綠原酸的生理功能及其在畜禽生產中的應用
2021-06-05 02:25:00侯扶江張宏福
動物營養學報 2021年5期
萬 凡 侯扶江 伊 寶 張宏福
(1.中國農業科學院北京畜牧獸醫研究所,動物營養學國家重點實驗室,北京 100193;2.蘭州大學草地農業科技學院,草地農業生態系統國家重點實驗室,農業農村部草牧業創新重點實驗室,蘭州 730020)
隨著畜禽養殖規模化、集約化的不斷發展,養殖環境污染等問題越來越嚴重,由此影響畜禽健康,導致動物免疫力低下、易受病原菌侵襲等。在畜禽飼糧中添加抗生素可以增強動物自身免疫力,但卻導致抗生素殘留和細菌耐藥性等問題,嚴重危害畜禽養殖環境和畜產品質量安全。因此,開發安全高效、無殘留的新型飼料添加劑成為科研工作者的研究重點。綠原酸作為一種廣泛存在于天然植物中的多酚類物質,具有抗氧化[1]、抗炎[2]、抑菌[3]、抗病毒[4]及抗癌[5]等生物學功能。其中,金銀花、山銀花、杜仲葉以及咖啡豆等是綠原酸的主要來源[6]。研究發現,添加綠原酸能夠緩解動物腸道氧化應激和炎癥,維持腸道穩態和免疫機能,保障動物腸道健康[7-8]。因此,綠原酸可作為一種潛在的增強動物自身免疫力的飼料添加劑。本文就國內外研究現狀,簡要介紹綠原酸的來源,系統總結綠原酸的生理功能及其可能的作用機理,并探討其作為飼料添加劑在畜禽生產中的應用。
1 綠原酸
綠原酸在植物中廣泛存在,屬于苯丙烯酸(羥基肉桂酸)家族,具有6個異構體,分別為隱綠原酸(4-咖啡酰奎寧酸)、新綠原酸(5-咖啡酰奎寧酸)、異綠原酸A(3,5-二咖啡酰奎寧酸)、異綠原酸B(3,4-二咖啡酰奎寧酸)、異綠原酸C(4,5-二咖啡酰奎寧酸)和洋薊素(1,3-二咖啡酰奎寧酸)[9](圖1)。Payen[10]于1864年首次報道發現綠原酸,其從咖啡中提取出一種物質,約占咖啡總量的5%,發現該種物質在堿性氧化物的作用下變為綠色,并將其命名為“綠原酸”。后續有報道指出,奎寧酸和咖啡酸能合成一種白色晶體物質,即為綠原酸,具體為奎寧酸與咖啡酸結合形成半綠原酸,2分子半綠原酸縮合,失去1分子水生成綠原酸[11]。21世紀初,隨著質譜和高效液相色譜技術的發展,綠原酸的分子結構和化學性質逐漸被清晰認識[12],并且研究發現,綠原酸具有抗氧化[1,13]、抗炎[2]、抑菌[3,6]、抗病毒[4]及抗癌[5]等多種生理功能,目前已被應用于保健食品、醫藥等領域。

圖1 綠原酸及其異構體的分子結構
綠原酸主要提取自金銀花、山銀花、杜仲葉和元寶楓葉等中草藥的莖葉。我國中草藥資源豐富,這為綠原酸的開發和利用提供了有力保障。據國家林業局頒布的《全國杜仲產業發展規劃(2016—2030年)》[14],預計2030年,我國杜仲種植面積將達到233.33萬hm2。杜仲植株的葉子是綠原酸的主要提取部位,不同的季節綠原酸的含量不同,以6月含量最高[15]。元寶楓是槭樹科槭樹屬的一種落葉喬木,其楓葉中含有綠原酸和黃酮等多種活性成分,與杜仲類似,6月份綠原酸含量最高,可達4%左右[16]。此外,忍冬科植物山銀花(又名山花)廣泛分布于我國南方各地,尤其在四川、廣東和廣西等地,也是綠原酸的重要來源之一。
2 綠原酸在動物體內的代謝途徑
