膜樣品內均檢測到α-proteobacteria(35.94%~53.77%)、β-proteobacteria(11.06%~15.54%)、γ-proteobacteria(3.58%~4.75%)和δ-proteobacteria(0.25%~1.89%)(圖3(b))。
與氨氮的施加相比[4],硝酸鹽負荷下沉水植物附著生物膜中Nitrospirae和Chloroflexi的相對豐度較低,而Planctomycetes的相對豐度較高。α-proteobacteria和β-proteobacteria在異養反硝化反應器中對硝酸鹽的去除起著重要作用[12],其在M2、M8、M20、M40樣品中的豐度也顯著高于M0(P<0.05)。Planctomycetes含碳水化合物水解基因[2],Bacteroidetes和Actinobacteria能消化植物纖維、水解有機物[13]。這些結果表明硝氮可刺激與硝酸鹽轉化相關細菌的生長。藍細菌門Cyanobacteria光合能力較強,是河湖系統中常見的自養細菌[14],其豐度在對照M0中較高。
2.4 優勢屬及反硝化基因的變化
5個生物膜樣品中共監測出23個優勢屬(至少在一個生物膜樣品中排前10)(圖4(a))。其中包含7個反硝化細菌屬:Rhodobacter(7.53%~27.68%)、Hyphomicrobium(1.09%~5.42%)、Agrobacterium(0.38%~2.59%)、Flavobacterium(0.66%~1.66%)、Acinetobacter(0%~1.06%)、Pseudomonas(0.08%~0.91%)和Bacillus(0.08%~0.31%)[2]。反硝化是自然生態系統中氮循環的重要一環。在本研究中,Rhodobacter是生物膜中最主要的反硝化細菌。Rhodobacter、Acinetobacter和Bacillus的豐度在M2、M8、M20、M40樣品中均高于M0。Rhodobacter在異養條件下可去除大量硝態氮[15];Pang等[16]發現Rhodobacter和Acinetobacter也是狐尾藻附著生物膜的優勢反硝化細菌;Bacillus是污水處理廠和濕地脫氮中常見的異養反硝化菌[17]。這些結果表明硝氮刺激了上述反硝化細菌的增長,在濕地微生物反硝化過程中發揮了重要作用。
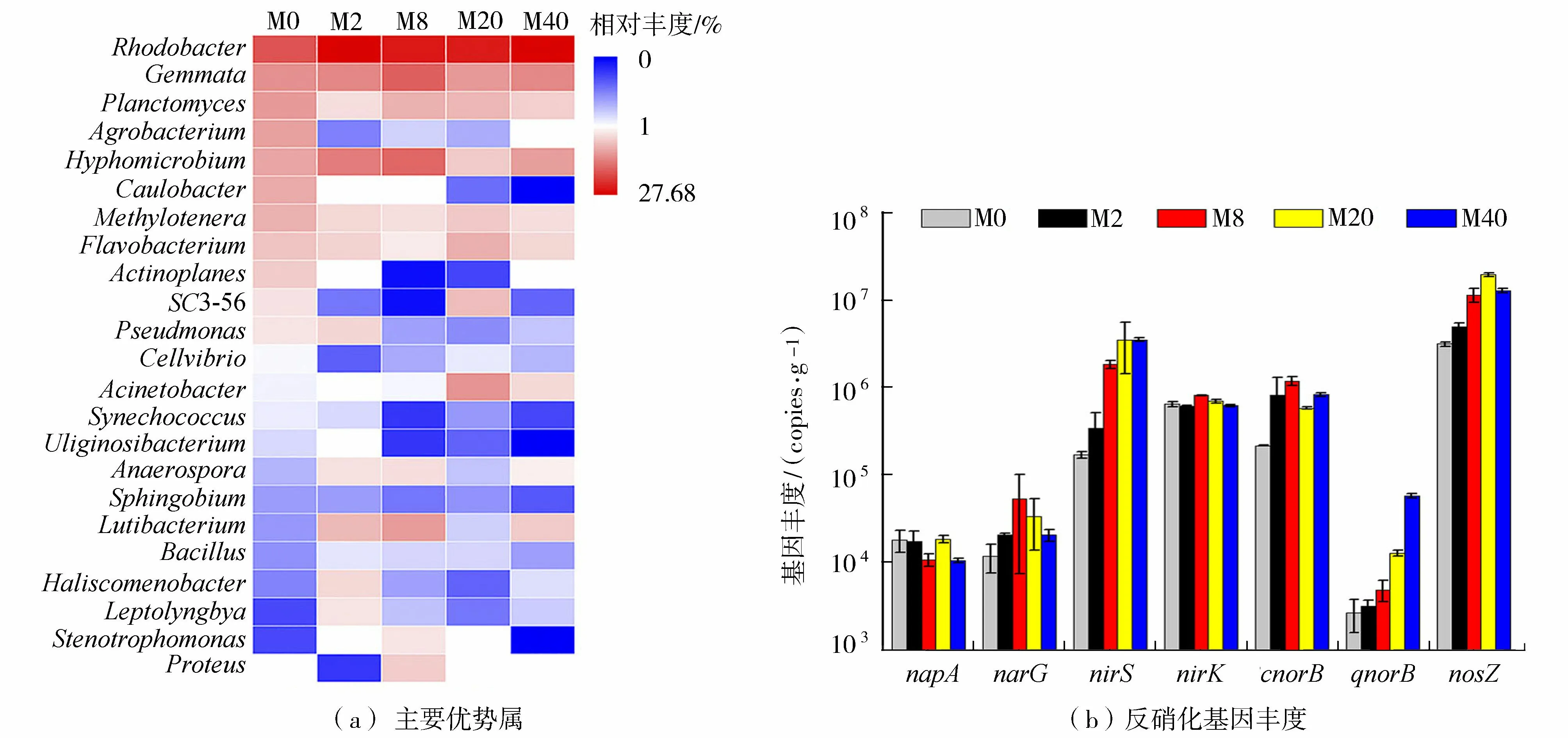
圖4 5個生物膜樣品中主要優勢屬及反硝化基因豐度Fig.4 Dominant genera and abundance of denitrification genes in five biofilm samples

