PKD2在骨髓間充質(zhì)干細(xì)胞成脂分化過(guò)程中的表達(dá)變化及作用
2021-05-27 09:57:28單麗英吳曉文師曉麗朱恩東袁海瑞王寶利
山東醫(yī)藥 2021年14期
關(guān)鍵詞:信號(hào)
單麗英,吳曉文,師曉麗,朱恩東,袁海瑞,王寶利
1天津醫(yī)科大學(xué)朱憲彝紀(jì)念醫(yī)院天津市內(nèi)分泌研究所 國(guó)家衛(wèi)健委激素與發(fā)育重點(diǎn)實(shí)驗(yàn)室,天津300134;2天津醫(yī)科大學(xué)基礎(chǔ)醫(yī)學(xué)院
骨髓間充質(zhì)干細(xì)胞(BMSCs)在特定條件誘導(dǎo)下可以向脂肪細(xì)胞、成骨細(xì)胞、軟骨細(xì)胞、神經(jīng)細(xì)胞、成肌細(xì)胞等細(xì)胞分化[1]。BMSCs向脂肪細(xì)胞和成骨細(xì)胞分化的過(guò)程中存在一種平衡關(guān)系,當(dāng)此平衡被打破時(shí),將導(dǎo)致相關(guān)疾病的發(fā)生[2-4]。例如,當(dāng)成脂分化占優(yōu)勢(shì)時(shí)會(huì)削弱BMSCs向成骨細(xì)胞分化的能力,從而引起骨質(zhì)疏松、與年齡相關(guān)的骨丟失等疾病[5-7]。因此,闡明成脂分化和成骨分化之間的平衡關(guān)系對(duì)骨質(zhì)疏松等代謝性疾病的治療有著重要意義。多囊腎致病基因2(PKD2)突變會(huì)導(dǎo)致常染色體顯性遺傳性多囊腎病(ADPKD)的發(fā)生。MESNER等[8]發(fā)現(xiàn)PKD2可以促進(jìn)BMSCs向成骨細(xì)胞分化,但研究未涉及PKD2調(diào)控BMSCs向脂肪細(xì)胞分化的作用。2019年6月—2020年6月,本研究以骨髓間充質(zhì)干細(xì)胞系ST2作為實(shí)驗(yàn)對(duì)象,探討PKD2對(duì)BMSCs成脂分化的作用及可能的調(diào)控機(jī)制,旨在為骨代謝相關(guān)代謝性疾病的治療提供新的思路。
1 材料與方法
1.1 細(xì)胞與主要實(shí)驗(yàn)材料 ST2細(xì)胞為本實(shí)驗(yàn)室保存。無(wú)內(nèi)毒素質(zhì)粒小提試劑盒購(gòu)自CWBIO公司;限制性內(nèi)切酶(NheⅠ、EcoRⅠ)、Opti-MEM、逆轉(zhuǎn)錄試劑盒購(gòu)自Thermo Fisher公司;ClonExpress?Ⅱ重組克隆試劑盒購(gòu)自Vazyme Biotech公司;引物合成、測(cè)序由Sangon Biotech公司完成。α-MEM培養(yǎng)基、胎牛血清(FBS)購(gòu)自Hyclone公司;轉(zhuǎn)染試劑Attrac?tene Transfection Reagent購(gòu)自QIAGEN公司;總RNA提取試劑盒購(gòu)自GeneMark公司;胰島素、地塞米松、3-異丁基-1-甲基黃嘌呤購(gòu)自Sigma公司;吲哚美辛購(gòu)自Cyagen公司;油紅O染液購(gòu)自Sigma公司。倒置顯微鏡購(gòu)自O(shè)lympus公司。
1.2 ST2細(xì)胞成脂分化過(guò)程中PKD2表達(dá)觀察 當(dāng)ST2細(xì)胞匯合度達(dá)100%時(shí),進(jìn)行成脂誘導(dǎo)。使用含有α-MEM培養(yǎng)基(含1%雙抗、5%胎牛血清)、0.5μmol/L地塞米松、0.25 mmol/L IBMX、5 mg/L胰島素、50μmol/L吲哚美辛的成脂誘導(dǎo)培養(yǎng)基誘導(dǎo)72 h,之后更換只含5 mg/L胰島素的α-MEM完全培養(yǎng)基繼續(xù)誘導(dǎo)。采用qRT-PCR法分別檢測(cè)成脂分化誘導(dǎo)0、1、2、3、4、5 d時(shí)細(xì)胞中的PKD2基因。