綠原酸在動物胃和小腸中,主要代謝生成咖啡酸和奎寧酸[6]。研究表明,只有約1/3綠原酸在胃和小腸被吸收,其主要原因在于綠原酸的親水性使其較難通過親脂性的膜屏障[17]。細胞培養試驗發現,綠原酸及其異構體在胃上皮細胞的吸收機制,發現均以細胞旁途徑轉運、被動擴散方式吸收,其中異綠原酸(異綠原酸A、異綠原酸B和異綠原酸C)、綠原酸、新綠原酸以及隱綠原酸的表觀滲透系數約為0.2 cm/s,時間和濃度梯度呈線性關系[18]。另外,大鼠胃結扎后灌服綠原酸,在胃的靜脈和主動脈均檢測到綠原酸[19],這也證實了綠原酸在胃內以原型被吸收。Stalmach等[20]研究發現,給健康人飲用咖啡(含有412 μmol的綠原酸)后1 h,在血漿中檢測到綠原酸的代謝產物咖啡酸-3′-硫酸鹽、阿魏酸-4′-硫酸鹽以及3-O-咖啡酰基奎寧內酯硫酸鹽,說明綠原酸在小腸中發生分解。而小鼠小腸灌注綠原酸,發現部分綠原酸在小腸中被糖苷酶和酯酶分解,酯鍵斷裂生成奎寧酸和咖啡酸[21];其中部分咖啡酸被腸道吸收,通過肝腸循環進入肝臟,咖啡酸的羥基兒茶酚鄰位在甲基轉移酶的作用下發生甲基化,形成鄰位羥基-甲氧基,轉化為阿魏酸和異阿魏酸[21]。部分阿魏酸在還原酶的作用下生成二氫阿魏酸,阿魏酸、異阿魏酸以及二氫阿魏酸由肝靜脈進入血液循環[21-22],部分發生硫酸鹽化,分別生成阿魏酸-4′-硫酸鹽、異阿魏酸-4′-硫酸鹽以及二氫阿魏酸-4′-硫酸鹽[21]。另外,小鼠小腸灌注綠原酸,在血漿中并未檢測到奎寧酸及其代謝產物,可能是奎寧酸并未在小腸中被吸收,而是在結腸被腸道菌群代謝[23]。再者,小腸中部分綠原酸被腸道直接吸收,進入血液循環系統,發生硫酸鹽化反應生成硫酸鹽代謝產物[6,24]。
未被胃和小腸吸收的綠原酸直接進入大腸,約占總量的2/3,結腸對其代謝和吸收發揮關鍵作用[23]。大部分綠原酸在結腸被大腸桿菌、雙歧桿菌和加氏乳桿菌等菌群產生的酯酶水解生成咖啡酸和奎寧酸[23]。還有部分綠原酸發生酯基位置異構形成新綠原酸和隱綠原酸,進入血液循環系統,發生硫酸鹽化反應分別生成5-O-咖啡酰基奎寧內酯硫酸鹽和4-O-咖啡酰基奎寧內酯硫酸鹽[21]。綠原酸分解生成的咖啡酸有2種代謝途徑:1)通過細胞旁途徑轉運和被動擴散方式吸收[22],進入肝臟進行后續代謝[25],在肝臟中還原酶作用下發生氫化作用生成二氫咖啡酸,二氫咖啡酸通過肝靜脈進入血液循環,部分二氫咖啡酸發生硫酸鹽化,生成二氫咖啡酸-3′-硫酸鹽[23],或者與小腸中咖啡酸進入肝臟代謝途徑相同[23];2)未被腸道吸收的一部分咖啡酸由腸道菌群作用發生甲基化反應生成阿魏酸和異阿魏酸,而后被腸道直接吸收,進入血液循環,發生硫酸鹽化反應,分別生成阿魏酸-4′-硫酸鹽和異阿魏酸-4′-硫酸鹽[22]。還有一種途徑是阿魏酸、異阿魏酸以及腸道中未降解的咖啡酸在腸道中發生葡糖苷酸化代謝,以葡糖苷酸化形式吸收[26]。綠原酸分解生成的奎寧酸在結腸微生物的作用下,代謝產生馬尿酸和香豆素[27]。其中馬尿酸的生成途徑為:奎寧酸在腸道菌群的作用下首先脫去1分子水形成莽草酸,而后莽草酸脫甲基形成苯甲酸,苯甲酸與甘氨酸結合形成馬尿酸隨尿液排出體外[27]。奎寧酸分解生成的香豆素被腸道直接吸收進入血液循環。研究證實,口服綠原酸,在動物的血漿和尿液中均檢測到綠原酸的代謝產物阿魏酸、馬尿酸以及香豆素[19]。另外,酯化后生成的新綠原酸在兒茶酚-O-甲基轉移酶作用下發生甲基化反應生成5-阿魏酰奎寧酸,進而被腸道微生物產生的酯酶水解形成阿魏酸和奎寧酸。后續代謝途徑與上述相同[22]。綜上所述,綠原酸在體內的主要代謝途徑如圖2所示。

