根際促生菌的生防機理及用作生防制劑的潛能
2021-05-27 02:10:26姬彥飛董欣欣楊洪一
中國農(nóng)學(xué)通報 2021年14期
姬彥飛,董欣欣,田 野,張 杰,楊洪一
(東北林業(yè)大學(xué)生命科學(xué)學(xué)院/東北鹽堿植被恢復(fù)與重建教育部重點實驗室,哈爾濱 150040)
0 引言
植物根際促生菌(Plant Growth Promoting Rhizobacteria,PGPR)是指存在于植物根際土壤中的一些可以經(jīng)由各種機制來促進植物生長、提高植株生活力、提高植物對于病害或環(huán)境脅迫的抵抗能力的天然土壤細菌[1]。PGPR已被證實可以通過調(diào)節(jié)植物代謝、改善根際營養(yǎng)環(huán)境、降解土壤污染物等多種途徑促進植物生長,提高植物在不利環(huán)境條件下的生活力,并已作為微生物肥料而在農(nóng)業(yè)生產(chǎn)中加以應(yīng)用。除已被證實的促生長能力外[2],一些PGPR在實際生產(chǎn)與相關(guān)研究中展現(xiàn)出了廣泛的拮抗病原菌的能力,這種能力使其具有抑制病原菌生長、防治植物病害的潛在可能。
植物病害的發(fā)生已成為影響農(nóng)林業(yè)發(fā)展的一個重要難題[3],因而不斷探索新的防治手段就變得尤為重要。植物病害大多為侵染性微生物(真菌、細菌等)通過植株表面?zhèn)诨蚱渌课贿M入植物組織中,經(jīng)由侵染微生物的生命活動或分泌毒素對植物生長發(fā)育或代謝過程造成不利影響,甚至造成植株死亡[4]。利用PGPR對植物病原菌的拮抗活性,可以借由PGPR的生長來對植物病原菌的侵染加以限制或阻滯,從而可以在促進植物生長的同時,提高植物的抗病性能,減少植物病害的發(fā)病率或病害嚴重程度,從而對農(nóng)林業(yè)生產(chǎn)提供助益。
與傳統(tǒng)化學(xué)藥劑防治相比,這種利用特定生物的活動或分泌產(chǎn)物而對另一生物種群的生長產(chǎn)生抑制或殺滅作用的生物防治(biological control,biocontrol)方法具有諸多優(yōu)良特性,如具有作用范圍的廣譜性、持效時間的長效性、不易產(chǎn)生抗藥性的穩(wěn)定性、對生態(tài)環(huán)境的安全性等特點[5],因而具有十分廣泛的應(yīng)用前景。且PGPR還具備一定的促生長機能,能在發(fā)揮生物防治性能之余對植物的生長產(chǎn)生促進,這不僅可以提高植物的產(chǎn)量,對于染病植株的恢復(fù)也尤為重要。
1 PGPR的生防機理
PGPR的生物防治活性依賴于其在植物根際的穩(wěn)定定殖,并經(jīng)由特定生防活性物質(zhì)的產(chǎn)生,及對植物系統(tǒng)抗性的誘導(dǎo)來加以實現(xiàn)。
1.1 PGPR在植物根際的定殖是生防作用發(fā)揮的基礎(chǔ)
植物根際是植物從外界獲取營養(yǎng)的重要部位,根際土壤中的微生物數(shù)量與類群對植物的生長有著極為重要的作用[6]。植物病原菌在根際土壤的大量存在往往導(dǎo)致病害侵染的出現(xiàn),而PGPR的穩(wěn)定定殖則可帶來植物生長參數(shù)與生長活性的顯著提升[7]。PGPR生物防治活性的發(fā)揮也高度依賴其在植物根際土壤的穩(wěn)定定殖。
