莼菜葉片對高溫/低溫脅迫的響應
2021-05-26 05:45:48陳銀花覃章輝沈永琴鐘克王明紅
長江蔬菜 2021年8期
陳銀花 覃章輝 沈永琴 鐘克 王明紅
莼菜(Brasenia schreberiJ.F.Gmel.),是莼菜科(Cabombaceae)莼屬(Brasenia)多年水生草本植物[1],是藥食兩用的珍稀水生蔬菜,已被中國列入國家Ⅰ級重點保護野生植物名錄[1,2]。目前對莼菜的研究主要在栽培管理技術[3,4]、莼 菜 多 糖 提 取[5~7]、莼菜群落[8]和品種資源遺傳多樣性[9]等幾個方面,關于不同生態型莼菜植株對環境因子的生理響應研究較少。
植物都有較適生長溫度范圍,超出這個范圍,會發生溫度脅迫(高溫、低溫),造成生長不良或死亡。持續高溫或低溫脅迫會破壞植物細胞膜結構,使植物體內產生過量的活性氧物質,丙二醛(MDA)含量增加,嚴重影響植株正常的生長和發育[10,11]。有研究表明,植物在高、低溫脅迫下葉片內可溶性蛋白含量顯著增加,以減輕逆境傷害[12];可溶性蛋白質總量與各品系對高溫的耐性呈正相關,耐性強的品系含量高[13]。此外,高溫或低溫脅迫會誘導超氧化物歧酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)等保護酶活性發生變化,活性變化幅度與植物抗逆性密切相關[14,15]。 保護酶活性及MDA含量可作為植物受到溫度脅迫程度和植物對溫度脅迫抵御能力的衡量指標[16]。前期研究結果表明,莼菜有夏季休眠期和冬季休眠期[17],休眠時間長。因此,研究溫度脅迫對不同生態型莼菜植株葉片的生理影響,為篩選抗寒或耐熱莼菜品種和打破莼菜休眠提供理論依據,對莼菜種質資源的保護以及產業發展具有指導意義。

圖1 3類莼菜葉片樣品
1 材料與方法
1.1 試驗材料
