廣西犬源H9N2亞型流感病毒分離株的全基因組測序分析及其對小鼠的致病性實驗
2021-06-03 08:18:50李三木孫翔翔孔子榮何奇松馮淑萍楊可妍曾詠芳顏健華
中國人獸共患病學報 2021年5期
關鍵詞:小鼠
李三木,孫翔翔,孔子榮,何奇松,馬 琳,馮淑萍,楊可妍,曾詠芳,熊 毅,顏健華
H9N2亞型流感病毒屬于A型流感病毒,正粘病毒科,基因組分8個節(jié)段編碼約11種蛋白。H9N2亞型流感病毒首次分離是1966年從意大利的火雞身上分離出毒株A/Turkey/Wisconsin/1/16[1],國內(nèi)在1994年于廣東省的雞群中第一次分離到該亞型[2]。雖然H9N2亞型流感病毒具有低致病性的特點[3-4],但其宿主范圍廣,不僅能夠侵染禽類、野鳥,還能感染包括豬在內(nèi)的哺乳動物。相對其他亞型病毒,更容易跨物種在人群中傳播[5-6]。流行病學和病毒學證據(jù)表明,目前從人身上分離的H9N2病毒都來自禽類,沒有發(fā)現(xiàn)人傳人的病例,并且在基因上與當?shù)丶仪葜辛餍械腍9N2病毒相似[7-8]。
自然條件下犬不太容易感染上流感病毒,實驗也沒有發(fā)現(xiàn)犬流感的流行[9],既往有研究中發(fā)現(xiàn),犬能被多種不同亞型的流感病毒所感染,包括H3N8[10]、H3N2[11]、H5N1[12]、H1N1[13]、H5N2[14]等,而且也有研究表明犬對H9N2亞型禽流感病毒易感,并且能夠形成種間傳播[15-16],我國養(yǎng)犬人群基數(shù)很大,犬作為人類伴侶動物能夠與人親密頻繁接觸,無疑增加了人感染H9N2流感病毒的風險[17]。對廣西犬只尤其是寵物犬中分離出的13株H9N2亞型流感病毒A/Canine/Guangxi/1~13/2011(H9N2)進行分子生物學特性分析,了解分離病毒株的來源和遺傳特征,再通過關鍵位點分析,了解分離毒株的抗原性、耐藥性,并選取其中部分毒株進行哺乳動物致病性分析,為廣西犬源H9N2亞型流感病毒的防控提供科學依據(jù)。同時,對防控人獸共患病也具有重要的意義。
1 材料與方法
1.1試驗材料 A/Canine/Guangxi/1~13/2011(H9N2)(后文縮寫為Ca/GX/1~13)源自廣西各地動物醫(yī)院以及飼養(yǎng)場采集的犬棉拭子。SPF雞胚購自北京梅里亞維通有限公司。SPF BALB/c 雌性小鼠購自廣東省醫(yī)學實驗動物中心。病毒總RNA抽提試劑盒、dNTP、反轉(zhuǎn)錄酶(AMV)、膠回收試劑盒、質(zhì)粒提取試劑盒等購自天跟生化科技(北京)有限公司。pMD18-T載體以及DH5α感受態(tài)細胞等購自TaKaRa公司。
1.2引物設計與合成 參照GenBank上H9N2的序列,利用Primer 5.0軟件設計8對引物擴增H9N2亞型流感病毒基因組各片段。使用Uni12引物進行反轉(zhuǎn)錄。所用引物均送TaKaRa公司合成。引物序列見表1。

表1 H9N2亞型流感病毒全基因組擴增的引物
1.3核酸提取、PCR擴增 病毒尿囊液RNA抽提,使用Uni12引物經(jīng)反轉(zhuǎn)錄獲得cDNA,接著PCR,退火溫度為56 ℃。反應結束后經(jīng)1.5%瓊脂糖凝膠電泳。