植物根際土壤營養(yǎng)極其豐富,因而植物根際也是微生物種類與數(shù)量極為繁多的區(qū)域。對根際營養(yǎng)物質(zhì)及生態(tài)位的競爭是PGPR保護植物免受病原菌侵染的基本機制[8]。除PGPR自身的運動能力外,對植物特定產(chǎn)物的趨化作用也是PGPR定殖的一個關(guān)鍵因素。氨基酸及特定糖類都是引起PGPR趨化的常見化學(xué)誘導(dǎo)劑[9]。PGPR的趨化作用經(jīng)由趨化受體蛋白(Methylaccepting Chemotaxis Proteins,MCP)對特定化學(xué)誘導(dǎo)劑的識別而產(chǎn)生。經(jīng)由這一途徑,一些利于植物生長的菌種被迅速、大量地富集在植物根際,植物根部通過分泌有機物供給PGPR生存并利用這些PGPR的活動來對植物的生長加以促進,從而形成一個共生系統(tǒng)[10]。植物分泌趨化物質(zhì)的特異性與PGPR對特定趨化物質(zhì)的選擇性皆由遺傳因素決定[11],這表明植物往往只能對特定的PGPR產(chǎn)生吸引,同時也意味著可利用多種趨化物質(zhì)的PGPR更容易在多種植物的根際穩(wěn)定定殖[12]。除普遍的趨化作用外,特定PGPR表面的特殊結(jié)構(gòu)如胞外基質(zhì)、表面多糖、菌毛等也與其根際定植能力息息相關(guān)[13]。
PGPR通過根際趨化在植物根表大量聚集,隨后形成生物膜并實現(xiàn)穩(wěn)定定殖[14]。這一過程的實現(xiàn)是PGPR產(chǎn)生生防作用的重要基礎(chǔ)。通過PGPR在根際的穩(wěn)定定殖與特定生理活性物質(zhì)在根際圈的積累,土壤病原菌被隔離在植物根際圈之外,PGPR的生防活性得以產(chǎn)生,從而可在根際生態(tài)位進行防御性保持,這對于植物的抗病能力至關(guān)重要(圖1)。
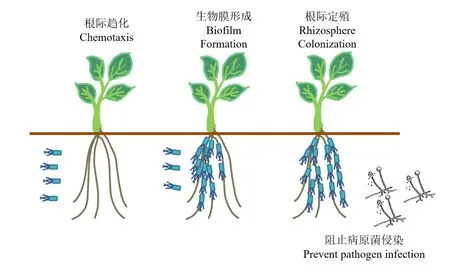
圖1 PGPR在植物根際的定殖
1.2 生防活性物質(zhì)的產(chǎn)生是PGPR生防性能的重要來源
除根際定殖所帶來的生態(tài)位占據(jù)及營養(yǎng)物質(zhì)競爭外,PGPR所分泌產(chǎn)生的一些特定活性物質(zhì)也可對病原菌的生長產(chǎn)生抑制,從而減少病原菌在根際土壤的聚集,或?qū)σ汛嬖诘闹参锊≡鸬綒缱饔谩GPR可分泌產(chǎn)生的生防活性物質(zhì)有鐵載體、降解酶類、抑菌物質(zhì)、解毒因子等多種類型。
1.2.1 鐵載體 土壤中許多營養(yǎng)物質(zhì)溶解度極低,如鐵元素即以溶解度極低的氧化物形式存在。對于PGPR與植物病原菌而言,鐵元素都是其生長所不可或缺的微量元素。許多PGPR可產(chǎn)生一種稱為鐵載體(Siderophore)的小分子有機物,并經(jīng)由這種物質(zhì)鰲合土壤中溶解的鐵離子Fe3+加以吸收。不同菌種產(chǎn)生的鐵載體成分不同,大體分為羧酸型、異羥肟酸型、兒茶酚鹽型等3種[15]。