2.5 優勢屬和反硝化基因對環境參數的響應
采用RDA方法描述了環境參數與生物膜內優勢屬、反硝化基因間的潛在關系(圖5),結果顯示反硝化細菌(Rhodobacter、Acinetobacter和Bacillus)及反硝化基因(nirS、cnorB、qnorB和nosZ)與硝氮濃度呈正相關,這與Yan等[3]的研究結果總體一致。基因napA需在完全好氧條件下表達及發揮功能[17],好氧反硝化細菌Hyphomicrobium和Pseudomonas[2]及基因napA均與DO呈正相關。pH也是影響反硝化的重要指標,已有研究表明pH值較低的環境會抑制反硝化細菌Agrobacterium[20]和Flavobacterium[21]的增長。在本研究中這兩種反硝化菌的豐度也均與pH呈正相關。這些結果說明硝氮負荷下,硝氮濃度、DO和pH是影響濕地反硝化微生物群落的主要環境因素。

圖5 優勢屬和反硝化基因對環境參數響應的冗余分析(RDA)Fig.5 Redundancy analysis (RDA) of the response of dominant genera and denitrification genes to environmental parameters
2.6 優勢屬和反硝化基因的網絡分析
生物膜微生物間存在復雜的相互作用[2],網絡共現性分析常用于描述系統內微生物間的相互作用。本研究對23個優勢屬及7個反硝化基間的相互關系進行了分析(圖6),其中每個節點代表一個屬/基因,每條邊代表其所連接屬/基因間的相關性(紅色為正相關,藍色為負相關)。對無任何顯著相關性的節點剔除后,共得到25個節點(20個細菌屬,5個反硝化基因),74條具有顯著相關性邊(52條邊呈正相關,22條邊呈負相關)。參與有機物降解菌Anaerospora[22]和Lutibacterium[23]分別具有8條和7條相關關系邊,是本研究生物膜網絡結構的關鍵節點,在營養循環及維持生物膜結構穩定方面起著關鍵作用[22]。

圖6 優勢屬及反硝化基因之間相互關系網絡Fig.6 Relationship network diagram among dominant genera and denitrification genes
3 結 語
沉水植物狐尾藻對濕地水體中硝態氮的初次去除具有顯著效果(92.11%~94.17%),而后去除效率逐步降低。硝酸鹽負荷對生物膜的生長具有促進作用,但降低了附著生物膜細菌多樣性。βSOR多樣性指數表明硝酸鹽脅迫下,細菌群落多樣性構建過程中物種更替現象(βSIM=0.44)大于物種損失(βNES=0.16);硝酸鹽負荷對附著生物膜的細菌微生物群落結構具有顯著影響,且刺激了反硝化細菌(Rhodobacter、Acinetobacter和Bacillu)的生長及反硝化基因(napG、nirS、cnorB、qnorB和nosZ)豐度的增加;生物膜微生物間存在復雜的相互作用,參與有機物質降解菌屬Anaerospora和Lutibacterium是生物膜網絡結構的關鍵節點。