試驗所用莼菜葉片于2018年11月10日(莼菜休眠期)采自湖北利川繼昌莼菜基地(30°10'5''N,108°48'22''E;海拔1 100 m左右)。材料來自同一試驗田地,已連作5 a,以每年冬季莼菜爛葉作為自給肥料。樣品包括2種生態型:冬季休眠生態型(DL和WB)和冬季非休眠生態型(ND)。DL是莼菜植株功能葉,隨冬季溫度的持續降低將全部腐爛壞死;WB是莼菜植株在越冬過程中形成的休眠芽葉片,在持續低溫環境下葉片仍然長勢良好;ND是頂端卷曲小葉能正常展開、功能葉長勢良好,并且冬季不會進入休眠的莼菜植株葉片。3類葉片,如圖1所示。
1.2 試驗設計
采取長勢基本一致且都帶有完整健康浮葉的莼菜植株,先用純水將其附帶的雜質清洗干凈,再用種植盆(外口徑21 cm,高12 cm)以水培的形式培養,置于LRH-250-G型光照培養箱(每天12 h光照/12 h黑暗交替,PPFD=121 μmol·m-2·s-1)中,分別進行低溫(5°C)和高溫(35°C)處理,每組處理設3次重復,每個重復15株,以0、2、4、6 d為取樣時間點,每個溫度同一時間點取3株幼苗,每株取長勢相對一致的功能葉片測定其生理生化指標。
1.3 指標測定
可溶性蛋白提取參照宋松泉等[18]的方法,用考馬斯亮藍法測定[19];丙二醛(MDA)含量采用硫代巴比妥酸(TBA)比色法測定[20];抗壞血酸過氧化物酶(APX)活性測定參照宋松泉等[18]的方法;過氧化氫酶(CAT)活性測定參照孫兵[21]、孫祥生[22]的方法;過氧化物歧化酶(SOD)活性測定參照孫兵[21]、Rao等[23]的方法;過氧化物酶(POD)活性測定使用愈創木酚法[21];多酚氧化酶(PPO)活性測定使用鄰苯二酚法[21]。
1.4 數據分析
文中所有數值均表示為:平均值±標準誤。試驗數據用Excel 2010和SPSS 22.0軟件進行統計和分析,用one-way ANOVA進行單因素方差分析和Student-Newman-Keuls檢測進行多重比較(P=0.05)。
2 結果與分析
2.1 高溫脅迫對莼菜休眠和非休眠生態型葉片生理指標的影響
①可溶性蛋白含量的變化 隨高溫脅迫時間延長,休眠生態型植株體內可溶性蛋白含量呈現不同的變化趨勢,DL體內可溶性蛋白質含量呈顯著下降的趨勢,葉片在第5天全部腐爛,WB的可溶性蛋白含量則呈現先降后升的變化趨勢(P<0.05);非休眠生態型植株(ND)在高溫脅迫下呈先降后升的變化趨勢(P<0.05,表1)。
