miR-124-3p通過靶向Rab11a緩解創傷性腦損傷引起的氧化應激和炎性反應*
2021-05-26 06:02:36何買定余慧敏楊永花蘇文兵
國際檢驗醫學雜志 2021年10期
何買定,余慧敏,楊 偉,楊永花,蘇文兵
云南省曲靖市第一人民醫院康復醫學科,云南曲靖 655000
創傷性腦損傷(TBI)是一種由于外部原因引起的顱腦損傷,當腦組織挫傷后,大量炎癥因子釋放,氧自由基過載引起海馬神經元細胞氧化應激,進一步促進TBI的發展[1-2],因此,深入探討引起神經元細胞氧化應激和炎性反應的分子機制,對臨床治療TBI極為重要。研究發現,微小RNA(miRNA)在TBI引起的氧化應激和炎性反應中具有重要調控作用[3-5],其中miR-124-3p在TBI后表達下調,上調miR-124-3p能抑制炎癥因子釋放,緩解TBI的發展[6]。腦Ras相關蛋白11a(Rab11a)是一種小分子GTP酶,主要參與調節細胞內吞再循環過程,近年有研究發現Rab11a能促進TBI的發展進程[7],但有關miR-124-3p通過調控Rab11a表達緩解TBI引起的氧化應激和炎性反應的研究,目前鮮有報道。為此,本研究將重點探討miR-124-3p/Rab11a分子軸在TBI引起的氧化應激和炎性反應中的作用,為臨床治療TBI提供一定研究方向。
1 材料與方法
1.1細胞與主要試劑 小鼠海馬神經元(HT22)細胞為BNCC細胞庫產品(中國北納),DMEM培養基和GeneJET RNA 純化試劑盒為Thermo Fisher公司產品(美國);SYBR Green qPCR 主混合液購自Med Chem Express公司(美國);qPCR HifairTM Ⅲ 1st Strand cDNA化學合成超混合液購自上海翊圣生物科技有限公司;PCR引物、miR-124-3p mimic/inhibitor由GenePharma公司構建(中國);pmirGLO熒光素酶靶表達載體及Dual-Luciferase?報告檢測系統購自Promega公司(美國);活性氧(ROS)活性測定試劑盒購于AmyJet Scientific公司(中國);免疫印跡一抗及二抗均購自Abcam公司(英國)。
1.2方法
1.2.1HT22細胞的培養、過氧化氫(H2O2)處理及轉染 將HT22細胞置于含10%胎牛血清的DMEM培養基中,在37 ℃、5% CO2條件的培養箱中培養,取對數生長期細胞為空白對照組(BC組);取10 μmol/L H2O2加入HT22細胞培養基中處理12 h,為氧化應激對照組(OSC組);HT22細胞在H2O2處理后,根據LipofectamineTM2000 檢測試劑盒說明書轉染miR-124-3p mimic、sh-Rab11a和sh-Rab11a+miR-124-3p inhibitor,分別為miR-124-3p mimic組、sh-Rab11a組和sh-Rab11a+miR-124-3p inhibitor組。設3個復孔,轉染48 h后置熒光顯微鏡下觀察細胞的轉染效果。