1.3 PKD2基因過(guò)表達(dá)質(zhì)粒的構(gòu)建 根據(jù)NCBI Ge n e b a n k數(shù)據(jù)庫(kù)中PKD2基因序列設(shè)計(jì)PCR擴(kuò)增引物。PKD2上游引物序列為5'-CTATAGGGAGACCCAAGCTGGCTAGCGCCACCAT?GGTTAACTCCAGACGCGT-3',下游引物序列為5'-CACTGTGCTGGATATCTGCAGAATTCTTAG?GCATGGACGTTGGCAC-3'。以C57BL/6小鼠肝臟cDNA為模板,按照95℃預(yù)變性2 min,95℃變性30 s、56℃退火30 s、72℃延伸150 s共30個(gè)循環(huán),72℃延伸10 min進(jìn)行PCR擴(kuò)增。將擴(kuò)增產(chǎn)物和經(jīng)過(guò)NheⅠ/EcoRⅠ雙酶切的pcDNA3.1用試劑盒純化回收,然后將二者通過(guò)ClonExpress?Ⅱ重組克隆試劑盒進(jìn)行重組反應(yīng)(37℃、30 min),將重組產(chǎn)物轉(zhuǎn)化到DH5α感受態(tài)細(xì)胞中,用LB培養(yǎng)基培養(yǎng)1 h,之后涂板于LB固體培養(yǎng)基(含有100μg/mL氨芐青霉素)上,37℃培養(yǎng)16 h,次日挑取單菌落,搖菌過(guò)夜后提取質(zhì)粒,用NheⅠ/EcoRⅠ雙酶切鑒定,進(jìn)行一代測(cè)序,鑒定正確的菌種即PKD2基因過(guò)表達(dá)質(zhì)粒。
1.4 ST2細(xì)胞的培養(yǎng)及分組處理 轉(zhuǎn)染前1 d,將ST2細(xì)胞接種于24孔細(xì)胞培養(yǎng)板,待細(xì)胞匯合度達(dá)60%~80%時(shí)進(jìn)行轉(zhuǎn)染。將ST2細(xì)胞分為對(duì)照組和實(shí)驗(yàn)組,分別取pcDNA3.1與PKD2過(guò)表達(dá)質(zhì)粒稀釋于Opti-MEM,靜置5 min后加入適量轉(zhuǎn)染試劑Attractene,靜置15~20 min。24孔板中每孔滴加轉(zhuǎn)染混合物50μL,于37℃、5%CO2細(xì)胞培養(yǎng)箱中培養(yǎng)。轉(zhuǎn)染16~18 h后更換新的α-MEM完全培養(yǎng)基。
1.5 ST2細(xì)胞的成脂分化誘導(dǎo)及鑒定 兩組細(xì)胞匯合度達(dá)100%時(shí),進(jìn)行成脂誘導(dǎo)。待脂滴成熟后,采用油紅O染色進(jìn)行鑒定:用4%多聚甲醛室溫固定細(xì)胞15 min;1×PBS洗滌1次;加入適量60%異丙醇,1 min后棄去;最后加入適量油紅O染液,室溫靜置5 min后用蒸餾水沖洗干凈;倒置顯微鏡下觀察細(xì)胞分化情況并拍照;每孔加100%異丙醇200μL萃取油紅O,采用多功能酶標(biāo)儀測(cè)定OD520,表示油紅O的著色程度,定量分析各組間脂滴差異。
1.6 成脂分化相關(guān)基因及蛋白檢測(cè) 采用qRTPCR法檢測(cè)成脂分化相關(guān)基因[過(guò)氧化物酶體增殖物激活受體(PPARγ)、CCAAT增強(qiáng)子結(jié)合蛋白α(C/EBPα)、脂 肪 酸 結(jié) 合 蛋 白4(FABP4)、adip?sin mRNA]。成脂誘導(dǎo)48 h后收集細(xì)胞提取總RNA,用Thermo逆轉(zhuǎn)錄試劑盒逆轉(zhuǎn)成cDNA,然后進(jìn)行qRT-PCR反應(yīng)。反應(yīng)體系:模板cDNA 2μL,上、下游引物(10μmol/L)各0.5μL,2×SYBR Green PCR Master Mix 5μL,去離子水補(bǔ)充至10μL。