CRISPR-Cas系統(tǒng)在微生物研究中的應用進展
2021-06-02 09:01:40格日樂其木格牛振峰董丹張濤濤崢嶸
生物技術進展 2021年3期
格日樂其木格, 牛振峰, 董丹, 張濤濤, 崢嶸
1.內蒙古師范大學生命科學與技術學院,呼和浩特 010022;2.北京市農林科學院植物保護環(huán)境保護研究所,北京 100097
微生物種類繁多,與我們的生活密切相關,很多抗生素、免疫抑制劑、除草劑等醫(yī)療和農業(yè)相關的次級代謝產物都來源于微生物。同時,自然界還存在著一些有害微生物,如致病真菌、細菌、病毒等。CRISPR-Cas系統(tǒng)(clustered regularly interspaced short palindromic repeats-crispr associated proteins)的發(fā)展,促進了微生物的基因功能和次級代謝產物的挖掘等方面的研究工作。除此之外,CRISPR-Cas系統(tǒng)也展現(xiàn)出核酸檢測應用的潛力。本文主要從CRISPR-Cas系統(tǒng)的作用機理、分類、微生物基因編輯和核酸檢測中的應用等方面進行綜述,旨在為相關研究提供借鑒。
1 CRISPR-Cas系統(tǒng)概述
CRISPR-Cas系統(tǒng)就是細菌漫長生活史中進化出的一種具有免疫記憶的獲得性免疫防御系統(tǒng),用于抵制外源核酸及噬菌體的入侵[1]。該系統(tǒng)于1987年由日本科學家在大腸埃希菌(Escherichiacoli)中發(fā)現(xiàn)[2]。CRISPR基因座通常由許多間隔的重復序列與非重復的間隔序列(這些序列主要對應于捕獲的病毒和質粒序列片段)組成,并且通常與Cas基因(CRISPR相關)相鄰(圖1)。Cas基因編碼一個大型的異質蛋白質家族,該家族攜帶典型的核酸酶、解旋酶、聚合酶和多核苷酸結合蛋白的功能域[3]。
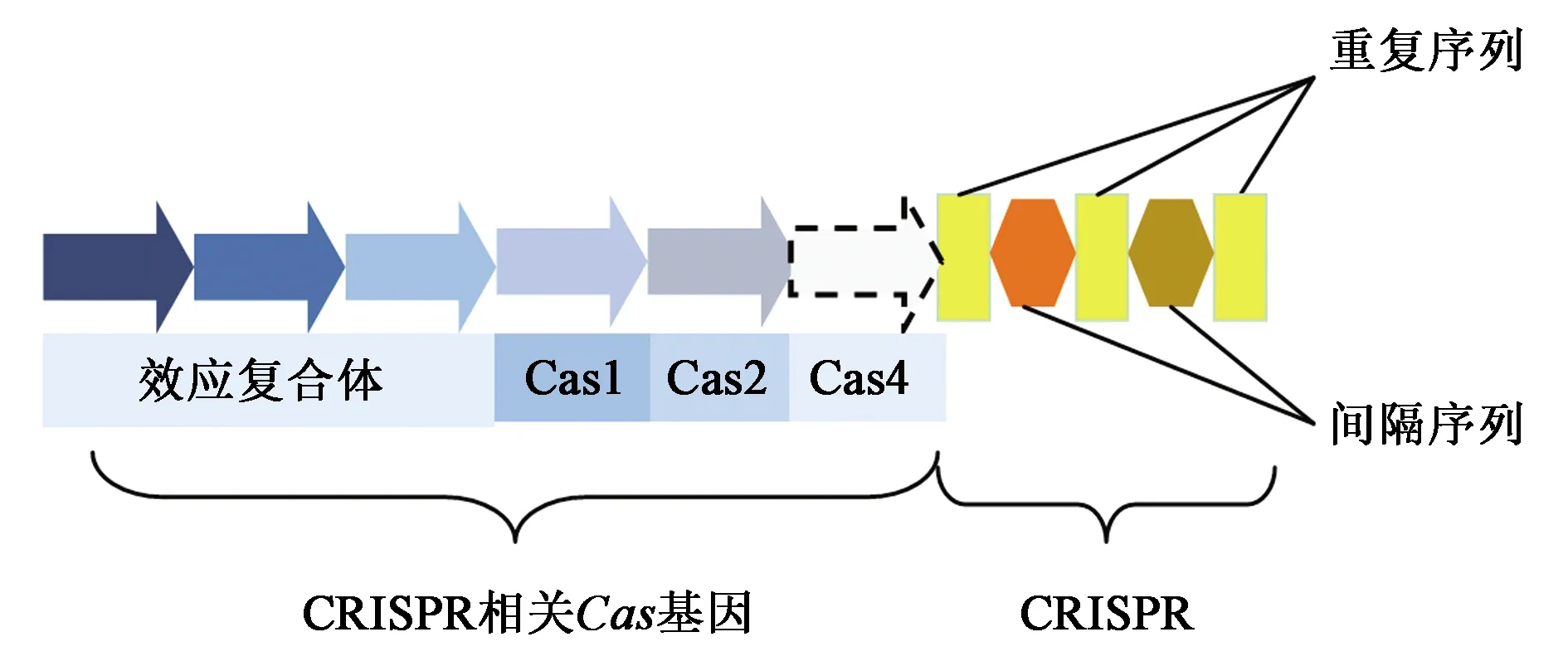
圖1 CRISPR-Cas系統(tǒng)結構
1.1 作用機理
CRISPR-Cas的免疫應答主要包括三個階段:適應、表達和干擾。在適應階段,一種Cas蛋白復合物在識別出特定的前間隔序列鄰近基序(PAM)后結合到目標DNA上,并分裂出目標DNA的一部分,即前間隔序列。再將其整合到自身CRISPR序列中,使其成為新的間隔序列。人們還發(fā)現(xiàn)Cas1和Cas2這兩種高度保守的蛋白在這一階段中起著關鍵作用[2, 4];在表達階段,CRISPR陣列通常被轉錄為一個單一的pre-CRISPR RNA(pre-crRNA),隨后被Cas內切酶(如Cas6)或非Cas蛋白(如RNaseⅢ)加工成更小、更成熟的CRISPR RNA(crRNA)[5];在干擾階段,由成熟的crRNA與單個或多個Cas蛋白形成的復合物識別并裂解侵入核酸[6]。
1.2 分類
2011年Makarova等[7]通過對Cas蛋白序列和結構的比較分析,將CRISPR-Cas系統(tǒng)分為三種類型(Ⅰ,Ⅱ和Ⅲ)以及未分類的系統(tǒng)(U型)。所有這些系統(tǒng)都包含兩個通用基因:Cas1和Cas2。Cas1編碼一種沒有序列特異性的金屬依賴性DNA酶DNAse,可將外源DNA(間隔序列)整合到CRISPR序列中。Cas2編碼一種金屬依賴性核糖核酸內切酶,可能還參與間隔序列獲取階段。三種類型的CRISPR-Cas系統(tǒng)的組成基因組各不相同,并且每種都有其獨特的特征基因來表征[8]。