粒徑對(duì)平菇栽培用玉米芯發(fā)酵料代謝物的影響
2021-06-06 06:59:37劉芹胡素娟孔維麗崔筱袁瑞奇孔維威邱立友
中國(guó)瓜菜 2021年3期
劉芹 胡素娟 孔維麗 崔筱 袁瑞奇 孔維威 邱立友


摘 要:為了解玉米芯粒徑對(duì)平菇栽培用發(fā)酵料中代謝物的影響,采用代謝組學(xué)技術(shù)分析添加不同粒徑玉米芯的發(fā)酵料中微生物代謝物及其代謝通路。結(jié)果表明,添加小粒徑玉米芯(D50 = 0.5 cm)和大粒徑玉米芯(D50 = 1.5 cm)的發(fā)酵料中微生物代謝物差異顯著。在正離子(POS)和負(fù)離子(NEG)模式下分別篩選得到464種和201種差異代謝物,包括芳香族化合物、氨基酸、糖及醇類(lèi)、脂質(zhì)、生物堿等。差異代謝物分別富集到90條(POS模式)和94條(NEG模式)代謝通路,差異顯著的有2條,分別為源自鳥(niǎo)氨酸、賴(lài)氨酸、煙酸生物合成生物堿和組氨酸代謝。說(shuō)明不同粒徑玉米芯發(fā)酵料中微生物代謝物具有顯著差異,為發(fā)酵料栽培平菇原料選擇提供了理論依據(jù)。
關(guān)鍵詞:平菇;發(fā)酵料;玉米芯;代謝組;差異代謝物
Abstract: Metabolomics technique was used to analyze the differences of microbial metabolites in fermentation materials for Pleurotus ostreatus cultivation adding corncobs with different particle sizes. Results showed a significant difference between microbial metabolites in fermentation materials adding small particle (D50 = 0.5 cm) and large particle (D50 = 1.5 cm) of corncobs. 464 and 201 differential metabolites were screened at POS and NEG modes, respectively, including aromatic compounds, amino acids, sugars and alcohols, lipids, alkaloids. The analysis of KEGG metabolic pathway showed that differential metabolites were enriched in 90 (POS mode) and 94 (NEG mode) metabolic pathways, respectively, and two of which were significantly different. They were Biosynthesis of alkaloids derived from ornithine, lysine and nicotinic acid and Histidine metabolism. The results indicated that there were significant differences between the microbial metabolites in the fermentation materials adding corncobs with different particle sizes, which provided important information for the selection of cultivation materials and the improvement of cultivation techniques of P. ostreatus.