圖2 綠原酸的主要代謝途徑
3 綠原酸的生理功能及其作用機理
3.1 抗氧化
酚類物質因含有酚羥基而具有抗氧化作用,羥基的位置和數量決定酚類物質抗氧化能力的強弱。綠原酸分子結構包含5個酚羥基和1個羧基,其中酚羥基易與自由基發生反應,形成具有抗氧化作用的氫自由基,因此,綠原酸具有較強的清除自由基和抗氧化的生物學功能[28]。細胞色素P450 2E1(cytochrome P450 2E1,CYP2E1)和細胞色素P450 4A(cytochrome P450 4A,CYP4A)均屬于細胞色素P450[29]。當機體發生氧化應激時,活性氧(reactive oxygen species,ROS)增加,CYP2E1能夠輔助脂質發生過氧化,改變呼吸鏈,進一步誘導產生更嚴重的氧化應激[29]。而CYP4A是一種氧化應激的引發劑,能夠促進肝臟脂質過氧化,其在肝臟中的表達上調也伴隨ROS的產生[30]。體外HepG2細胞試驗發現,高濃度的綠原酸(2 mmol/L)通過抑制CYP2E1和CYP4A的表達來改善油酸誘導的細胞氧化損傷[31]。綠原酸還能調控相關信號通路發揮抗氧化作用。在乙酰氨基酚誘導的小鼠肝臟急性衰竭模型中,提前口服綠原酸能夠緩解肝臟損傷,主要作用機制是通過增加血紅素加氧酶-1(heme oxygenase-1,HO-1)和NADPH醌脫氫酶1基因的表達量,激活細胞核因子E2相關因子2(nuclear factor E2 related factor 2,Nrf2)信號通路,發揮自身的抗氧化作用[32]。類似的,在四氯化碳誘導小鼠肝纖維化的損傷模型研究中,給小鼠腹腔注射金銀花提取物(主要成分為綠原酸),同樣發現能激活Nrf2信號通路,降低氧化應激,緩解肝損傷[33]。此外,還有報道指出,綠原酸能夠降低氧化應激誘導的白細胞介素-8的分泌及其轉錄表達[34]。另外,肉雞飼糧中添加1 000 mg/kg綠原酸可提高肉雞的抗熱應激能力和雞胸肉的氧化穩定性[35];斷奶仔豬飼糧中添加1 000 mg/kg綠原酸,可顯著提高仔豬血清中超氧化物歧化酶、谷胱甘肽過氧化物酶以及過氧化氫酶的活性,并且降低腹瀉率,表明綠原酸具有提高動物機體的抗氧化能力的作用[36]。總之,綠原酸通過降低ROS水平和激活Nrf2信號通路降低或阻止氧化應激的發生,達到抗氧化的效果,其作為飼料添加劑可提高動物機體的抗氧化能力。
3.2 抗炎