1.2.2實時熒光定量PCR(RT-qPCR)檢測miR-124-3p和炎癥因子mRNA的表達 采用RNA提取試劑盒提取各組細胞的總RNA,用反轉錄試劑盒對總RNA進行反轉錄,反轉錄反應體系(20 μL):10 μL變性后反應液、4 μL 5×PrimeScript緩沖液、0.5 μL RNase Inhibitor、1.0 μL PrimeScript RTase、4.5 μL dH2O。反轉錄反應條件:30 ℃ 10 min;45 ℃ 60 min,40個循環。根據SYBR Green qPCR主混合液試劑盒說明書檢測mRNA的表達,以U6為檢測miR-124-3p的內參對照,以甘油醛-3-磷酸脫氫酶(GAPDH)為檢測炎癥因子的內參對照。引物序列見表1。RT-qPCR反應體系(20 μL):2.0 μL反轉錄產物、10.0 μL SYBR Green qPCR主混合液、0.4 μL ROX Reference Dye、上下游引物(10 μmol/L)各0.8 μL、6.0 μL dH2O。RT-qPCR反應條件:95 ℃ 5 min;94 ℃變性30 s,60 ℃退火30 s,35個循環。試驗結果用2-ΔΔCt法計算,每個實驗重復3次。

表1 各項靶標的引物
1.2.3Western blot檢測Rab11a及氧化應激相關蛋白的表達 收集各組細胞提取總蛋白,并檢測硒蛋白N1(SEPN1)、谷胱甘肽過氧化物酶1(GPX1)和誘導型一氧化氮合酶(iNOS)蛋白表達水平;SDS-PAGE分離蛋白條帶后,采用半干轉膜法進行轉膜,用5%脫脂奶粉避光室溫孵育1 h,加入一抗(1∶1 000),4 ℃條件下孵育并過夜;棄去一抗,洗膜緩沖液洗滌后,加入二抗(1∶2 000),37 ℃避光孵育1 h后,采用增強化學發光(ECL)液進行顯膜色,凝膠成像儀曝光觀察,拍照記錄,并用Image J軟件進行灰度定量分析。
1.2.4ROS活性檢測 收集處理后的各組細胞(2×105個/mL)重懸于稀釋好的DCFH-DA溶液中,于37 ℃、5% CO2細胞培養箱內培養20 min;培養期間顛倒混勻4~5次;取出細胞后,采用無血清培養基洗滌3次;采用500 μL無血清培養基,加入到96孔板中,并吹打,在熒光酶標儀激發波長488 nm處和發射波長525 nm處檢測熒光強度。
1.2.5雙熒光素酶報告基因試驗驗證miR-124-3p對Rab11a的靶向調控 擴增Rab11a基因3′-UTR片段,將miR-124-3p與Rab11a結合部位的序列及其突變體序列插入到pmirGLO 熒光素酶靶表達載體構建載體,將miR-124-3p mimic與pmirGLO-Rab11a-WT/MUT重組質粒,對照組與LipofectamineTM2000脂質體混合后轉染HEK 293T細胞,轉染48 h后,根據Dual-Luciferase?報告檢測系統檢測熒光素酶活性。
1.3觀察指標 比較各組miR-124-3p表達、SEPN1、iNOS和GPX1蛋白表達,以及caspase 1、白細胞介素1β(IL-1β)、白細胞介素18(IL-18)、腫瘤壞死因子α(TNF-α)和ROS水平。
1.4統計學處理 采用SPSS20.0統計軟件進行數據處理及統計分析,采用GraphPad Prism 7.0軟件進行繪圖。兩組間比較采用t檢驗,多組間比較采用方差分析。以P<0.05為差異有統計學意義。
2 結 果
2.1過表達miR-124-3p對H2O2處理細胞的氧化應激和炎癥因子水平的影響 與BC組比較,OSC組中miR-124-3p表達、SEPN1和GPX1蛋白表達下調,但caspase 1、IL-1β、IL-18、TNF-α、iNOS蛋白表達和ROS水平上調,差異均有統計學意義(P<0.05)。與OSC組比較,miR-124-3p mimic組中miR-124-3p表達上調,SEPN1和GPX1蛋白表達上調,而caspase 1、IL-1β、IL-18、TNF-α、iNOS蛋白表達和ROS水平下調,差異均有統計學意義(P<0.05)。miR-124-3p mimic組與BC組比較,所有檢測指標水平差異均無統計學意義(P>0.05)。見表2和圖1。