Key words: Pleurotus ostreatus; Fermentation material; Corncob; Metabolomics; Differential metabolite
平菇(Pleurotus ostreatus)是世界上栽培最廣泛的食用菌之一[1],由于其豐富的營(yíng)養(yǎng)(如蛋白質(zhì)、礦物質(zhì)和維生素),顯著的抗氧化、抗病毒、提高免疫力等生物活性而受到消費(fèi)者的青睞[2]。平菇易于栽培,可以在各種農(nóng)業(yè)副產(chǎn)物(玉米芯、秸稈、麩皮等)上生長(zhǎng),其年產(chǎn)量在世界范圍內(nèi)持續(xù)快速增長(zhǎng)[3]。目前平菇的栽培技術(shù)主要分為3種:生料栽培、熟料栽培和發(fā)酵料栽培[4-5]。與生料栽培和熟料栽培相比,發(fā)酵料栽培因其低污染、低成本、工藝簡(jiǎn)單、經(jīng)濟(jì)效益高等優(yōu)點(diǎn)而在世界范圍內(nèi)廣泛應(yīng)用[4]。平菇栽培用發(fā)酵料是以秸稈、玉米芯等農(nóng)業(yè)副產(chǎn)物為主要原料,加入少量的麩皮、石灰等,在微生物的參與下,經(jīng)短期、好氧發(fā)酵而制備的[1]。玉米是我國(guó)三大糧食作物之一,玉米芯約占玉米產(chǎn)量的21%,是一種產(chǎn)量巨大的農(nóng)副產(chǎn)品,來(lái)源廣泛、價(jià)廉易得[6-7]。目前,玉米芯除用于制備糠醛、木糖醇等外,很大一部分被作為農(nóng)業(yè)廢棄物直接燃燒,造成資源浪費(fèi)和環(huán)境污染。玉米芯組織均勻、硬度適宜、吸水性強(qiáng),將其堆制發(fā)酵后用于平菇栽培,不僅有助于平菇產(chǎn)量和品質(zhì)的提高,而且可以實(shí)現(xiàn)農(nóng)業(yè)副產(chǎn)物的高效生物轉(zhuǎn)化[7]。
微生物在發(fā)酵料制備過(guò)程中起著重要的作用,微生物群落動(dòng)態(tài)變化影響有機(jī)物的降解,而有機(jī)物的降解決定了發(fā)酵的成熟度[8]。微生物能將培養(yǎng)料中復(fù)雜的有機(jī)質(zhì)(纖維素、半纖維素、木質(zhì)素等)進(jìn)行降解,使其變成簡(jiǎn)單、易于利用的小分子物質(zhì),從而為平菇生長(zhǎng)提供營(yíng)養(yǎng)物質(zhì);有些特定微生物如Actinobacteria、Thermus和Bacillus還可以產(chǎn)生抗菌、殺蟲(chóng)的物質(zhì),為平菇生長(zhǎng)創(chuàng)造一個(gè)適宜的環(huán)境[9]。河南省農(nóng)業(yè)科學(xué)院植物營(yíng)養(yǎng)與資源環(huán)境研究所食用菌創(chuàng)新團(tuán)隊(duì)前期研究發(fā)現(xiàn),不同粒徑的玉米芯持水率、孔隙度等物理性質(zhì)的不同導(dǎo)致發(fā)酵料升溫開(kāi)始時(shí)間、高料溫保持時(shí)間、發(fā)酵周期及菌袋污染率等顯著不同,這些指標(biāo)影響了發(fā)酵料質(zhì)量。發(fā)酵料質(zhì)量與其中微生物的活動(dòng)和代謝產(chǎn)物有關(guān),目前還未有不同粒徑玉米芯發(fā)酵料制備過(guò)程中微生物代謝差異的研究報(bào)道。