炎癥反應是機體保護自身的一種免疫反應,但炎癥因子的過度激活會導致組織損傷、器官衰竭,如不能及時治療甚至會造成死亡[37]。綠原酸具有抗炎作用,可下調促炎因子的表達量,提高機體的抵抗力。總結綠原酸發揮抗炎作用的研究顯示:1)綠原酸通過介導核轉錄因子-κB(nuclear factor-κB,NF-κB)通路發揮抗炎功能[38]。Zamani-Garmsiri等[39]研究發現,非酒精性脂肪肝小鼠飼喂含有綠原酸(0.2 g/kg)的飼糧,12周后,綠原酸抑制了多囊性腎病NF-κB信號通路中p65蛋白的移位和核轉錄因子-κB抑制蛋白激酶(nuclear factor-kappa B inhibitor protein kinase,IκB)的磷酸化,顯著降低腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)、白細胞介素-1β(interleukin-1β,IL-1β)以及白細胞介素-6(interleukin-6,IL-6)等促炎因子的表達量,從而實現抗炎癥效果。類似報道證明,綠原酸可以降低脂多糖(LPS)誘導的乳腺上皮細胞中IL-1β和IL-6的轉錄表達,并抑制NF-κB p65蛋白的表達[40]。2)細胞試驗結果表明,綠原酸能夠緩解LPS誘導的RAW 264.7小鼠巨噬細胞炎癥,通過抑制環氧合酶-2和NOD樣受體家族蛋白3(NOD-like receptor family protein 3,NLRP3)炎癥小體的蛋白表達,發揮抗炎作用[38]。3)在緩解糖尿病和腎病的研究中發現,綠原酸能增加Nrf2的核易位和HO-1的表達,通過調節Nrf2/HO-1信號通路發揮抗炎作用[41];Zhao等[42]利用隱綠原酸對LPS處理的RAW 264.7巨噬細胞炎癥反應進行干預,發現隱綠原酸使HO-1基因的表達量上升,能夠激活Nrf2信號通路發揮抗炎作用。4)綠原酸可介導絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)通路發揮抗炎功能[43]。綠原酸能夠緩解葡聚糖硫酸鈉誘導的小鼠結腸炎性損傷,降低細胞外信號調節蛋白激酶(extracellular signal-regulated protein kinase,ERK)1/2、磷酸化細胞外調節蛋白激酶、p38激酶、磷酸化p38激酶、c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)以及磷酸化c-Jun氨基末端激酶蛋白的表達,通過抑制MAPK/ERK/JNK信號通路,緩解小鼠腸道炎癥[43]。Tan等[44]研究發現,綠原酸能夠緩解鼠傷寒沙門氏菌誘導的小鼠腸道炎癥,主要機制為綠原酸促使腸道中生長阻滯特異性轉錄因子5(growth arrest specific 5,GAS5)的表達,而后長鏈非編碼RNA GAS5競爭性結合微小RNA 23a(microRNA 23a,miR-23a),進而上調第10號染色體缺失的磷酸酶與張力蛋白同源物基因(phosphatase and tensin homolog deleted on chromosome 10,PTEN)的表達量,降低p38 MAPK通路相關基因p38和磷酸化p38蛋白的表達水平,通過長鏈非編碼RNA GAS5/miR-23a/PTEN軸抑制p38 MAPK信號通路,從而抑制腸道炎癥。
3.3 抑菌