由PGPR產(chǎn)生的鐵載體可以有效限制土壤中可溶性鐵的供應(yīng),導(dǎo)致土壤中的病原菌產(chǎn)生鐵缺乏,從而停止生長甚而死亡。有研究指出細菌所產(chǎn)生鐵載體對Fe3+的親和力要高于真菌鐵載體,這表明主要為真菌的植物病原菌難以在鐵供給上與PGPR產(chǎn)生有效競爭,此外,許多PGPR被證實具有利用異源鐵載體的能力,這一能力更是加劇了PGPR對土壤鐵供給的限制能力[16]。鐵元素對微生物細胞結(jié)構(gòu)極為重要。鐵載體所造成的病原菌鐵缺乏還會引發(fā)麥角甾醇合成途徑受阻,而麥角甾醇是真菌細胞膜的重要組成成分[17]。因而PGPR可經(jīng)由鐵載體的分泌來抑制病原菌細胞的膜合成,并由此造成其膜功能異常,引起物質(zhì)滲漏,甚而導(dǎo)致病原菌死亡[18]。此外,鐵載體在特定條件下對于其他金屬離子也具有螯合作用,因此對病原菌還可形成其他微量元素的爭奪[19]。
PGPR對土壤微量元素的爭奪可以極大減少病原菌所能獲得的營養(yǎng)供給,從而減輕土壤病原菌對植物的威脅作用,降低植物病害的發(fā)病率,對農(nóng)業(yè)生產(chǎn)產(chǎn)生積極作用。
1.2.2 降解酶類 PGPR還可在生長過程中產(chǎn)生各種降解酶類,這些酶類可以對病原菌的生理結(jié)構(gòu)與生理機能造成損傷與破壞,從而對其侵染能力產(chǎn)生限制,甚至造成病原菌菌體死亡。
解淀粉芽孢桿菌Bacillusamyloliquefaciens可在其生長過程中產(chǎn)生葡聚糖酶與蛋白酶,而葡聚糖與蛋白質(zhì)正是某些侵染性真菌細胞壁的主要成分。土壤中解淀粉芽孢桿菌的活動可對尖孢鐮刀菌Fusarium oxysporum、膠孢炭疽菌Colletotrichumgloeosporioides等病原菌的生長產(chǎn)生抑制,甚至是殺滅作用,從而對某些植物土傳病害起到有效防控的作用[20]。
PGPR還展現(xiàn)出了寄生活性,可以在侵染植物的真菌菌絲上生長,并經(jīng)由分泌胞壁水解酶對菌絲結(jié)構(gòu)造成破壞。李夢婕等[21]在其研究中發(fā)現(xiàn)了石楠銹孢銹菌Aecidiumpourthiaea上的重寄生現(xiàn)象,這些重寄生菌的活動破壞了侵染真菌的孢子產(chǎn)生機制與菌絲結(jié)構(gòu),對于植物銹病的防控有著積極的意義。Dutta[22]通過構(gòu)建轉(zhuǎn)座子文庫對熒光假單胞菌Pseudomonas fluorescens的抗灰霉活性進行了研究,認為幾丁質(zhì)結(jié)合蛋白(chitin-binding protein)等物質(zhì)的合成可能是其生物防治能力的關(guān)鍵。胡小加等[23]利用實時熒光定量PCR技術(shù)分析了菌核病病原菌核盤菌Sclerotinia sclerotiorum拮抗菌的生防機制,發(fā)現(xiàn)該過程中伴隨著一些特定蛋白的產(chǎn)生,這些特定蛋白與核盤菌菌絲結(jié)構(gòu)的破壞有關(guān)。菌核病是一種較難進行化學(xué)防治的植物侵染性真菌病害,而利用PGPR進行生物防治可對該病害起到較好的效果。
1.2.3 抑菌物質(zhì) 一些抑菌性化學(xué)物質(zhì)的產(chǎn)生也是PGPR生防機制的重要一環(huán)。如假單胞菌屬PGPR,被證實具有廣譜的生防活性[24],其生防活性的發(fā)揮與生長過程中CN-的分泌有很大關(guān)聯(lián)[25]。除氰化物的產(chǎn)生外,PGPR還可產(chǎn)生一些特定的代謝物,這些物質(zhì)的分泌可以對病原菌的生長與侵染造成阻滯與抑制,從而在一定程度上避免植物病害對植株生長所造成的不利影響。