Ⅰ型CRISPR-Cas系統(tǒng)的特征基因為Cas3,其包含HD磷酸水解酶結構域和DExH解旋酶結構域[6],該系統(tǒng)還可分為6種亞型(IA-IF)。Ⅱ型CRISPR-Cas系統(tǒng)可分為2種亞型,其特征基因是Cas9,該基因編碼一種含有HNH 核酸酶結構域和 RuvC 核酸酶結構域蛋白。Ⅰ型系統(tǒng)進行防御時需要幾種蛋白質來完成,而Ⅱ型系統(tǒng)僅需要Cas9核酸酶進行防御[9]。最后一種Ⅲ型CRISPR-Cas系統(tǒng),其特征基因為Cas10(編碼一種結構域與核酸聚合酶和核苷酸環(huán)化酶結構域同源的蛋白),Ⅲ型系統(tǒng)也分為2種亞型(ⅢA,ⅢB),而且兩個亞型靶向不同的核酸[6, 8]。
隨著新類型的CRISPR-Cas系統(tǒng)不斷被發(fā)現(xiàn),在2015年Makarova等再一次對CRISPR-Cas系統(tǒng)進行了分類。根據Cas蛋白在干擾階段的體系結構,將CRISPR-Cas系統(tǒng)分為兩大類:1類(多蛋白效應復合體)和2類(單蛋白效應復合體)系統(tǒng)[10-11]。1類系統(tǒng)(包括類型Ⅰ,Ⅲ和Ⅳ)存在于細菌和古細菌,2類系統(tǒng)(包括類型Ⅱ,Ⅴ和Ⅵ)幾乎完全限于細菌[5, 12]。其中Ⅳ、Ⅴ型和Ⅵ型系統(tǒng)的特征基因分別為Csf1、Cas12和Cas13[4, 10, 13]。目前,CRISPR-Cas系統(tǒng)可分為2大類,6個類型,33個亞型(圖2)。由于2類系統(tǒng)結構簡單,操作方便,是目前應用最廣泛的CRISPR-Cas系統(tǒng)。
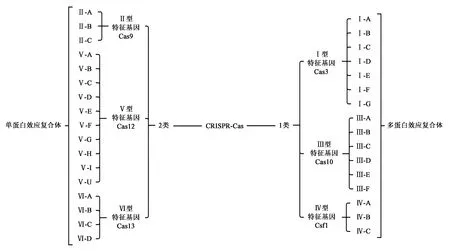
圖2 CRISPR-Cas系統(tǒng)分類
2 CRISPR-Cas系統(tǒng)在微生物基因編輯中的應用
2.1 真菌中CRISPR-Cas基因編輯技術的應用
目前CRISPR-Cas9基因編輯技術已應用于多種真菌基因的功能研究中。早在2013年,DiCarlo等[14]首次利用CRISPR-Cas9基因編輯系統(tǒng)成功在釀酒酵母(Saccharomycescerevisiae)中敲除了細胞膜精氨酸透性酶CAN1基因,使單鏈及雙鏈靶基因斷裂效率分別提高至5和130倍,同源重組率接近100%。2017年劉倩等[15]利用CRISPR/Cas9系統(tǒng)成功地在嗜熱真菌(Myceliophthorathermophila)和嗜熱毀絲霉(Myceliophthoraheterothallica)中進行基因編輯,并對纖維素酶生產途徑中的4個基因(CRE-1、RES-1、GH1-1和ALP-1)同時進行多位點的編輯。利用該系統(tǒng)產生了多個表現(xiàn)出明顯超纖維素酶生產的突變菌株,其分泌的蛋白質和木質纖維素酶活性顯著增加(分別達到野生型菌株的5和13倍)。但仍存在需要復雜的獨立表達盒來靶向多重基因組位點,以及有限數量的可用選擇性標記基因等局限性。2019年,他們利用前期構建的CRISPR-Cas9基因組編輯技術,建立了一種基于V型AsCas12a核酸酶的新型基因組編輯系統(tǒng)對絲狀真菌進行基因編輯[16-17]。Cas12a核酸酶只需要一個Pol Ⅲ啟動子驅動幾個小的crRNAs[18]。基于這個特性CRISPR-Cas12a系統(tǒng)可以同時刪除或插入多個基因。