綠原酸是金銀花、山銀花和杜仲等中草藥的主要抑菌成分[6]。綠原酸對包括大腸桿菌、沙門氏菌、痢疾桿菌、假單胞菌屬及幽門螺桿菌屬等革蘭氏陰性菌和包括金黃色葡萄球菌、芽孢桿菌、鏈球菌及葡萄球菌屬等革蘭氏陽性菌均有明顯的抑制作用[6]。Chai等[45]認為綠原酸抑菌作用是由于綠原酸協同抑制了細菌細胞壁和細胞膜的合成,并抑制了細菌蛋白和DNA的合成。動物試驗表明,綠原酸不僅抑制腸道有害細菌,還可促進腸道有益菌的增殖。Gong等[46]研究發現,金黃色葡萄球菌誘導奶牛乳腺炎,添加綠原酸可提高牛奶中κ-酪蛋白的水平,增強中性粒細胞對病原菌的吞噬能力。另外,綠原酸可以增加斷奶仔豬回腸和盲腸揮發性脂肪酸含量,提高乙酸鹽和丙酸鹽的含量,還可增加盲腸菌群多樣性,提高盲腸中乳酸菌屬、普氏桿菌屬和厭氧弧菌屬等有益菌屬的豐度,維持仔豬腸道菌群穩態,保護腸道健康[47-48]。
綠原酸的抗菌機制可能與以下有關:1)綠原酸可作用于病原菌的細胞外膜,促使其凹陷和脫落,并降低細胞外膜LPS的含量,使細胞外膜通透性增加,引起細胞內蛋白質和ATP的釋放,導致細胞外膜破壞、代謝中斷,最終導致細胞死亡[49]。Sun等[8]在冷藏雞肉中評估綠原酸對沙門氏菌病原體的抗菌活性,發現綠原酸能夠改變沙門氏菌細胞外膜的通透性,促進胞內蛋白質和ATP的釋放,并且抑制蘋果酸脫氫酶和琥珀酸脫氫酶的活性,破壞細胞膜和細胞代謝,引發細胞死亡。2)在發光二極管(LED)光照射下,綠原酸進入病菌細胞膜發生光氧化,生成過氧化氫(H2O2),H2O2在光的作用下分解生成羥基自由基(·OH),·OH使細胞內DNA發生氧化損傷,導致DNA斷裂、細胞凋亡[50]。3)綠原酸可作用于細菌中的酶,發揮抗菌作用。肺炎鏈球菌神經氨酸酶具有去除宿主細胞表面聚糖末端的唾液酸殘基,促進病菌黏附和定殖的作用。最新研究發現,綠原酸通過氫鍵、鹽橋與肺炎鏈球菌神經氨酸酶建立較強的結合,涉及的氨基酸殘基包括Arg347、Ile348、Lys440、Asp372、Asp417以及Glu768,其中與Arg347的側鏈可形成帽狀結構,將綠原酸鎖定在神經氨酸酶的活性位點,并與其發生結合,因此綠原酸是一種潛在的肺炎鏈球菌神經氨酸酶抑制劑[51]。4)綠原酸可以誘導細胞內ROS耗竭,最終影響細菌凋亡。ROS是細胞內信號傳導的重要分子,細胞內ROS耗竭是細菌凋亡的核心調節因子。研究表明,先在大腸桿菌中添加綠原酸,而后用H2O2進行處理,綠原酸能誘導大腸桿菌中ROS耗竭,進而影響大腸桿菌細胞內的信號通路傳導,從而導致細菌凋亡[13]。
3.4 抗病毒
綠原酸具有抗病毒作用,對流感病毒[52]、呼吸道合胞病毒[53]、皰疹病毒[54]、腸道病毒[55]、H5N1病毒[52]、腺病毒[53]和乙型肝炎病毒[56]均有明顯抑制作用。關于綠原酸抗病毒的機理還不是很清楚,根據文獻報道,推測綠原酸的抗病毒機制可能與以下幾點有關:1)綠原酸可以抑制病毒表面酶的活性。Karar等[52]研究指出,神經氨酸酶是流感病毒被膜上的一種糖蛋白,具有抗原性,能催化糖苷水解,協助成熟的病毒顆粒脫離宿主細胞并感染新的細胞,造成流感病毒在體內繼續擴散。綠原酸、酚酸和衍生物對產氣莢膜桿菌、H5N1和重組H5N1 N端組氨酸標簽的神經氨基酶有抑制活性的作用。綠原酸通過與神經氨酸酶結合,達到抑制病毒在動物體內擴散的目的。2)綠原酸的抗病毒活性與其自身含有的鄰苯二酚基團有關,Ikeda等[57]研究發現,可能是由于綠原酸水解出的具有鄰苯二酚基團的咖啡酸,而咖啡酸能抑制皰疹病毒的活性,干擾皰疹病毒DNA的復制,從而發揮抗病毒的作用。