②MDA含量的變化 高溫脅迫下DL的MDA含量顯著高于ND和WB。其隨脅迫時間的延長,休眠生態型植株(DL和WB)的MDA含量呈現先降后升的變化趨勢(P<0.05),非休眠生態型植株的ND的MDA含量則呈現“降—升—降”的變化趨勢(P<0.05,表2)。
③高溫脅迫對莼菜休眠和非休眠生態型葉片SOD、POD、CAT、APX和PPO活性的影響 高溫脅迫下,隨時間的延長,DL、ND和WB的SOD活性均呈顯著下降的變化趨勢(P<0.05),但DL的SOD活性遠高于ND和WB;DL的POD活性隨高溫脅迫時間的延長呈先降后升的變化趨勢(DL在第5天全部腐爛),WB和ND的POD活性呈現“降—升—降”的變化趨勢(P<0.05);DL的CAT活性隨脅迫時間的延長呈顯著上升的變化趨勢(DL在第5天全部腐爛,P<0.05),WB的CAT活性在高溫下脅迫2 d時呈顯著下降(P<0.05),隨后隨脅迫時間延長顯著上升(P<0.05),ND的CAT活性則無顯著性變化(P>0.05);DL和WB的APX活性隨高溫脅迫時間延長無顯著性差異(P>0.05),而ND的APX活性呈顯著上升變化趨勢(P<0.05);DL和ND的PPO活性呈顯著上升的變化趨勢(P<0.05),WB的PPO活性則無顯著性變化(P>0.05,表3)。

表1 高溫脅迫下莼菜休眠和非休眠生態型葉片可溶性蛋白含量(mg/g)隨培養時間的變化
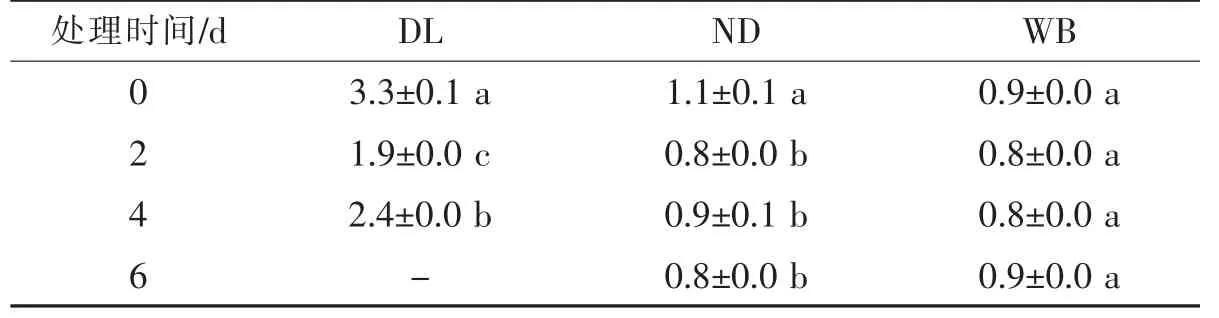
表2 高溫脅迫下莼菜休眠和非休眠生態型葉片MDA含量(nmol/mg)隨培養時間的變化
2.2 低溫脅迫對莼菜休眠和非休眠生態型葉片生理指標的影響
①可溶性蛋白含量的變化 低溫脅迫下,3種莼菜葉片的可溶性蛋白含量隨脅迫時間延長整體呈上升趨勢,其中冬季休眠生態型植株(WB)的可溶性蛋白含量呈顯著上升的變化趨勢(P<0.05,表4)。