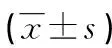
表2 過表達miR-124-3p對H2O2處理HT22細胞的氧化應激和炎癥因子水平的影響
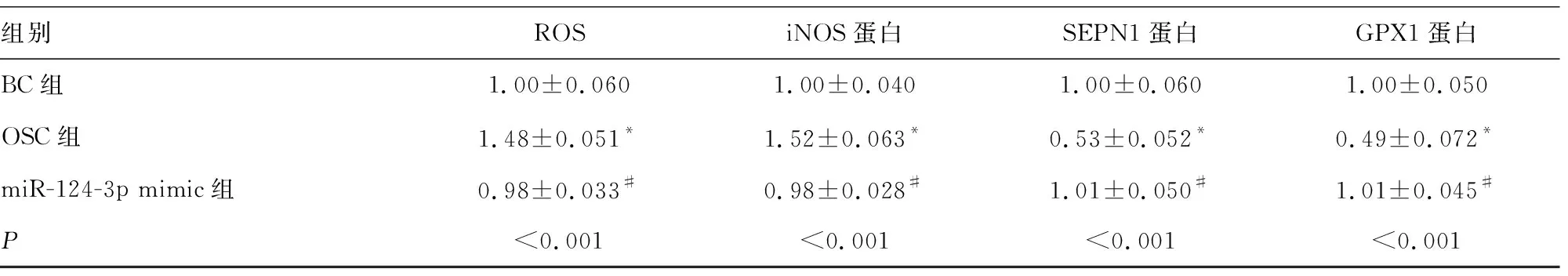
組別ROSiNOS蛋白SEPN1蛋白GPX1蛋白BC組1.00±0.0601.00±0.0401.00±0.0601.00±0.050OSC組1.48±0.051*1.52±0.063*0.53±0.052*0.49±0.072*miR-124-3p mimic組0.98±0.033#0.98±0.028#1.01±0.050#1.01±0.045#P<0.001<0.001<0.001<0.001
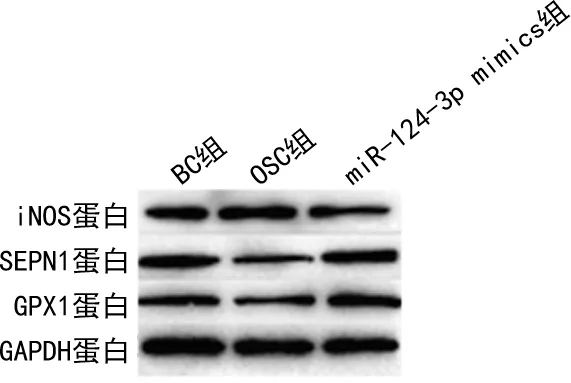
圖1 過表達miR-124-3p抑制H2O2處理HT22細胞產生氧化應激相關蛋白表達
2.2miR-124-3p靶向調控Rab11a的表達 從starBase數據庫預測發現Rab11a是miR-124-3p潛在下游靶基因(圖2A)。與BC組比較,miR-124-3p mimic組中野生型Rab11a的mRNA和蛋白的表達降低,差異有統計學意義(P<0.05),突變型Rab11a中mRNA差異無統計學意義(P>0.05)。見表3和圖2。
表3 相對熒光強度和Rablla表達水平
2.3miR-124-3p通過下調Rab11a抑制H2O2處理HT22細胞氧化應激和炎性反應 與OSC組比較,sh-Rab11a組中Rab11a、iNOS蛋白、ROS、caspase 1、IL-1β、IL-18及TNF-α水平下調,SEPN1和GPX1蛋白表達上調,差異均有統計學意義(P<0.05)。與sh-Rab11a組比較,sh-Rab11a+miR-124-3p inhibitor組中Rab11a、iNOS蛋白、ROS、caspase 1、IL-1β、IL-18表達上調,TNF-α、SEPN1和GPX1蛋白表達下調,差異均有統計學意義(P<0.05)。sh-Rab11a+miR-124-3p inhibitor組與OSC組比較,所有檢測指標差異無統計學意義(P>0.05)。見表4和圖3。
表4 miR-124-3p通過Rab11a對H2O2誘導HT22細胞產生氧化應激和炎性反應的影響
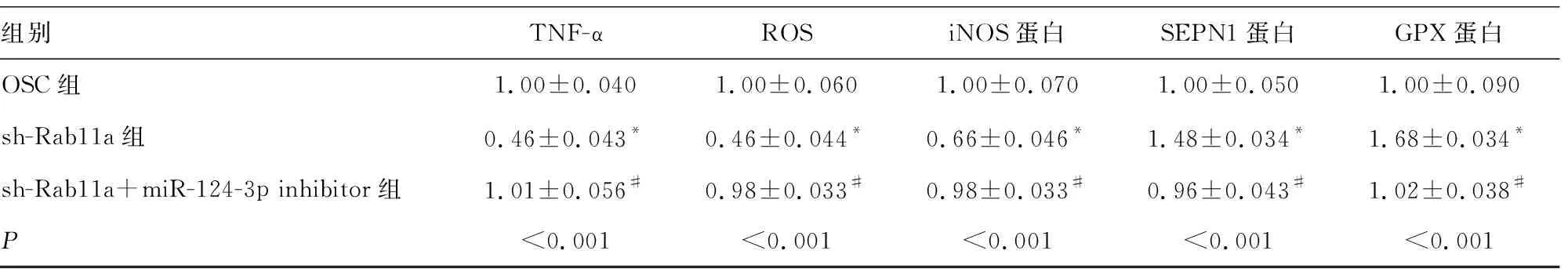
組別TNF-αROSiNOS蛋白SEPN1蛋白GPX蛋白OSC組1.00±0.0401.00±0.0601.00±0.0701.00±0.0501.00±0.090sh-Rab11a組0.46±0.043*0.46±0.044*0.66±0.046*1.48±0.034*1.68±0.034*sh-Rab11a+miR-124-3p inhibitor組1.01±0.056#0.98±0.033#0.98±0.033#0.96±0.043#1.02±0.038#P<0.001<0.001<0.001<0.001<0.001

注:A為miR-124-3p和Rab11a的結合位點;B為Western blot檢測Rab11a蛋白表達水平。
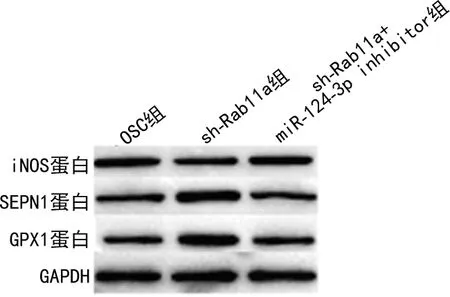
圖3 miR-124-3p通過下調Rab11a抑制H2O2處理HT22細胞產生氧化應激相關蛋白表達