同時(shí)采用200次響應(yīng)的置換檢驗(yàn)來(lái)驗(yàn)證模型的過(guò)擬合現(xiàn)象,避免采用OPLS-DA方法進(jìn)行分析時(shí)出現(xiàn)過(guò)擬合。圖3為OPLS-DA模型的置換檢驗(yàn)結(jié)果。Q2的回歸直線與y軸的交點(diǎn)在負(fù)半軸;POS模式R2Y(cum)(0.0,0.84),Q2(cum)(0.0,-0.84);NEG模式R2Y(cum)(0.0,0.73),Q2(cum)(0.0,-1.02)。這表明建立的OPLS‐DA模型是穩(wěn)健可靠的,不存在過(guò)擬合現(xiàn)象。因此,可以用于分析不同粒徑玉米芯發(fā)酵料中代謝物的差異性。
2.2 差異化合物的篩選與分析
以p<0.05和VIP>1為條件篩選出具有差異性表達(dá)的化合物,在POS模式下篩選鑒定得到464個(gè)差異代謝物,其中上調(diào)代謝物為214種,下調(diào)代謝物為250種(圖4-a);在NEG模式經(jīng)過(guò)篩選鑒定得到201個(gè)差異代謝物,其中上調(diào)代謝物為111種,下調(diào)代謝物為90種(圖4-b)。
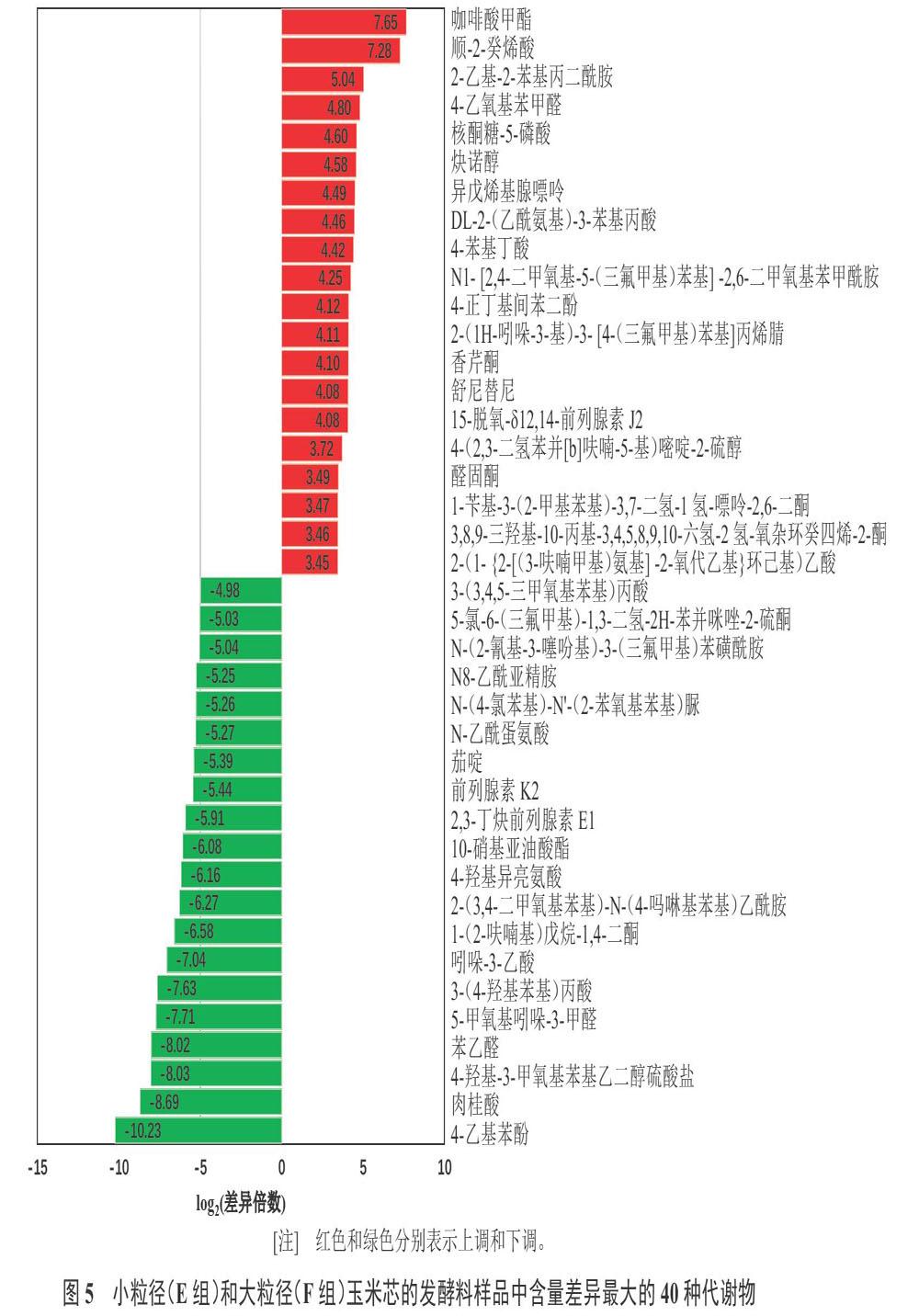
對(duì)篩選到的差異代謝物進(jìn)行l(wèi)og2處理,變化最顯著的40種(上調(diào)和下調(diào))差異代謝物見(jiàn)圖5。與小粒徑玉米芯發(fā)酵料相比,大粒徑玉米芯發(fā)酵料中咖啡酸甲酯(Methyl caffeate)、順-2-癸烯酸(cis-2-Decenoic acid)、2-乙基-2-苯基丙二酰胺(2-Ethyl-2-phenylmalonamide)、4-乙氧基苯甲醛(4-Ethoxybenzaldehyde)、核酮糖-5-磷酸(Ribulose-5-phosphate)、炔諾醇(Nirvanol)、異戊烯基腺嘌呤(Isopentenyladenine)等物質(zhì)的含量顯著增加,而1-(2-呋喃基)戊烷-1,4-二酮(1-(2-Furyl)pentane-1,4-dione)、吲哚-3-乙酸(Indole-3-acetic acid)、3-(4-羥基苯基)丙酸(3-(4-Hydroxyphenyl) propionic acid)、5-甲氧基吲哚-3-甲醛(5-Methoxyindole-3-carbaldehyde)、苯乙醛(Phenylacetaldehyde)、4-羥基-3-甲氧基苯基乙二醇硫酸鹽(4-Hydroxy-3-methoxyphenylglycol sulfate)、氫化肉桂酸(Hydrocinnamic acid)、4-乙基苯酚(4-Ethylphenol)等物質(zhì)含量顯著降低。
2.3 差異代謝物的代謝通路富集分析
將不同粒徑玉米芯發(fā)酵料中的差異代謝物導(dǎo)入KEGG(Kyoto Encyclopedia of Genes and Genomes)數(shù)據(jù)庫(kù)進(jìn)行代謝通路富集分析,共計(jì)富集到90(POS模式)和94條(NEG模式)代謝通路。其中,富集差異代謝物較多的為組氨酸代謝(Histidine metabolism)、源自鳥(niǎo)氨酸和賴(lài)氨酸及煙酸生物合成生物堿(Biosynthesis of alkaloids derived from ornithine, lysine and nicotinic acid)、煙酸和煙酰胺代謝(Nicotinate and nicotinamide metabolism)、苯丙素類(lèi)的生物合成(Biosynthesis of phenylpropanoids)、磷酸轉(zhuǎn)移酶系統(tǒng)(Phosphotransferase system)、芳香族化合物的降解(Degradation of aromatic compounds)等(圖6)。