他們還研發(fā)了一種標記回收方法,并將其稱為CRISPR-Cas-assisted marker recycling technology(Camr technology),通過CRISPR-Cas12a/Cas9系統(tǒng)去除標記基因,實現(xiàn)篩選標記的回收和循環(huán)使用。再通過 Camr technology 系統(tǒng)對嗜熱毀絲霉基因組進行了三輪轉化。最終共編輯了纖維素酶分泌途徑的9個關鍵靶基因和兩個選擇性標記基因neo和bar,得到了蛋白質產量和木質纖維素酶活性分別比野生型高9和18.5倍的M9突變體[16-17]。成功解決了絲狀真菌中無法進行多輪編輯的難題,也讓絲狀真菌中的多基因編輯變得更加簡單高效。2020年劉倩等[19]再一次以嗜熱真菌為宿主,結合2A肽策略和CRISPR-Cas9技術異源表達MhglaA和egfp兩個基因,獲得了與野生菌株相比蛋白質產量和淀粉酶活性分別提高約12.0和8.2倍的工程菌。實現(xiàn)了絲狀真菌中多個基因的異源共表達。上述幾種技術提高了嗜熱真菌的工業(yè)價值,也為絲狀真菌的基因編輯提供了新思路。
2.2 細菌中CRISPR-Cas基因編輯技術的應用
2.2.1放線菌中的應用放線菌是革蘭氏陽性細菌,占土壤微生物群的13%~30%,是臨床藥物(如甲酸霉素、法沙霉素、2-烷基-4-羥基喹啉、紅霉素等[20-22])和工業(yè)天然產物的主要來源之一[23]。CRISPR-Cas9基因編輯技術的誕生給放線菌新的次生代謝產物挖掘工作帶來了助力工具。與傳統(tǒng)方法相比,CRISPR-Cas9系統(tǒng)在放線菌中的應用具有效率高、操作方便等優(yōu)點。早在2015年研究人員在鏈霉菌中成功構建了CRISPR-Cas9基因編輯系統(tǒng)并在變鉛青鏈霉菌(Streptomyceslividans)、天藍色鏈霉菌(Streptomycescoelicolor)等多種鏈霉菌中進行了基因編輯[24-26]。2018年Tong等[27]開發(fā)了一個用于放線菌基因組編輯的高效CRISPR-Cas9工具包。該工具包包括sgRNA識別軟件、基因簇敲除系統(tǒng)、基因功能喪失研究系統(tǒng)、生成隨機大小刪除庫的系統(tǒng)和一個用于基因敲除的系統(tǒng)。該團隊成功在StreptomycescoelicolorA3(2)和StreptomycescollinusTu 365中進行基因編輯,證明了該工具包的實用性。雖然CRISPR-Cas9系統(tǒng)已經成功用于放線菌遺傳操作,但由于DNA雙鏈斷裂(DSBs)而引起的基因組不穩(wěn)定和Cas9的(過)表達導致大量不必要的非靶效應等問題仍然存在。
為了解決這些問題,他們又開發(fā)了單核苷酸分辨率基因組編輯系統(tǒng)(CRISPR-base editing system,CRISPR-BEST),CRISPR-BEST與CRISPR-Cas9的區(qū)別在于不需要產生DSBs。針對sgRNA識別的目標序列,CRISPR-cBEST(使用胞苷脫氨)可以有效地將C:G堿基對轉換為T:A堿基對;CRISPR-aBEST(使用腺苷脫胺)可以將A:T堿基對轉換為G:C堿基對(圖3)。此外,該系統(tǒng)通過提供基于Csy4(還稱為I-F型CRISPR相關內切核糖核酸酶Cas6;GenBank登錄號:PHP80843.1)的sgRNA處理系統(tǒng)來支持單個質粒的多重編輯[28-29]。目前CRISPR-BEST已應用于多種鏈霉菌中。