1.4目的條帶回收與克隆 DNA回收純化,將回收純化好的產(chǎn)物與pMD18-T載體連接,轉(zhuǎn)化,菌液涂布到LB固體瓊脂培養(yǎng)基培養(yǎng)過夜,挑斑擴培后進行菌液PCR鑒定,抽提陽性質(zhì)粒送大連TaKaRa公司測序。
1.5病毒基因組測序及分析 利用DNAStar進行序列拼接然后與參考毒株進行同源性比對,MEGA 6.0繪制毒株遺傳進化樹并分析相關功能位點。
1.6雞胚半數(shù)感染量EID50的測定 根據(jù)對分離毒株的序列分析,選取A/Canine/Guangxi/1/2011(H9N2)、A/Canine/Guangxi/4/2011(H9N2)、A/Canine/Guangxi/8/2011(H9N2)和A/Canine/Guangxi/10/2011(H9N2)進行病毒感染實驗,毒株Ca/GX/1和Ca/GX/4自健康犬中分離得到,毒株Ca/GX/8和Ca/GX/10源自流感癥狀的犬只。經(jīng)尿囊腔接種10日齡的SPF雞胚,純化3代,根據(jù)Reed-Muench法計算病毒的雞胚半數(shù)感染量。
1.7小鼠的致病性實驗 將6周齡的雌性SPF BALB/c隨機分成5組,每組8只。按照106EID50/50 μL的濃度攻毒小鼠,攻毒第3 d,采集3只,其腦、鼻甲、脾、腎、肺用于病毒滴定和組織切片,并觀察剩余小鼠至14 d。同時進行小鼠的傳代實驗,研究H9N2病毒分離株對哺乳動物的適應能力及其與宿主轉(zhuǎn)換事件相關的突變,觀察病毒致病性的變化。體重變化率(%)=[(感染后平均體重-感染前平均體重)/感染前平均體重]×100%+100%。動物感染實驗均在廣西壯族自治區(qū)動物疫病預防與控制中心生物安全二級實驗室開展,符合生物安全條件。
2 結 果
2.1病毒全基因組的核苷酸同源性及遺傳進化分析 13個毒株基因片段的核苷酸同源性為PB1: 99.8%~100%、HA: 99.9%~100%、NA: 99.7%~100%,PB2、PA、NP、M、NS同源性均為100%。HA、NS遺傳進化與A/Chicken/Beijing/1/94在同一個分支,屬于BJ-like譜系。NA與A/Chicken/HongKong/G9/97在同一個分支,屬于G9-like譜系。M與A/Quail/HongKong/G1/97在同一個分支,屬于G1-like譜系。NP、PA、PB1基因與A/Chicken/Shanghai/F/98在同一個分支,屬于SHF98-like譜系。PB2在遺傳學上屬于一個新的分支,屬于DK/ST/163/2004-like譜系(圖1、圖2)。內(nèi)部基因M、NP、NS、PA、PB1和PB2通過遺傳進化分析發(fā)現(xiàn)與2013年在上海、杭州等地從人身上分離到的H7N9毒株關系較近,其氨基酸同源性為M:98.4%~99.6%、NP:98.0%~98.2%、NS:92.9%~93.4%、PA:100%、PB1:100%、PB2:98.9%。

圖1 H9N2病毒株的HA基因遺傳進化樹