按p值從小到大的順序進(jìn)行了排序,共找到顯著性富集的KEGG路徑2條,分別為源自鳥(niǎo)氨酸、賴(lài)氨酸和煙酸生物合成生物堿(p=0.005 9),主要包括哌啶酸(Pipecolic acid)、莨菪堿(Tropine)、野靛堿(Cytisine)、煙酸(Nicotinic acid)、L-天冬氨酸(L-Aspartic acid)共5種代謝物;組氨酸代謝(p=0.019 8),主要包括咪唑丙烯酸(Urocanic acid)、L-組氨酸(L-Histidine)、L-組氨醇(L-Histidinol)、麥角硫因(Ergothioneine)、甲基咪唑乙酸(Methyl imidazole acetic acid)、1-甲基組氨酸(1-Methylhistidine)、咪唑乙酸(Imidazole acetic acid)、L-天冬氨酸(L-Aspartic acid)共8種代謝物(表1)。
3 討論與結(jié)論
采用非靶向代謝組學(xué)技術(shù),以不同粒徑玉米芯發(fā)酵料為材料,對(duì)其中的微生物代謝產(chǎn)物進(jìn)行檢測(cè)和分析。在POS和NEG模式下分別篩選和鑒定出464種和201種差異代謝物。這些差異代謝物是影響發(fā)酵質(zhì)量的主要因素[9]。差異代謝物涉及的代謝通路有芳香化合物降解、苯丙素類(lèi)的生物合成、組氨酸代謝、酪氨酸代謝等。組氨酸是含有異吡唑環(huán)的氨基酸,是機(jī)體蛋白質(zhì)的構(gòu)成氨基酸,也是一些功能蛋白質(zhì)(如血紅蛋白)的主要組成氨基酸,在代謝中起重要作用[16]。酪氨酸是蛋白質(zhì)代謝的標(biāo)志物,在適當(dāng)?shù)臈l件下,某些特定的細(xì)菌能夠從苯丙氨酸合成L-酪氨酸[17]。與大粒徑玉米芯發(fā)酵料相比,小粒徑玉米芯發(fā)酵料中酪氨酸代謝和組氨酸代謝中涉及的大部分差異代謝物含量較高,表明小粒徑玉米芯發(fā)酵料中微生物酪氨酸代謝和組氨酸代謝能力較強(qiáng)。賈玲[18]研究發(fā)現(xiàn)玉米芯中纖維素、木質(zhì)素和半纖維素的質(zhì)量分?jǐn)?shù)分別為41.87%、24.33%和36.93%。在自然界中,木質(zhì)纖維素可以被微生物分解成低分子的芳香族化合物,如苯環(huán)之間用C-C鍵連接的聯(lián)苯化合物、C-O-C鍵連接的芳醚、二芳基丙烷及一些單體芳香族化合物如水楊酸、阿魏酸等[19]。這些低分子化合物被運(yùn)輸?shù)桨麅?nèi)經(jīng)過(guò)一系列外圍分解途徑和中心代謝途徑生成三羧酸循環(huán)中間體,進(jìn)入三羧酸(TCA)循環(huán),為細(xì)胞供能[19]。Actinobacteria、Thermus和Bacillus屬的微生物具有降解木質(zhì)纖維素和產(chǎn)生抑菌物質(zhì)的能力,從而為后續(xù)雙孢蘑菇生長(zhǎng)創(chuàng)造一個(gè)適宜環(huán)境[9]。本研究結(jié)果表明,玉米芯發(fā)酵料也含有豐富的木質(zhì)纖維素降解產(chǎn)物和抑菌物質(zhì)。其中,小粒徑玉米芯發(fā)酵料中木質(zhì)纖維素代謝產(chǎn)物如原兒茶酸、水楊酸、4-氯苯酚、松柏醇等含量較高。此外,小粒徑玉米芯中還有較高含量的抑菌物質(zhì)如金雀花堿(Cytisine)、毛菜堿(Salsolinol)、柚皮素(Naringenin)、白藜蘆醇(Resveratrol)、苯乙醛等。