②MDA含量的變化 在低溫(5℃)下,隨脅迫時間的延長,冬季休眠生態型植株(DL)的MDA含量呈顯著下降趨勢(P<0.05),且均顯著高于ND和WB的MDA含量。而ND和WB的MDA含量并無顯著性變化(P>0.05,表5)。
③低溫脅迫對莼菜休眠和非休眠生態型葉片SOD、POD、CAT、APX和PPO活性的影響 隨低溫脅迫時間的延長,DL的SOD活性呈現先降后升的變化趨勢(P<0.05),ND的SOD活 性 呈 持 續 下 降的變化趨勢(P<0.05),而WB的SOD活性則無顯著性變化(P>0.05);ND和WB的POD活性在低溫脅迫2 d時均顯著增加,隨后顯著下降(P<0.05),DL的POD活性則與之相反,呈先降后升的變化趨勢(P<0.05);DL、WB和ND的CAT活性均呈顯著上升變化趨勢(P<0.05);DL和WB的APX活性呈現先升后降的變化趨勢(P<0.05),ND的APX活性則呈顯著上升的變化趨勢(P<0.05);ND的PPO活性呈先升后降的變化趨勢(P<0.05),而DL和WB的PPO活性則無顯著性變化(P>0.05,表6)。
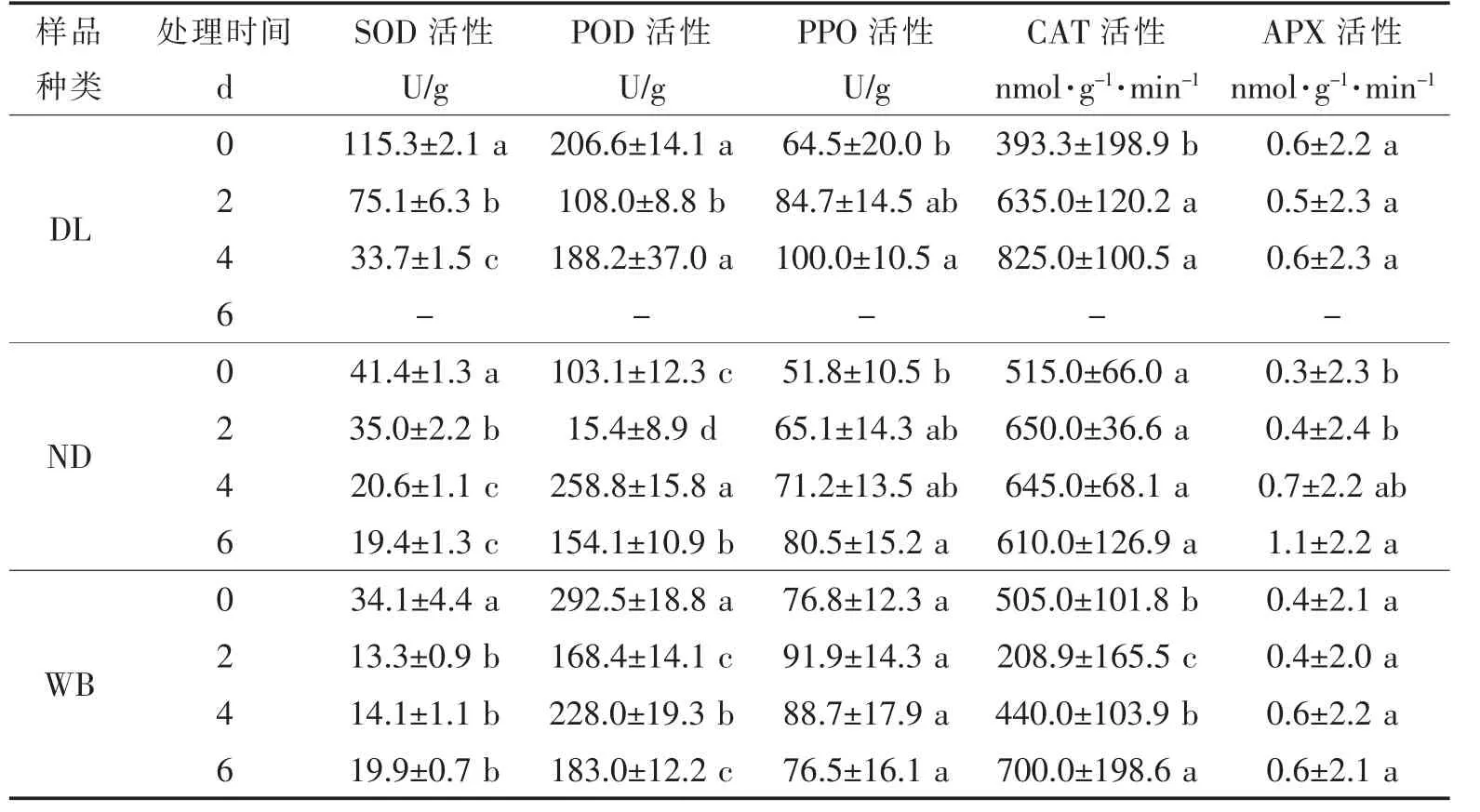
表3 高溫脅迫下休眠和非休眠生態型葉片SOD、POD、CAT、APX和PPO活性隨培養時間的變化
3 討論與結論
植物對逆境的響應與其體內可溶性蛋白含量的變化存在相關性[24],植物體內可溶性蛋白含量的積累可作為植物滲透調節物質和防脫水劑而有利于植物抗寒[25]。本研究中,非休眠生態型植株(ND)和休眠生態型植株(DL、WB)在低溫脅迫下,隨脅迫時間延長,其蛋白質含量整體呈上升趨勢;高溫脅迫下,DL的可溶性蛋白含量隨脅迫時間的增加呈顯著下降的趨勢,這可能是由于溫度過高阻礙了DL植株葉片蛋白質的正常合成,而且植株自身為抵抗逆境其蛋白質會降解,植物自身保護酶系統也會消耗,導致蛋白含量降低。這是DL葉片在高溫脅迫5 d后全部腐爛,而ND和WB植株葉片仍然長勢較好的原因。
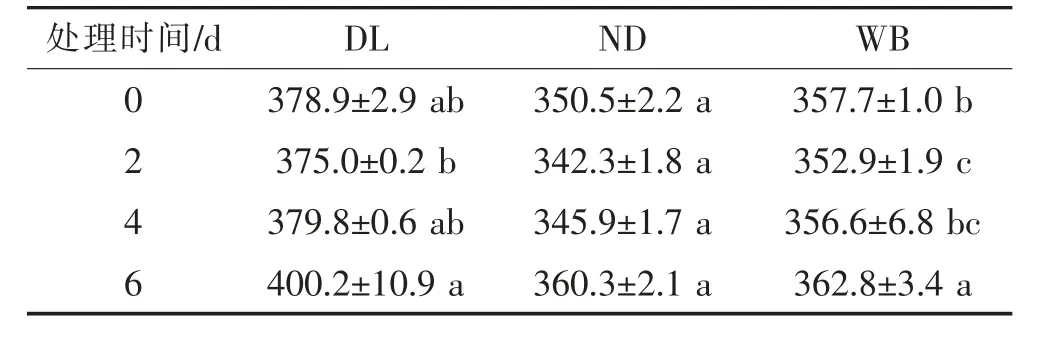
表4 低溫脅迫下莼菜休眠和非休眠生態型葉片可溶性蛋白含量(mg/g)隨培養時間的變化