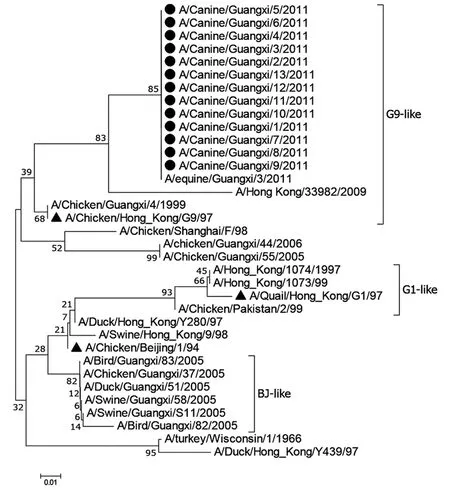
圖2 H9N2病毒株的NA基因遺傳進化樹
2.2HA氨基酸關鍵位點分析 通過氨基酸序列分析發(fā)現(xiàn)13株犬源H9N2流感病毒株的HA基因的氨基酸位點均相對保守,226位為亮氨酸(Leu),具有與SAα-2,6-Gal特異性結合的能力,裂解位點為RSSR↓GLF,位于335和341位之間(表2)。糖基化位點和受體結合位點都較保守,除了313和551位的糖基化位點與參考毒株不一致。抗原表位除234位同參考毒株不同外其他都相對保守。HA基因共發(fā)生14個位點的突變,而315位突變可能導致一個潛在糖基化位點的增加。

表2 H9N2流感病毒株HA受體結合相關氨基酸位點及連接肽序列
2.3NA氨基酸關鍵位點分析 對分離株NA基因進行的氨基酸序列分析發(fā)現(xiàn)其基質(zhì)結合位點同參考毒株一致。抗原位點331位V、367位E、432位K的突變可能導致病毒在耐藥性上的差異。NA基因潛在的糖基化位點與參考毒株相比比較保守。紅細胞結合位點在367、399、432位點處有變動與參考毒株不一致。
2.4其它氨基酸關鍵位點分析 分離株M1的氨基酸序列發(fā)生多個位點的突變,包括37(A)、46(I)、95(K)、140(A)、187(R)、219(V)、224(N)、242(N)和248(L),這些位點的突變同2013年杭州分離的毒株A/Hangzhou/1/2013(H7N9)完全一致。M2基因第31位氨基酸位點發(fā)生了S到N的突變。其它內(nèi)部基因M、NP、NS、PA、PB1和PB2通過遺傳進化分析發(fā)現(xiàn),13株毒株與2013年在上海、杭州等地暴發(fā)流感從人類身上分離得到的H7N9病毒株有較近的親緣關系。
2.5雞胚半數(shù)感染量EID50的測定 EID50通過Reed-Muench公式計算得出,4株H9N2病毒株的EID50結果如表3所示,從數(shù)據(jù)看出毒株A/Canine/Guangxi/4/2011(H9N2)在SPF雞胚上較其它毒株有略高的增殖滴度。
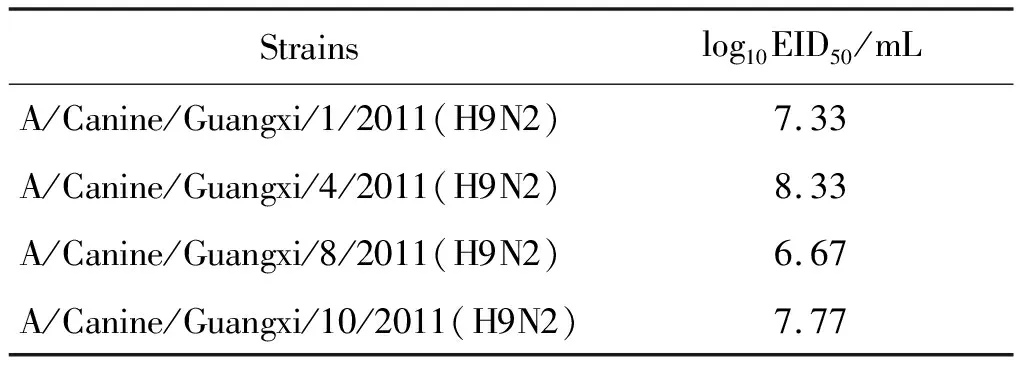
表3 廣西犬源H9N2亞型流感病毒EID50結果
2.6病毒對小鼠致病性的實驗結果 從圖3可知對照組小鼠的體重在14 d內(nèi)持續(xù)增加,實驗組也都呈增加趨勢,除Ca/GX/8組與對照組相比增加幅度相仿,其余增加幅度都較小。未出現(xiàn)小鼠死亡。
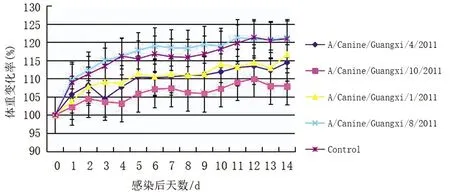
圖3 小鼠感染后體重變化率
感染3 d后,隨機剖殺3只小鼠,組織樣品勻漿處理,接種10日齡SPF雞胚進行病毒滴度實驗。4株病毒株攻毒的小鼠中脾臟、腦和腎臟中均無病毒,攻毒小鼠的肺臟和鼻甲骨中可以檢測到病毒,但同種亞型的不同毒株之間病毒的復制能力也表現(xiàn)出差異。Ca/GX/10株在肺臟組織和鼻甲骨中顯示病毒滴度,且小鼠體重變化最大,體重變化最小的Ca/GX/8株只在鼻甲骨中檢測到,而Ca/GX/1和Ca/GX/4未檢測到病毒(表4)。