而大粒徑玉米芯發(fā)酵料中芐星青霉素(Benzathine)含量較高,這意味著大粒徑玉米芯發(fā)酵料中含有較多的青霉屬雜菌,這也與實(shí)際平菇栽培中小粒徑玉米芯發(fā)酵料的染菌率顯著低于大粒徑玉米芯發(fā)酵料的結(jié)論相一致[20]。小粒徑玉米芯發(fā)酵料中吲哚乙酸(Indole-3-acetic acid)含量也較高,它是一種刺激植物生長(zhǎng)的激素類(lèi)物質(zhì),對(duì)平菇菌絲生長(zhǎng)也具有較強(qiáng)的促進(jìn)作用[21]。Xiang等[22]研究發(fā)現(xiàn),Alcaligenes、Pseudomonas屬的一些細(xì)菌可以產(chǎn)生生長(zhǎng)素,這可能是發(fā)酵料浸提液能夠促進(jìn)平菇菌絲生長(zhǎng)的原因之一[23]。
綜上所述,小粒徑玉米芯發(fā)酵料質(zhì)量?jī)?yōu)于大粒徑玉米芯發(fā)酵料,更適宜種植平菇,該結(jié)果為發(fā)酵料栽培平菇原料的選擇提供了理論依據(jù)。本研究主要是針對(duì)發(fā)酵料制備開(kāi)展的代謝組學(xué)研究,而對(duì)栽培過(guò)程中不同粒徑玉米芯發(fā)酵料對(duì)平菇生長(zhǎng)發(fā)育的影響還需深入而詳細(xì)的研究。
參考文獻(xiàn)
[1] KONG W L,SUN B,ZHANG J Y,et al.Metagenomic analysis revealed the succession of microbiota and metabolic function in corncob composting for preparation of cultivation medium for Pleurotus ostreatus[J].Bioresource Technology,2020,306:123156.
[2] KOUTROTSIOS G,KALOGEROPOULOS N,STATHOPOULOS P,et al.Bioactive compounds and antioxidant activity exhibit high intraspecific variability in Pleurotus ostreatus mushrooms and correlate well with cultivation performance parameters[J].World Journal of Microbiology and Biotechnology,2017,33(5):98.
[3] LIU Q,KONG W L,HU S J,et al.Effects of Oudemansiella radicata polysaccharide on postharvest quality of oyster mushroom (Pleurotus ostreatus)and its antifungal activity against Penicillium digitatum[J].Postharvest Biology and Technology,2020,166:111207.
[4] HERN?NDEZ D,S?NCHEZ J E,YAMASAKI K.A simple procedure for preparing substrate for Pleurotus ostreatus cultivation[J].Bioresource Technology,2003,90(2):145-150.