揮發(fā)性有機物(Volatile Organic Compounds,VOCs)是一類被廣泛證實的抑菌物質(zhì)。許多PGPR均具有產(chǎn)生醛類、醇類、酯類、羧酸類、酮類、吡嗪類等VOCs的能力[26]。VOCs易于擴散進入病原菌細胞,并對病原菌細胞的菌絲結(jié)構(gòu)與細胞形態(tài)造成損傷,使得病原菌出現(xiàn)細胞腫脹、空泡沉積、結(jié)構(gòu)損傷等異常改變,從而顯著降低其生活力與致病力[27]。VOCs的抑菌作用主要表現(xiàn)在其對病原菌菌絲及分生孢子膜結(jié)構(gòu)的破壞,在該作用下,膜通透性發(fā)生變化,引起胞質(zhì)溶出,并最終導(dǎo)致細胞的死亡。此外VOCs也被證實可在胞內(nèi)引發(fā)活性氧等自由基的產(chǎn)生,從而對病原菌造成進一步損傷[28]。胞內(nèi)活性氧的產(chǎn)生不僅會對細胞膜造成氧化損傷,還會對核內(nèi)DNA分子及相關(guān)蛋白造成不可逆破壞[29]。此外,VOCs還可對植物根系發(fā)育與植株生長產(chǎn)生促進,這有利于植株抗病力的提高與染病后的恢復(fù)[30]。
除揮發(fā)性有機物外,一些PGPR還可在生長過程中產(chǎn)生一些特定的抗真菌代謝物如脂肽(Lipopeptides)、聚酮(Polyketides)等,這些物質(zhì)的存在會抑制植物病原菌的生長[31],對于多種植物病原菌均具有較強的拮抗能力。
脂肽是一類由微生物細胞合成的由脂質(zhì)與氨基酸復(fù)合而成的抑菌物質(zhì)。該物質(zhì)具有廣譜抑菌性能,其抑菌性能的發(fā)揮依賴其對膜結(jié)構(gòu)的破壞[32]。脂肽可經(jīng)由破壞膜脂結(jié)構(gòu)、引發(fā)膜脂過氧化來造成細胞膜滲漏,從而破壞質(zhì)膜通透性并引發(fā)胞質(zhì)溶出,可以顯著抑制病原菌生長、破壞菌體結(jié)構(gòu)并抑制孢子萌發(fā)[33]。脂肽可破壞膜脂中的磷脂雙分子層,在膜上形成孔洞,并經(jīng)由自身表面活性劑的性質(zhì)來加速細胞膜的溶解[34]。
聚酮類化合物是由微生物細胞合成的由低級脂肪酸聚合形成的具有長碳鏈結(jié)構(gòu)的天然抑菌物質(zhì),其抑菌性能的發(fā)揮依賴于其對菌體物質(zhì)合成途徑的抑制[35]。聚酮被病原菌細胞吸收后,可與葡萄糖胺合酶等特異結(jié)合,對病原菌的細胞壁形成造成阻斷,從而對其生長產(chǎn)生抑制。聚酮類物質(zhì)也可抑制病原菌的蛋白質(zhì)合成,這對于病原菌的生長、侵染、毒素分泌等十分不利,甚至可以造成病原菌細胞的死亡[36]。
1.2.4 解毒因子 許多植物病原菌通過特定毒素的產(chǎn)生來對植物正常生長造成抑制或引發(fā)植株結(jié)構(gòu)與功能異常。植物毒素可通過抑制植物酶活性、干擾膜結(jié)構(gòu)正常功能、破壞植物信號傳導(dǎo)等多種途徑來對植物的正常生理功能造成破壞[37]。PGPR可以對植物毒素進行滅活,或通過阻止其產(chǎn)生來保護植物免受植物病原菌傷害[38]。