3.5 抗腫瘤
上世紀80年代,綠原酸被發現具有抗腫瘤的作用,引起了科研工作者的廣泛興趣。Xue等[58]首次報道,綠原酸能夠促進M1型巨噬細胞和抑制M2型巨噬細胞,抑制腦膠質瘤的生長,提出作用途徑是介導巨噬細胞的M1/M2極化。也有報道指出,綠原酸能降低癌細胞的增殖速率、線粒體ATP的產生以及侵襲能力,作用方式是綠原酸提高癌細胞分化基因小泛素相關修飾物1(small ubiquitin-related modifier 1,SUMO1)的表達,同時不改變正常細胞中該基因的表達量。因此,綠原酸有望成為一種安全有效的癌細胞分化誘導劑[59]。此外,有研究認為,綠原酸緩解結腸癌的作用是通過降低ROS發揮抑制作用,還可通過降低細胞凋亡相關基因的表達抑制肺癌細胞生長[60-61]。因此,綠原酸具有抗癌作用,但作用方式不盡相同,其潛在靶點和作用機制還有待進一步研究。綜上所述,綠原酸具有抗氧化、抗炎、抑菌、抗病毒及抗癌等多種生物學功能,其可能的作用機理如圖3所示。
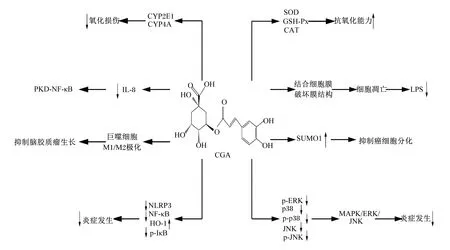
↑:提高 increased;↓:降低 decreased;CGA:綠原酸 chlorogenic acid;CYP2E1:細胞色素P450 2E1 cytochrome P450 2E1;CYP4A:細胞色素P450 4A cytochrome P450 4A;SOD:超氧化物歧化酶 superoxide dismutase;GSH-Px:谷胱甘肽過氧化物酶 glutathione peroxidase;CAT:過氧化氫酶 catalase;PKD-NF-κB:多囊性腎病核轉錄因子-κB nuclear factor kappa B in polycystic kidney disease;IL-8:白細胞介素-8 interleukin-8;LPS:脂多糖 lipopolysaccharide;SUMO1:小泛素相關修飾物1 small ubiquitin-related modifier 1;NLRP3:NOD樣受體家族蛋白3 NOD-like receptor family protein 3;NF-κB:核轉錄因子-κB nuclear factor kappa B;HO-1:血紅素加氧酶-1 heme oxygenase-1;p-IκB:磷酸化核轉錄因子-κB抑制蛋白激酶 phosphorylated nuclear factor-kappa B inhibitor protein kinase;ERK:細胞外信號調節蛋白激酶 extracellular signal-regulated protein kinase;p-ERK:磷酸化細胞外信號調節蛋白激酶 phosphorylated extracellular signal-regulated protein kinase;p38:p38激酶 p38 kinase;p-p38:磷酸化p38激酶 phosphorylated p38 kinase;JNK:c-Jun氨基末端激酶 c-Jun N-terminal kinase;p-JNK:磷酸化c-Jun氨基末端激酶 phosphorylated c-Jun N-terminal kinase;MAPK:絲裂原活化蛋白激酶 mitogen-activated protein kinase。