反應(yīng)條件:95℃預(yù)變性3 min,95℃5 s、56℃15 s、72℃15 s共35個(gè)循環(huán)。以β-actin為內(nèi)參,以2-ΔΔCt表示目的基因相對(duì)表達(dá)量。采用Western blotting法檢測(cè)成脂分化相關(guān)蛋白(PPARγ、C/EBPα、FABP4)。兩組細(xì)胞成脂誘導(dǎo)72 h后用RIPA裂解液裂解,在4℃條件下,12 000 r/min離心10 min,取上清液置于新的離心管中,即為總蛋白。配制10%SDS-PAGE凝膠,將獲得的總蛋白加到凝膠的孔里進(jìn)行蛋白電泳,恒壓60 V至蛋白進(jìn)入分離膠,將電壓調(diào)至120 V至電泳結(jié)束。250 mA恒流將蛋白轉(zhuǎn)印至NC膜,用5%脫脂牛奶室溫封閉2 h,4℃下加一抗過(guò)夜,次日加二抗后曝光,用Image J軟件進(jìn)行灰度值分析,并計(jì)算相關(guān)蛋白與β-actin的灰度比值作為目的蛋白的相對(duì)表達(dá)量。
1.7 Wnt信號(hào)通路相關(guān)蛋白檢測(cè) 將ST細(xì)胞分為對(duì)照組和實(shí)驗(yàn)組,分別轉(zhuǎn)染pcDNA3.1與PKD2過(guò)表達(dá)質(zhì)粒,不進(jìn)行成脂誘導(dǎo)處理,轉(zhuǎn)染后72 h采用Westeren blotting法檢測(cè)Wnt信號(hào)通路相關(guān)蛋白(t-Lrp6、p-Lrp6、t-GSK3β、p-GSK3β、t-β-catenin、活化β-catenin、TCF7L2)。
1.8 統(tǒng)計(jì)學(xué)方法 采用SPSS17.0統(tǒng)計(jì)軟件。計(jì)量資料以±s表示,兩組間比較采用t檢驗(yàn),多組間比較采用單因素方差分析。P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 ST2細(xì)胞成脂分化過(guò)程中PKD2基因表達(dá) 成脂分化誘導(dǎo)后0、1、2、3、4、5 d的ST2細(xì)胞中PKD2基因相對(duì)表達(dá)量分別為1.00±0.19、3.04±0.16、3.02±0.10、2.70±0.88、4.57±1.03、4.45±0.90,成脂分化誘導(dǎo)1 d以后PKD2基因相對(duì)表達(dá)量均顯著高于0 d時(shí),以誘導(dǎo)4 d時(shí)PKD2基因相對(duì)表達(dá)量最高(P均<0.05)。
2.2 PKD2對(duì)ST2細(xì)胞成脂分化的影響 實(shí)驗(yàn)組形成的脂滴數(shù)目顯著減少。實(shí)驗(yàn)組、對(duì)照組油紅定量分別為0.18±0.01、0.42±0.05,實(shí)驗(yàn)組低于對(duì)照組(P<0.05)。
2.3 PKD2對(duì)ST2細(xì)胞成脂分化相關(guān)基因和蛋白表達(dá)的影響 實(shí)驗(yàn)組細(xì)胞中PPARγ、C/EBPα、FABP4、adipsin mRNA表達(dá)低于對(duì)照組;PPARγ、C/EBPα、FABP4蛋白表達(dá)低于對(duì)照組(P均<0.05)。見(jiàn)表1。
表1 實(shí)驗(yàn)組和對(duì)照組細(xì)胞中PPARγ、C/EBPα、FABP4、adipsin mRNA及蛋白表達(dá)比較(±s)

表1 實(shí)驗(yàn)組和對(duì)照組細(xì)胞中PPARγ、C/EBPα、FABP4、adipsin mRNA及蛋白表達(dá)比較(±s)
注:與對(duì)照組相比,*P<0.05。
?