然而,正確編輯突變體的篩選和質粒消除的過程仍然是耗時費力的。為了解決這個問題,Wang等[30]開發(fā)了一個基于兩個顯色報告系統(tǒng)(GusA和IdgS)的放線菌CRISPR-Cas9基因組編輯系統(tǒng)。該系統(tǒng)促進了陽性克隆篩選和質粒消除兩個過程,并在放線菌StreptomycescoelicolorM145和Verrucosisporasp.MS100137中完成了不同片段大小的基因敲除,證明這個系統(tǒng)是更快和更有效的基因編輯系統(tǒng)。
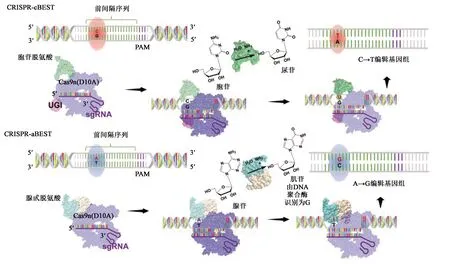
圖3 CRISPR-BEST系統(tǒng)的作用機制[29]
2.2.2其他細菌中的應用CRISPR-Cas系統(tǒng)主要以兩種形式在細菌中進行基因編輯:引入外源CRISPR-Cas系統(tǒng)和利用自身的CRISPR-Cas系統(tǒng)進行基因編輯[31]。Walker等[9]分別用原生Ⅰ-B型系統(tǒng)和異源Ⅱ型GeoCas9系統(tǒng)對嗜熱纖維梭菌(Clostridiumthermocellum)進行了基因編輯。并通過將兩種基因編輯系統(tǒng)與同源重組酶(Exo/Beta,來自Acidithiobacilluscaldus)結合起來,提升同源重組效率,從而提高編輯效率。Ⅰ-B型系統(tǒng)的編輯效率由原來的40%增加到71%,Ⅱ型GeoCas9系統(tǒng)的編輯效率由12.5%增加到94%。Suzuki等[32]構建了一個適用于變形菌門(Proteobacteria)的CRISPR-Cas9基因編輯系統(tǒng)。將來自化膿性鏈球菌(Streptococcuspyogenes)的Cas9基因引入到宿主范圍廣泛的質粒pBBR1MCS-2中,構建質粒pBBR1-Cas9,應用該系統(tǒng)對沙雷菌(Shewanellaoneidensis)MR-1的crp基因進行了編輯。
假單胞菌(Pseudomonas)是一類具有重要生物醫(yī)學、生態(tài)和工業(yè)意義的革蘭氏陰性菌。Chen等[33]報道了利用CRISPR-Cas9和噬菌體λ-Red重組系統(tǒng)構建的基因組編輯系統(tǒng)pCasPA/pACRISPR,該系統(tǒng)可以對銅綠假單胞菌進行有效的遺傳操作。他們還進一步開發(fā)了一個堿基編輯系統(tǒng)pnCasPA-BEC,能夠高效地對多種假單胞菌進行基因失活和點突變,如銅綠假單胞菌(Pseudomonasaeruginosa)、惡臭假單胞菌(Pseudomonasputida)、熒光假單胞菌(Pseudomonasfluorescens)和丁香假單胞菌(Pseudomonassyringae)等。
3 CRISPR-Cas系統(tǒng)在病毒檢測中的應用
病原體是人類的一大威脅,它們在全球范圍內自由傳播。因此,迫切需要提高檢測系統(tǒng)的檢測效率,以便及早發(fā)現(xiàn)。其中分子診斷發(fā)揮了關鍵作用,核酸檢測是主要的分子診斷方法之一[34]。2019年末新冠疫情的爆發(fā)也提醒著人們,核酸檢測技術在疫情防控中起著重要作用。一個高效、便捷的核酸檢測技術可以為疫情防控爭取更多的時間。