[5] 聞亞美,邵欣欣,丁亞通,等.7種平菇發(fā)酵料生產(chǎn)拌料用殺菌劑藥效對(duì)比及安全性評(píng)價(jià)[J].中國(guó)瓜菜,2020,33(8):49-52.
[6] 余敏佳,婷婷,范麗軍,等.培養(yǎng)料中玉米秸和玉米芯的不同顆粒度對(duì)栽培雙孢蘑菇效應(yīng)[J].食藥用菌,2018,26(5):289-293.
[7] 李秋月,馮占,金宇昌,等.不同產(chǎn)地玉米芯的持水率及其相關(guān)性分析[J].食藥用菌,2020,28(6):412-417.
[8] WEI H W,WANG L H,HASSAN M,et al.Succession of the functional microbial communities and the metabolic functions in maize straw composting process[J].Bioresource Technology,2018,256:333-341.
[9] CARRASCO J,PRESTON G M.Growing edible mushrooms: a conversation between bacteria and fungi[J].Environmental Microbiology,2020,22(3):858-872.
[10] 雷露,吳天祥,王川南.基于UPLC-QTOF-MS代謝組學(xué)研究灰樹(shù)花發(fā)酵的代謝差異[J].菌物學(xué)報(bào),2020,39(10):1920-1932.
[11] BOLTEN C J,KIEFER P,LETISSE F,et al.Sampling for metabolome analysis of microorganisms [J].Analytical Chemistry,2007,79(10):3843-3849.
[12] 尉杰,趙鋮,陸瑞利,等.菌物研究中的代謝組學(xué)應(yīng)用及存在的問(wèn)題[J].菌物研究,2020,https://doi.org/10.13341/j.jfr.2020.0003.
[13] 劉志強(qiáng).平菇發(fā)酵料袋栽技術(shù)[J].現(xiàn)代農(nóng)業(yè)科技,2019(24):58-59.
[14] 孔維麗,崔筱,袁瑞奇,等.河南省適宜發(fā)酵料栽培的平菇優(yōu)良品種篩選[J].中國(guó)食用菌,2019,38(11):23-28.
[15] 張萌,羅芳,王敏,等.奶牛分娩后早期血漿代謝物變化研究[J].生物技術(shù)通報(bào),2020,36(6):191-199.
[16] 蔣立銳.組氨酸在代謝中的作用[J].生理科學(xué)進(jìn)展,1985,16(2):174-176.
[17] 李偲奇.不同料型日糧對(duì)育肥羊生產(chǎn)性能胃腸道微生物組和代謝組的影響研究[D].山東泰安:山東農(nóng)業(yè)大學(xué),2020.
[18] 賈玲.玉米芯木質(zhì)纖維素組分分離研究[D].天津:天津大學(xué),2013.
[19] 梁叢穎,林璐.環(huán)境微生物介導(dǎo)的木質(zhì)素代謝及其資源化利用研究進(jìn)展[J].微生物學(xué)通報(bào),2020,47(10):3380-3392.
[20] 孔維麗,郭家穩(wěn),劉芹,等.平菇培養(yǎng)料發(fā)酵度快速測(cè)定指標(biāo)的建立[J].中國(guó)瓜菜,2021,34(2):54-60.
[21] 崔筱,張玉亭,劉芹,等.平菇生長(zhǎng)發(fā)育過(guò)程中IAA含量及IAA氧化酶活性變化研究[J].天津農(nóng)業(yè)科學(xué),2020,26(10):31-36.
[22] XIANG Q J,LUO L H,LIANG Y H,et al.The diversity, growth promoting abilities and anti-microbial activities of bacteria isolated from the fruiting body of Agaricus bisporus[J].Polish Journal of Microbiology,2017,66(2):201-207.
[23] 胡素娟,宋凱博,劉芹,等.玉米芯發(fā)酵料水浸提液對(duì)平菇菌絲的促生長(zhǎng)機(jī)理研究[J].河南農(nóng)業(yè)大學(xué)學(xué)報(bào),2021,https://doi.org/10.16445/j.cnki.1000-2340.20210306.002.