多粘類芽孢桿菌Paenibacilluspolymyxa可經(jīng)由減輕尖孢鐮刀菌毒素對黃瓜代謝產(chǎn)生的抑制來減輕黃瓜病害[39]。伯克氏菌Burkholderiaambifaria也可對鐮刀菌產(chǎn)生的真菌毒素進行降解,從而避免其侵染對植物造成的損傷[40]。此外伯克氏菌也被報道可以減少核盤菌對油菜的侵染,并可降解黃曲霉素等多種真菌毒素[41]。節(jié)桿菌Arthrobactersp.則可經(jīng)由對酚酸的降解來抑制鐮刀菌的生長,從而減少鐮刀菌毒素的分泌[42]。這對于鐮刀菌所致的各種植物病害的防治具有極為重要的意義。PGPR的降解真菌毒素的作用多借由酯酶、漆酶等解毒蛋白的活動來進行[43],通過該過程,真菌毒素被降解為低毒或無毒的小分子,從而可以避免植物病原菌毒素對植株造成的不利影響。
病原菌的侵染與毒素形成受特定基因的表達與調(diào)控[44],且與菌體間的群體感應(yīng)息息相關(guān)。群體感應(yīng)是微生物的一種胞間通訊,經(jīng)由自身誘導(dǎo)信號等小分子的釋放來進行相互感知[45]。PGPR可通過降解自身誘導(dǎo)信號,從而降低病原菌的群體感應(yīng)能力[46-47],阻止病原菌毒力基因的表達。不動桿菌Acinetobacter lactucae即可通過對信號分子的降解來抑制黃單胞菌對植物的侵染,并減輕其引起的植株枯萎[48],假單胞菌Pseudomonassegetis可降解N-酰基高絲氨酸內(nèi)酯信號分子,從而阻斷植物病原細菌的群體感應(yīng),對于軟腐病等植物病害的防治具有較好的效果[49]。
此外,毒素的分泌高度依賴于病原菌細胞結(jié)構(gòu)與功能的完整性[50],而PGPR可經(jīng)由其他機制對病原菌的細胞結(jié)構(gòu)造成損傷,這也可間接阻止毒素的產(chǎn)生,從而避免植物病原菌對植株造成的損傷[51]。
1.3 植物系統(tǒng)抗性的誘導(dǎo)是PGPR發(fā)揮生防作用的又一手段
系統(tǒng)抗性(Systemic Resistance)是植物在遭受侵染性病害時產(chǎn)生的防御機制,經(jīng)由一些特定基因的激活與代謝途徑的產(chǎn)生,致使植物體內(nèi)一些特定基因表達量或特定物質(zhì)的含量發(fā)生變化[52],整體表現(xiàn)為植物對外界侵染的抵抗性能的提高。根據(jù)產(chǎn)生機制的不同,系統(tǒng)抗性可分為獲得性系統(tǒng)抗性(Systemic Acquired Resistance,SAR)與誘導(dǎo)性系統(tǒng)抗性(Induced Systemic Resistance,ISR)(圖2)。
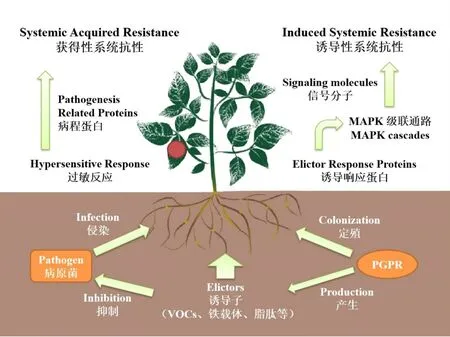
圖2 植物系統(tǒng)抗性的產(chǎn)生機制
SAR是植物在遭受病原微生物侵染或發(fā)病后所產(chǎn)生的防御機制,經(jīng)由特定蛋白對病原菌分泌產(chǎn)生的毒素的感應(yīng)而激活,經(jīng)由水楊酸來傳遞并調(diào)控信號[53],伴隨著病程蛋白的產(chǎn)生及特定酶類的積累。SAR的激活往往伴隨著植物過敏反應(yīng)的出現(xiàn),植株生理代謝發(fā)生顯著改變,植株酚性物質(zhì)積累大幅提升,導(dǎo)致患處胞壁強度提高,誘使感染組織木質(zhì)化壞死[54],從而與健康組織形成隔斷,阻止病原菌的進一步侵染。