因此,迫切需要一種具有高靈敏度和高特異性的核酸檢測方法。此前,張鋒研究團隊和Jennifer Doudna研究團隊分別利用Cas12a和Cas13在目標序列的激活下非特異性切割ssDNA和RNA的特性開發(fā)了SHERLOCK[35](specific high-sensitivity enzymatic reporter unlocking)、DETECTR[36](DNA endonuclease targeted CRISPR trans reporter)兩種核酸檢測技術。這兩種核酸檢測技術可以用來檢測HPV(human papillomavirus,人乳頭瘤病毒)、Zika virus(寨卡病毒)、Dengue virus(登革熱病毒)等病毒[36-37]。同樣也可以檢測COVID-19 virus(新型冠狀病毒),在疫情爆發(fā)之后,張鋒等根據新冠病毒全基因組序列研發(fā)了檢測COVID-19 virus的技術。他們針對COVID-19 virus設計了識別S基因和Orf1ab基因的兩個向導RNA(sgRNA)。這樣,一旦檢測樣本中含有COVID-19 virus,sgRNA就會引導Cas13核酸酶切割這兩個病毒基因,從而激活Cas13核酸酶非特異性切割報告基團并可以在基因試紙上形成視覺可見的條帶[38-40](圖4)。將經過處理的樣品滴在試紙上5 min之內就可以看到結果,在試紙上出現(xiàn)兩個條帶說明結果為陽性[39]。該方法結合重組聚合酶擴增技術(recombinase polymerase amplification, RPA),能夠對樣本中痕量的核酸在恒溫條件下進行大量擴增,因此具有高靈敏度(10~100拷貝·μL-1即可檢測出),從處理樣品到出結果全程不超過1 h且操作簡便,如果該技術實現(xiàn)產品化并推向市場有望彌補現(xiàn)有技術的不足[40]。

圖4 不同輸入濃度橫向流量讀出的示例圖像[39]
4 展望
CRISPR-Cas基因編輯系統(tǒng)的發(fā)展給微生物基因功能研究帶來了助力工具。近幾年CRISPR-Cas基因編輯技術也在不斷更新,從最初的CRISPR-Cas9編輯技術到可以對絲狀真菌同時進行多基因編輯的CRISPR-Cas12a,再到可以對鏈霉菌進行堿基編輯避免了DNA雙鏈斷裂引起的基因組不穩(wěn)定性和Cas9蛋白過表達導致的非靶效應等問題的CRISPR-BEST。CRISPR堿基編輯的誕生也更加豐富了微生物的遺傳操作“工具箱”。
基于CRISPR-Cas系統(tǒng)的核酸檢測技術因靈敏度高、檢測速度快、操作簡單、成本低為醫(yī)學診斷和檢測目標帶來了很大的便利,SHERLOCK已被用于細菌和病毒傳染病病原體的檢測和基因分型,包括區(qū)分單核苷酸變異和尋找抗生素耐藥基因[37]。我們可以利用SHERLOCK區(qū)分新冠病毒的變異毒株,早發(fā)現(xiàn)早隔離,防止新冠疫情大規(guī)模爆發(fā)。并且CRISPR-Cas核酸檢測技術還在不斷優(yōu)化,Melika等[41]認為CRISPR-Cas13不僅可以用來檢測COVID-19 virus,還有治療COVID-19 virus引起的疾病的潛力。新冠疫情為科研人員敲響了警鐘,要不斷地探索發(fā)現(xiàn)才能在危難時刻迅速找到應對之法。相信在不久的將來CRISPR-Cas系統(tǒng)還會應用到更多的領域,給人類帶來更多的驚喜。
猜你喜歡
工業(yè)設計(2022年8期)2022-09-09 07:43:20
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
中國洗滌用品工業(yè)(2019年4期)2019-05-11 09:27:34