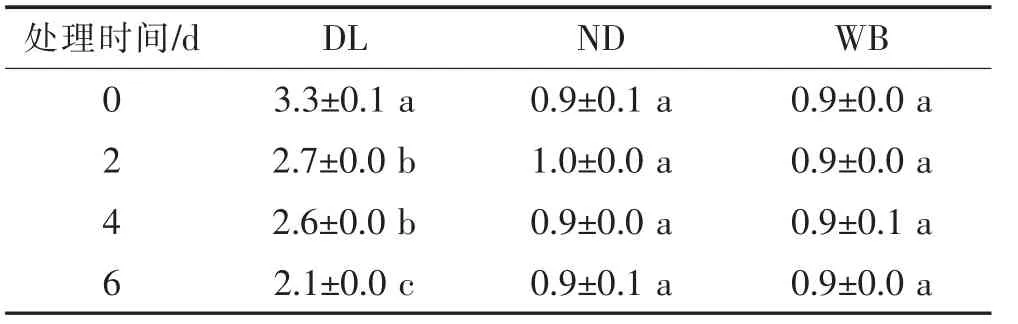
表5 低溫脅迫下莼菜休眠和非休眠生態型葉片MDA含量(nmol/mg)隨培養時間的變化
MDA含量是反映植物細胞受傷害程度以及對逆境的響應[26],耐性強的品種MDA含量較低[27]。植物利用抗氧化酶加強抗氧化作用抑制活性氧的積累,防止自由基的毒害,提高抗性,隨著逆境脅迫的增加,酶活性一般呈上升趨勢,或者呈先上升后下降的趨勢[28,29]。本研究中,隨低溫脅迫時間的延長,DL中的CAT活性顯著上升,MDA含量顯著下降,SOD和POD活性在脅迫2 d后也顯著增加,這說明SOD、POD和CAT能在一定程度上清除了DL中因溫度脅迫產生的過量活性氧物質,減輕植物的膜脂過氧化傷害;而莼菜葉片中APX活性一直較低,并無明顯相關性,這可能是由于APX在DL受低溫脅迫期間只是起著信號傳導作用。綜合分析可知,ND和WB較DL有較強的低溫耐性,ND和WB或為莼菜冬季復壯的理想材料。
高溫脅迫下,隨脅迫時間延長,DL、ND和WB中的SOD、POD活性最終呈下降趨勢;DL的CAT活性在高溫脅迫前期顯著增加,2 d后其活性并無顯著性變化,這說明對高溫脅迫最先做出響應的是SOD。此時莼菜休眠生態型植株葉片(DL)中MDA含量呈現先緩慢降低隨后升高的趨勢,均高于ND和WB 2種生態型植株葉片,這又說明高溫脅迫下DL內MDA含量隨脅迫時間延長而增加與保護酶活性的降低有關。然而隨高溫脅迫時間的延長,ND的CAT活性無顯著性變化,其PPO活性呈顯著上升的變化趨勢,這似乎表明ND在高溫脅迫中,通過PPO來抑制體內的活性氧物質,從而對高溫具有一定的耐性,這與Siddika等[30]的研究結果相似。
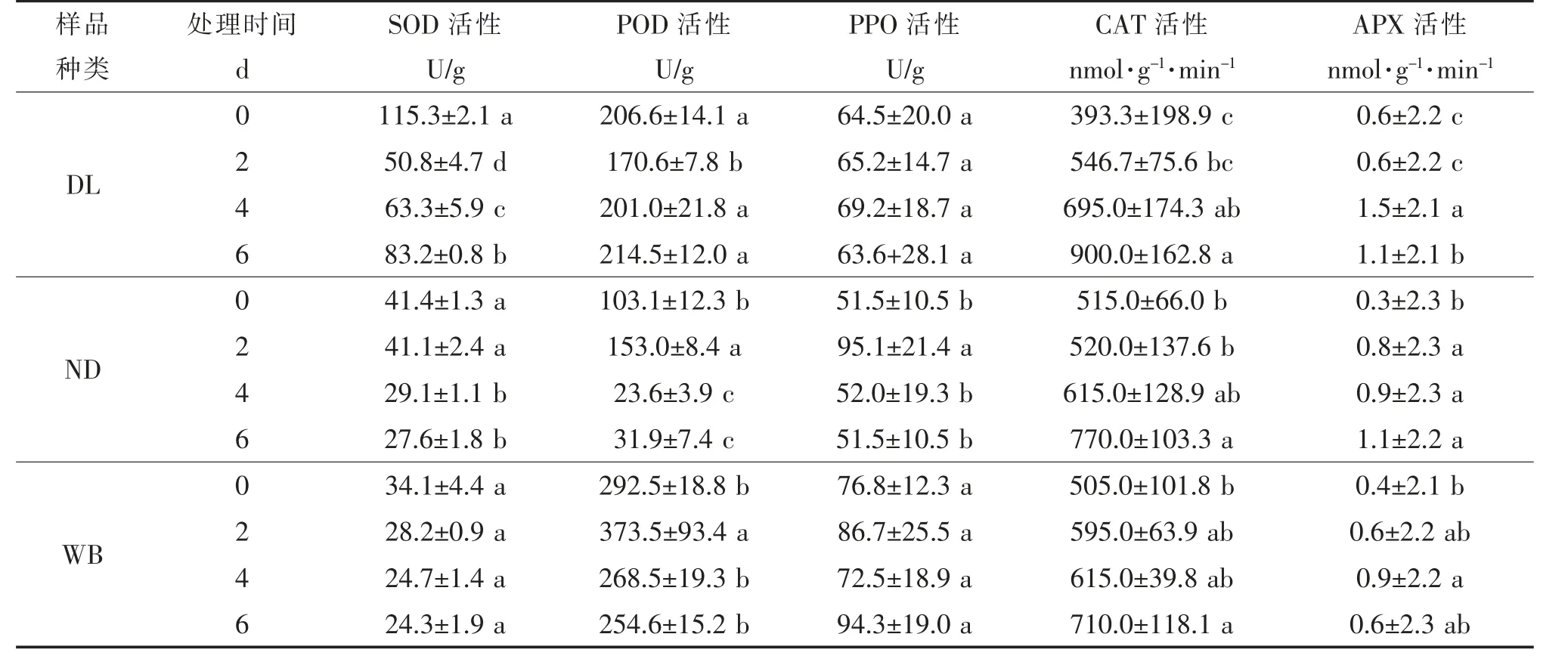
表6 低溫脅迫下休眠和非休眠生態型葉片SOD、POD、CAT、APX和PPO活性隨培養時間的變化
綜上推測,非休眠生態型植株(ND)和冬季休眠生態型的冬芽(WB)對低溫和高溫均具有較強的耐性,或可作為莼菜復壯的理想材料。