與SAR不同,ISR的引發(fā)不需要病害的實際侵染,其顯著特征是不會在植株上產(chǎn)生病變部位,也即誘發(fā)ISR的PGPR不會對植物造成損傷[55]。ISR的產(chǎn)生也是多基因協(xié)調(diào)的結(jié)果,通常可由PGPR代謝產(chǎn)物(如鐵載體、VOCs等)或特定結(jié)構(gòu)(如鞭毛等)誘發(fā),伴隨激素水平改變、抗氧化酶活性的提高或非酶類抗氧化劑含量的提升等。這些變化不但提高了植物對于侵染病害的抵御能力,更提高了植物對于環(huán)境脅迫的承受能力[56]。
近年來分子與組學(xué)研究的發(fā)展使得ISR的產(chǎn)生機制被進一步闡明。Abbasi[57]在其研究中指出,接種PGPR后,植物WRKY70、ERF1等抗性基因被激活,多種抗氧化酶類分泌水平不同程度提高,一些特定基因在植株不同部位差異表達,這些物質(zhì)水平的變化有助于提高植物對于侵染病害應(yīng)對能力。Yuan[58]在其研究中做出了進一步闡述,PGPR接種后,植株中的誘導(dǎo)子響應(yīng)蛋白LRRs、CDPKs等用以傳遞信號,經(jīng)由絲裂原活化蛋白激酶MAPK級聯(lián)通路激活植物激素合成基因,引發(fā)茉莉酸、乙烯含量提升,從而引發(fā)次級通路,系統(tǒng)抗性發(fā)生,相關(guān)應(yīng)激反應(yīng)蛋白與次生代謝產(chǎn)物表達量提升,與清除活性氧自由基相關(guān)的抗氧化酶活性提高,從而導(dǎo)致植物抗逆能力的提升。
系統(tǒng)抗性的誘導(dǎo)是PGPR生防能力的又一重要機制。該機制不依賴于PGPR的直接拮抗,而是通過引發(fā)或增強植物的自然抗病能力,從而對侵染的病原菌產(chǎn)生阻斷或殺滅的效果[59]。ISR的誘導(dǎo)可以時刻激活植物的防御基因,與其他生防機制一同組成PGPR-植物共生體系的抗病屏障。
2 PGPR在生物防治中的應(yīng)用
植物病害對農(nóng)業(yè)生產(chǎn)造成很大影響,植物病害的防范與控制是一個世界性的難題。生防菌株的使用為植物病害防治提供了一個新的道路,不僅可以有效取代化學(xué)藥劑的濫用,還可經(jīng)由PGPR的促生長作用更好的促進農(nóng)林生產(chǎn)[60]。表1列出了近年來使用PGPR進行植物病害防治的研究。

表1 使用PGPR進行生物防治的相關(guān)研究
使用PGPR進行植物病害的生物防治具有化學(xué)藥劑無法取代的諸多特點,如持效期長,PGPR在根際形成定殖后即可長效地發(fā)揮生防能力[65];無污染,菌種及代謝產(chǎn)物均為可被生物降解的物質(zhì),在土壤中不會產(chǎn)生化學(xué)殘留;作用范圍廣泛,對于多種植物病害病原菌具有廣譜的拮抗性能[64],特別是對于化學(xué)藥劑較難防治的病害(如菌核病)或暫無有效化學(xué)藥劑的病害(如甘薯黑斑病[66])具有無可匹敵的防治效果。
近年來隨著組學(xué)技術(shù)的發(fā)展,通過對基因組進行整體分析,可在不依賴培養(yǎng)的情況下評判菌種的生理特性,這為PGPR的發(fā)掘與研究開辟了一條不同以往的道路[67]。借助基因組技術(shù),人們更易于發(fā)掘潛在的PGPR菌株或已有菌株的潛在生防特性[68],這對于菌劑的開發(fā)極為關(guān)鍵。且組學(xué)技術(shù)的應(yīng)用也有利于各PGPR菌株種間關(guān)系的研究,利于人們針對各菌株特性而進行復(fù)合菌劑開發(fā)或探尋最佳施用條件[69]。