表4 小鼠組織感染的病毒復制滴度(log10TCID50/mL)
對Ca/GX/10毒株感染的小鼠肺部組織樣品做組織切片,結果顯示與未感染的空白對照組相比,攻毒組小鼠肺組織有明顯的炎性細胞滲出和浸潤,肺泡壁變厚,有多處的破裂,肺泡形狀受到不同程度的破壞,大小不一,形狀不規(guī)則,見圖4。
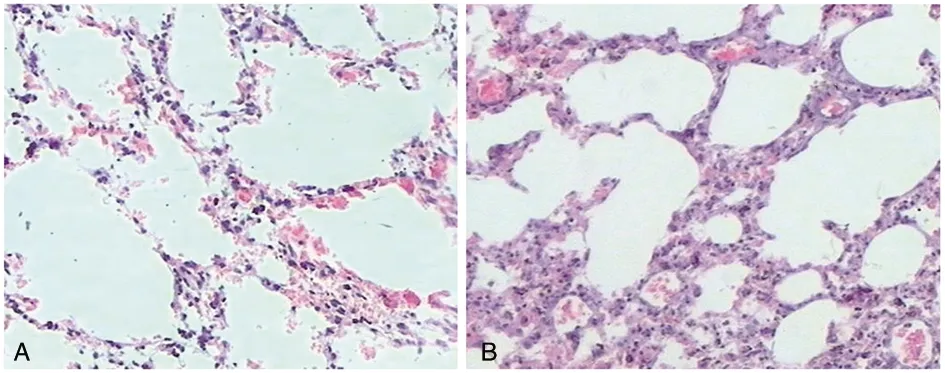
A為對照組;B為感染組
2.7病毒在小鼠中的傳代實驗結果 選用Ca/GX/8病毒株感染小鼠,連續(xù)4次傳代的結果如圖5所示,第1次體重持續(xù)增加,第2次感染小鼠后觀察出現(xiàn)被毛凌亂,精神萎靡,食欲不振等癥狀,體重較第1次在3 d變緩。第3次感染小鼠后與第2次類似,臨床癥狀更加明顯,體重變化小。第4次感染后小鼠體重基本不增加,臨床癥狀明顯,出現(xiàn)畏冷、食欲不振、精神萎靡等。
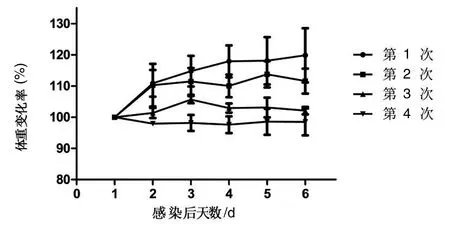
圖5 小鼠感染后體重變化情況
經(jīng)過對4次傳代后的病毒基因測序發(fā)現(xiàn),Ca/GX/8毒株PB2基因611和623氨基酸位點發(fā)生突變,分別是611位天冬氨酸突變?yōu)樘於0贰?23位由谷氨酸E突變?yōu)橘嚢彼酜;其余基因沒有發(fā)生變化。
3 討 論
本實驗所用13株H9N2流感病毒株都由采自廣西各地區(qū)的犬棉拭子中分離得到,表明廣西地區(qū)的犬只中普遍存在感染H9N2流感病毒的情況,這是世界范圍內(nèi)首次從犬只中分離得到H9N2亞型流感病毒,也表明了該亞型病毒的宿主范圍得到進一步擴大。H9N2病毒極易發(fā)生跨物種傳播,而犬類作為日常伴侶動物能夠與人類親密接觸,增加了人類感染H9N2亞型流感病毒的風險。
HA基因源于禽類,為歐亞譜系的BJ-like亞系,基因序列和中國地區(qū)的H9N2亞型都與流行代表毒株A/chicken/Guangxi/55/2005(H9N2)有較近的關系。氨基酸分析表明HA基因的226位為亮氨酸(Leu),具有與唾液酸受體SAα-2,6-Gal特異性結合的能力,表明分離得到的H9N2亞型流感病毒雖然來源于禽類,但特異性的結合受體已經(jīng)具備和人源受體特異性結合的能力,引起哺乳動物的易感[18]。HA基因的裂解位點為RSSR↓GLF,位于335和341位之間。HA1和HA2由兩個堿性氨基酸連接,說明分離的毒株為低致病力毒株[19]。HA基因的糖基化位點和受體結合位點都較保守,只有313和551位的糖基化位點與參考毒株不一致。313-315位點位于裂解位點附近,這可能會通過干擾蛋白酶裂解而增強致病性[20]。本研究分離株受體結合位點191位為N、198位為A,與禽源毒株一致,因此可以推斷其屬于禽源。
NA基因?qū)儆跉W亞譜系G9-like亞系,典型的禽源特征。13株毒株的抗原位點在331、367、432位點表現(xiàn)與參考毒株不一致,可能導致耐藥性的差異。紅細胞結合位點在367、399、432位點處與參考毒株不一致,這些位點是NA基因主要抗原位點,神經(jīng)氨酸酶的唾液酸結合位點易受抗體陽性選擇從而進化[21],推測NA基因氨基酸的變化可能是病毒逃避疫苗作用導致。