4 綠原酸在畜禽生產中的應用
4.1 家畜
規模化養豬生產中,由于早期斷奶、病原菌侵襲等,仔豬發生斷奶應激和腸道穩態失衡,造成腸炎和腹瀉[62]。王宇等[63]研究發現,與對照組相比,飼糧添加1 000 mg/kg的綠原酸顯著降低斷奶仔豬的料重比,認為其可能的機制是綠原酸提高了仔豬十二指腸黏膜中蔗糖酶活性和回腸黏膜中鈉-葡萄糖共轉運載體1(sodium-glucose co-transporter 1,SGLT1)mRNA的表達量,而SGLT1是位于小腸黏膜上皮細胞和腎單位近曲小管的一種跨膜葡萄糖轉運載體蛋白,可以促進葡萄糖的吸收和轉運。Zhang等[64]在斷奶仔豬的飼糧中添加1 000 mg/kg綠原酸,發現試驗組仔豬十二指腸的絨毛高度和絨隱比顯著提高,結腸食糜乳酸菌的豐度提高,大腸桿菌的豐度顯著降低,說明綠原酸可以保護豬的腸道健康。另外,綠原酸還可降低斷奶仔豬十二指腸和空腸中丙二醛的含量,緩解腸道氧化損傷,改善腸道屏障功能[62]。因此綠原酸具有調節仔豬腸道菌群,促進營養物質的消化吸收,維持腸道穩態的作用。
研究發現,綠原酸能夠改善母豬卵母細胞成熟、發育及受精能力。與對照組相比,體外補充綠原酸可以顯著促進卵母細胞成熟,保護卵母細胞免受氧化應激誘導的DNA損傷,同時提高胚泡形成的速率,改善豬胚胎體外發育能力[65]。另有報道指出,添加50 μmol/L綠原酸與卵母細胞共培養44 h后再進行體外受精,通過電穿孔技術檢測受精卵的發育和質量,發現綠原酸顯著提高胚泡的生長速率,降低細胞凋亡指數,改善胚胎發育[66]。綠原酸還可提高冷凍公豬精子的質量,補充100 μmol/L綠原酸,公豬精子存活率提高了8%~10%,質膜完整性提高了4%~6%[67],可替代維生素E作為豬精液的保護劑[68]。以上研究說明,綠原酸對母豬卵母細胞和公豬精子發育均有積極影響,可作為卵母細胞發育成熟和精子的保護劑,但綠原酸是否作用于抗氧化相關信號通路,實現促進母豬卵母細胞成熟和緩解公豬精液的氧化應激,目前仍不清楚,具體的作用機制仍需要進一步研究。
綠原酸在反芻動物上的研究,主要關注其抗氧化性能和對營養物質消化代謝的影響。Bongartz等[69]探究了堿處理對葵花粕中綠原酸和蛋白質的影響,發現堿處理增加了過瘤胃蛋白的含量,與對照組相比,堿處理組奶牛腸道蛋白質消化率提高了10%,原因在于經堿處理生成的綠原酸醌和葵花粕中的蛋白質發生反應,降低了瘤胃微生物對蛋白質的降解,增加了過瘤胃蛋白的含量。除了直接應用于反芻動物飼糧,因其抗氧化性能,綠原酸還在牛奶乳制品中表現出積極的影響。研究發現,在乳桿菌發酵乳中加入綠原酸,牛奶的抗氧化活性增加,這是由于綠原酸被乳桿菌發酵代謝生成3,4-二羥基肉桂酸,同時乳桿菌的蛋白水解活性增加了生物活性肽的釋放量,以達到抗氧化的作用[70]。另外,Liu等[7]在育肥羊飼糧中添加綠原酸,發現顯著提高了羊血清中超氧化物歧化酶活性和總抗氧化能力,顯著降低了血清中皮質醇含量。盡管綠原酸的抗氧化功能被一再證實,但由于反芻動物特有的消化器官,還需要著重研究綠原酸對反芻動物胃腸道健康、胃腸道菌群的調控機理以及作為乳制品添加劑的應用價值。