目前PGPR在實驗室中與各種植物均進行了相關(guān)實驗?zāi)P偷臉?gòu)建,并取得了較好的成果,但這些實驗室成果仍需經(jīng)過大量的實地實驗才能對其在真實田地中的作用效果加以檢驗[61-62]。有研究將油菜假單胞菌(一種證實的PGPR)與一些植物病害病原菌的基因組進行了比對研究,發(fā)現(xiàn)有益菌與病原菌間存在許多共同的基因特征[70]。這項研究啟示人們植物與微生物間的作用遠比想象的復(fù)雜,它們間相互作用的結(jié)果與植物基因型及生理狀態(tài)、外界脅迫的存在與否、細菌對植物免疫系統(tǒng)的影響能力等均有較大關(guān)聯(lián)。
3 PGPR生防制劑前景展望
PGPR生防能力的發(fā)揮高度依賴于其在植物根際的穩(wěn)定定殖,而PGPR的定殖依賴于對特定化學(xué)物質(zhì)的趨化作用,這意味著某種PGPR菌劑可能無法在所有植物中進行廣泛應(yīng)用。Polcyn[71]的研究也證實,植物品種不同時菌株接種的結(jié)果可能會有較大差異,因而需要進行復(fù)合菌劑的配伍來提高PGPR菌劑的穩(wěn)定性及有效性。PGPR對植物病害的防治效果與菌株數(shù)量、質(zhì)量、生活力息息相關(guān),也即只有定殖的PGPR達到一定規(guī)模才可起到穩(wěn)定的病害防控效果。因而在施用時需保證菌劑中含有足夠的活菌數(shù)。活菌制劑相較于化學(xué)藥劑更難儲存,在維持相近藥效時需要更高的儲藏成本,這在一定程度上制約了PGPR生防菌劑的應(yīng)用,這有待于新劑型的開發(fā)來加以解決[63]。
利用分子生物學(xué)手段對菌株進行改良可能在PGPR生防菌劑的研制中發(fā)揮作用。在明確PGPR的定殖與生防機制后,可利用基因工程技術(shù)定向改造或改良PGPR菌株從而提高其穩(wěn)定性或生防物質(zhì)產(chǎn)量,從而使其在病害防治中發(fā)揮更大的作用[72-74]。對于PGPR生防產(chǎn)物的研究提出了一條新的道路,即不依賴活菌制劑,而是利用PGPR發(fā)酵生產(chǎn)特定生防活性物質(zhì)用以病害防治,也可取得不錯的防治效果[75],并可打破活菌制劑不易保藏難于利用的限制。此外也有針對鐵載體進行的研究,通過改造其結(jié)構(gòu)或利用鐵載體作為藥物負載,也可以在植物病害的防治中發(fā)揮效果[76]。
PGPR菌劑的使用可以在發(fā)揮有效病害防治效果的同時對植物的生長產(chǎn)生促進,不僅可提高作物產(chǎn)量還有利于染病植株的恢復(fù),在農(nóng)林業(yè)生產(chǎn)中具有廣泛的應(yīng)用前景。PGPR菌劑的推廣不僅可減少化學(xué)藥劑的使用,還可減少藥劑生產(chǎn)或施用所造成的環(huán)境污染與威脅。目前仍有一些有待解決的問題制約了PGPR的開發(fā)與應(yīng)用,但隨著相關(guān)機理的進一步闡明與應(yīng)用手段的不斷拓展,可以肯定,PGPR菌劑必將深深改變農(nóng)林生產(chǎn)的格局,開啟生態(tài)種植業(yè)的新篇章。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