M2基因第31位氨基酸位點發(fā)生了S到N的突變,此突變導致病毒產(chǎn)生對金剛烷胺類藥物的耐藥性[22]。M1基因發(fā)生了多個氨基酸位點的突變,包括37(A)、46(I)、95(K)、140(A)、187(R)、219(V)、224(N)、242(N)和248(L),這些突變都與毒株A/Hangzhou/1/2013(H7N9)一致,突變對病毒的影響需要進一步的研究。其它內(nèi)部基因M、NP、NS、PA、PB1和PB2通過遺傳進化分析發(fā)現(xiàn),13株毒株與2013年在上海、杭州等地自人流感病毒分離株H7N9有較近的親緣關系。病毒的致病性以及不同種間傳播機制有待進一步研究。
經(jīng)同源性和系統(tǒng)進化樹分析發(fā)現(xiàn),13株病毒株的8個基因片段來源于5個經(jīng)典毒株。不屬于已有基因型A-R的劃分[23],而是一個新的基因型,屬于禽源歐亞譜系,這與華南地區(qū)的大部分禽源流感病毒一致。說明在華南地區(qū),特別是廣西地區(qū),基因型復雜,這可能是由于該地區(qū)混合飼養(yǎng)的模式,為病毒基因重組的發(fā)生提供了條件。
小鼠的致病性實驗中,健康犬中分離得到的毒株Ca/GX/1和Ca/GX/4在小鼠體內(nèi)尚未有效復制,而源自流感癥狀的犬只的Ca/GX/8和Ca/GX/10發(fā)現(xiàn)可以在小鼠體內(nèi)發(fā)生復制,Ca/GX/10毒株甚至可以出現(xiàn)顯著的臨床癥狀如體重下降、精神不振、沒有食欲以及被毛凌亂等。組織切片也顯示出炎癥反應造成的損傷。說明即使同種亞型的不同毒株在對宿主動物的致病力方面也表現(xiàn)出不同。本實驗中未出現(xiàn)小鼠死亡,對小鼠表現(xiàn)為低致病力,但不能排除其他重組毒株能夠?qū)е滤拗鲊乐氐膿p傷甚至死亡[24-26]。氨基酸位點分析,Ca/GX/8和Ca/GX/10完全一致,而Ca/GX/4同其他毒株相比氨基酸位點在PB2基因的296位發(fā)生了D到N的突變,Ca/GX/1則有多個位點的突變,HA152K→E,NA380I→T,M2的64S→A、66E→K,PB1的411M→L、422S→L、630P→L和PA210T→I,推測正是由于這些氨基酸位點的突變的綜合影響導致Ca/GX/1和Ca/GX/4對小鼠的致病性降低。流感病毒通過基因突變和重組實現(xiàn)對環(huán)境的適應[27-28],經(jīng)4次傳代后的病毒測序發(fā)現(xiàn),Ca/GX/8毒株PB2基因611和623氨基酸位點發(fā)生突變,經(jīng)查閱暫無文獻報道這兩個氨基酸位點突變有何影響,該突變對病毒致病性的影響還有待進一步的研究。
利益沖突:無
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國男科學雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