2.4 PKD2對(duì)Wnt信號(hào)通路相關(guān)蛋白表達(dá)的影響實(shí)驗(yàn)組細(xì)胞中p-Lrp6、p-GSK3β、活化β-catenin、TCF7L2蛋白表達(dá)均高于對(duì)照組(P均<0.05)。見(jiàn)表2。
表2 實(shí)驗(yàn)組與對(duì)照組Wnt信號(hào)通路相關(guān)蛋白表達(dá)比較(±s)
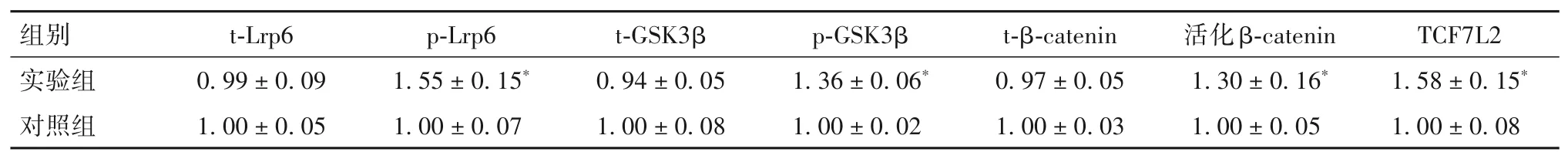
表2 實(shí)驗(yàn)組與對(duì)照組Wnt信號(hào)通路相關(guān)蛋白表達(dá)比較(±s)
注:與對(duì)照組相比,*P<0.05。
?
3 討論
在生理?xiàng)l件下,BMSCs具有向骨、軟骨和脂肪等組織分化的潛能,這對(duì)于維持骨髓內(nèi)骨組織及脂肪組織數(shù)量的平衡具有重要意義。BMSCs向脂肪細(xì)胞或成骨細(xì)胞的分化存在一種動(dòng)態(tài)平衡。在衰老過(guò)程中,BMSCs向脂肪細(xì)胞分化具有偏向性,隨著脂肪組織的積累,骨髓中骨的形成減少。而BMSCs向脂肪細(xì)胞的分化是一個(gè)復(fù)雜、高效的調(diào)控過(guò)程,需要多個(gè)信號(hào)通路、多種轉(zhuǎn)錄因子的參與。研究發(fā)現(xiàn),在基質(zhì)干細(xì)胞脂肪分化過(guò)程中,PPARγ和C/EBPα表達(dá)較高[9],是脂肪生成的重要早期調(diào)節(jié)因子,二者協(xié)同作用可以激活下游FABP4等脂肪生成基因的轉(zhuǎn)錄,從而誘導(dǎo)形成成熟脂肪細(xì)胞[10-11]。
PKD2位于人染色體4q21-23,由15個(gè)外顯子組成,編碼一個(gè)由968個(gè)氨基酸構(gòu)成的6次跨膜蛋白[12]。有研究證實(shí)PKD2基因突變后蛋白的表達(dá)缺失會(huì)導(dǎo)致ADPKD的發(fā)生,主要表現(xiàn)為雙側(cè)腎臟形成多個(gè)大小不等的圓球形液性囊腫[13],造成腎臟功能不可逆性損傷,最終導(dǎo)致腎衰竭[14-16]。PKD2在成人和胎兒的大多數(shù)組織中表達(dá)[17],其在心血管系統(tǒng)中的表達(dá)水平較高,在血管壁平滑肌細(xì)胞和內(nèi)膜細(xì)胞中含量也比較豐富,有助于心血管功能的維持和穩(wěn)定[18]。有研究發(fā)現(xiàn),PKD2作為雙尾基因1的下游基因,可促進(jìn)BMSCs向成骨細(xì)胞分化、增加骨密度[8]。本研究發(fā)現(xiàn),ST2細(xì)胞成脂分化過(guò)程中PKD2的表達(dá)增高,表明PKD2可能在BMSCs向脂肪細(xì)胞分化過(guò)程中發(fā)揮調(diào)控作用。然后我們構(gòu)建了PKD2過(guò)表達(dá)質(zhì)粒,在ST2細(xì)胞中進(jìn)行轉(zhuǎn)染。經(jīng)成脂誘導(dǎo)后,與轉(zhuǎn)染pcDNA3.1質(zhì)粒的對(duì)照組相比,實(shí)驗(yàn)組脂滴的數(shù)目顯著減少,與成脂分化有關(guān)的調(diào)控因子PPARγ、C/EBPα、FABP4、adipsin mRNA和蛋白表達(dá)水平均顯著降低,同時(shí)發(fā)現(xiàn)Wnt/β-catenin信號(hào)通路相關(guān)蛋白表達(dá)水平增加。