3 討 論
TBI是一種神經外科常見急重癥,其發生率占全身各部位創傷的9%~21%,病死率及致殘率處于全身各部位創傷首位[8-9]。當腦組織挫傷、顱腦結構被破壞后,血腦屏障失去功能,外周血循環中的中性粒細胞、單核巨噬細胞等浸入受損腦組織,啟動一連串分子反應,誘導大量炎癥因子長時間釋放,氧自由基過載并堆積造成活性氧與抗氧化系統之間的平衡失調[10-11],致使海馬神經元細胞等多種細胞發生氧化應激和炎性反應,進一步引起繼發性腦損傷[12-13]。本研究采用H2O2處理HT22細胞,發現caspase 1、IL-1β、IL-18、TNF-α及ROS水平均明顯升高,氧化應激相關蛋白SEPN1和GPX1表達降低,而iNOS蛋白表達水平上調,表明H2O2處理能促進HT22細胞產生氧化應激和炎性反應。此結果與ZHANG等[10]研究一致,且本研究以此為基礎構建TBI細胞模型,進一步探討miR-124-3p/Rab11a分子軸對TBI引起的氧化應激和炎性反應的作用。
非編碼miRNA在近年來的研究中多次被報道參與了原發性和繼發性TBI的發展進程,并通過自身水平變化促進或緩解TBI由原發性向繼發性的惡性轉變[14]。此外,miRNA也被證實在TBI氧化應激和炎性反應中具有重要調節作用,如LV等[15]研究發現上調miR-let-7c-5p能抑制caspase 3、IL-1β等炎癥因子釋放,緩解繼發性TBI發展;miR-124-3p在發生TBI后呈低表達,過表達miR-124-3p能靶向調控下游蛋白表達,抑制神經元發生炎性反應[16-17]。本研究采用H2O2處理HT22細胞構建TBI細胞模型,發現在氧化應激和炎性反應環境中,miR-124-3p明顯低表達,過表達miR-124-3p能恢復由H2O2誘導的炎癥因子caspase 1、IL-1β和IL-18,以及SEPN1、GPX1和iNOS蛋白的特異性表達。本研究結果與VUOKILA等[16]的研究結論一致,此外還進一步證實,過表達miR-124-3p能下調ROS水平,從而緩解HT22細胞所產生的氧化應激。
Rab11a屬于小GTPase超家族的Rab家族,是一種小分子GTP酶,主要參與細胞內吞再循環過程。研究發現,Rab11a參與調節自噬的早、晚期,可促進神經元細胞的自噬[18]和炎癥因子的分泌[19-20],進一步介導繼發性TBI的發展。本研究通過生物信息學預測發現Rab11a是miR-124-3p的潛在靶基因。H2O2處理HT22細胞后,HT22細胞中Rab11a高表達,且Rab11a能夠促進HT22細胞氧化應激和炎性反應,此結果與POEHLER等[18]的研究結論一致。此外本研究進一步證實,miR-124-3p通過靶向下調Rab11a能下調炎癥因子caspase 1、IL-1β、IL-18及ROS水平,上調SEPN1和GPX1蛋白的表達,并下調iNOS蛋白水平,從而緩解HT22細胞的氧化應激和炎性反應。
綜上所述,本研究通過模擬體內TBI產生氧化應激和炎性反應環境,從細胞水平證實,miR-124-3p通過靶向下調Rab11a的表達水平,從而緩解TBI引起的氧化應激和炎性反應。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
西南軍醫(2016年6期)2016-01-23 02:21:19
西南軍醫(2015年2期)2015-01-22 09:09:37
食品科學(2013年15期)2013-03-11 18:25:48