4.2 家禽
綠原酸對家禽影響的研究,主要關注改善生長性能、減緩熱應激以及緩解炎癥等。Zhao等[35]在白羽肉雞飼糧中添加1 000 mg/kg的綠原酸,肉雞的平均日增重和飼料轉化率顯著優于對照組,且添加綠原酸顯著提高了雞肉中多不飽和脂肪酸和飽和脂肪酸的比值;不僅如此,在熱應激條件下,飼糧添加綠原酸還顯著提高了雞胸肉的氧化穩定性和脂肪酸含量。綠原酸緩解熱應激的研究,可能的機制是綠原酸通過增加低氧誘導因子-1的活性,促進熱休克轉錄因子-1基因表達,增強自噬,提升機體抗熱應激的能力[71],而提高雞胸肉的氧化穩定性可能主要源于綠原酸的抗氧化性。此外,綠原酸還能夠緩解雛雞受損的軟骨,促使其恢復和再生。Cheng等[72]使用靜脈注射甲苯噻嗪(5 mg/kg)的方法建立雛雞的關節軟骨損傷模型,發現飼喂綠原酸可上調Y染色體性別決定區-盒轉錄因子9(sex-determining region Y-box transcription factor 9,SOX9)和Ⅱ型膠原α1鏈(collagen type Ⅱ alpha 1 chain,COL2A1)這2個基因的表達。而SOX9是軟骨生成所需的轉錄激活因子,COL2A1是SOX9的1個下游基因,二者上調可以促進軟骨生長[72]。另外,綠原酸抑制損傷部位血管異常生成和纖維化,下調炎癥細胞因子IL-1β和TNF-α、磷酸化p65以及金屬蛋白酶基因的表達,可能的作用機制是綠原酸通過作用于Nrf2信號通路緩解氧化應激,恢復受損的關節軟骨[72]。在水禽的研究中,給1日齡北京鴨接種鴨乙型肝炎病毒(duck hepatitis B virus,DHBV)陽性血清,建立鴨乙肝病毒感染模型,7~17日齡每天口服100 mg/kg的綠原酸,發現口服綠原酸降低了鴨血清中DHBV的水平,抑制了病毒DNA的復制[73],說明綠原酸具有抗DHBV的作用,但具體機制還有待進一步研究。另外,關于綠原酸改善家禽生產性能的作用機制,是否通過作用于腸道微生物區系提高腸道優勢菌群豐度以促進營養物質的消化吸收,還有待研究。
5 小 結
近年來,因其具有抗氧化、抗炎、抑菌、抗癌、提高繁殖性能以及保護畜禽腸道健康等特性,綠原酸的研究呈上升趨勢,成為天然植物提取物研究領域的熱點之一。我國綠原酸的植物來源廣泛、產量高,作為飼料添加劑,在畜禽生產中具有極大的前景。未來對綠原酸的研究應集中在以下方面展開:1)綠原酸的作用機制有待研究。目前綠原酸的作用機理尚不完全清楚,后續的研究需關注綠原酸的作用靶點,探索綠原酸作用的分子機制及其酯水解產物,甚至不同代謝產物對腸道健康的影響及其機制。2)綠原酸的利用方式有待研究。由于綠原酸口服效果不佳,如何提高畜禽對綠原酸的利用效率,也是后續研究的重點之一。3)作為飼料添加劑,綠原酸在畜禽生產中的適宜劑量還有待確定。綜上可知,系統研究綠原酸的代謝過程和生物學功能,揭示其作用的分子機制及其高效利用方式,對未來綠原酸成為畜牧業生產中的新型飼料添加劑至關重要。