Wnt/β-catenin信號(hào)通路參與調(diào)控多種生命發(fā)育過(guò)程,包括細(xì)胞增殖、分化、凋亡、遷移以及成人干細(xì)胞的維持[19-20]。當(dāng)激活信號(hào)傳來(lái),Wnt與Frizzled結(jié)合成復(fù)合物,GSK-3β磷酸化,使得β-catenin不能被降解,從而能夠進(jìn)入胞核與核內(nèi)轉(zhuǎn)錄因子TCF/LEF形成TCF/LEF/β-catenin復(fù)合體,特異性激活下游靶基因,發(fā)揮調(diào)控作用。當(dāng)前多數(shù)學(xué)者認(rèn)為Wnt/β-catenin信號(hào)通路在調(diào)控脂代謝、骨發(fā)育及骨代謝的生理過(guò)程中發(fā)揮重要作用[21]。Wnt/β-catenin信號(hào)通路可以抑制成脂分化[22],促進(jìn)成骨分化。有研究表明,通過(guò)Wnt3a激活的經(jīng)典Wnt通路可以抑制成脂分化過(guò)程中主要轉(zhuǎn)錄因子PPARγ和FABP的表達(dá),Wnt10b能抑制PPARγ和C/EBPα的表達(dá),從而抑制成脂分化[19]。因此,結(jié)合本研究的發(fā)現(xiàn),我們認(rèn)為,PKD2可能通過(guò)激活Wnt/β-catenin信號(hào)通路從而抑制BMSCs向脂肪細(xì)胞的定向分化。但關(guān)于PKD2調(diào)節(jié)Wnt/β-catenin信號(hào)通路的具體途徑仍需要明確。
經(jīng)典的Wnt/β-catenin信號(hào)通路參與了ADPKD的形成[13]。早期研究顯示,在體外,PKD1和PKD2的結(jié)構(gòu)域相互關(guān)聯(lián)[23]。小鼠胚胎成纖維細(xì)胞分泌的Wnt可作為PKD1/PKD2復(fù)合物活化的配體,破壞復(fù)合物的形成;減少PKD1的表達(dá)或降低PKD2的活性,會(huì)抑制Wnt的激活[24-25]。但也有研究表明,在小鼠腎上皮細(xì)胞中特異性敲除PKD2后,通過(guò)增加自分泌Wnt(例如Wnt7a)的產(chǎn)生激活了Wnt/β-catenin信號(hào)轉(zhuǎn)導(dǎo),導(dǎo)致Wnt信號(hào)通路的相關(guān)靶基因表達(dá)上調(diào)[26]。因此,我們推測(cè)在不同細(xì)胞和組織中PKD2對(duì)Wnt信號(hào)通路的異常激活有不同的影響,但仍需要進(jìn)一步驗(yàn)證。
綜上所述,BMSCs成脂分化過(guò)程中PKD2表達(dá)增高;PKD2過(guò)表達(dá)可抑制BMSCs向脂肪細(xì)胞分化,并伴隨Wnt通路相關(guān)蛋白表達(dá)上調(diào)。但細(xì)胞在不同環(huán)境下,PKD2如何調(diào)控Wnt/β-catenin信號(hào)通路還有待進(jìn)一步研究。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國(guó)生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(hào)(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(hào)(2018年2期)2018-04-18 12:18:10
鐵道通信信號(hào)(2016年11期)2016-06-01 12:11:32
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
中國(guó)病理生理雜志(2015年8期)2015-12-21 12:38:06
