梨小食心蟲細胞色素P450 CYP6AE123 基因的克隆及表達分析
2021-05-24 01:52:48王彪龍馬瑞燕高玲玲郭艷瓊
山西農業科學 2021年5期
韓 慧,王彪龍,李 恩,胡 軍,馬瑞燕,高玲玲,郭艷瓊
(1.山西農業大學植物保護學院,山西太谷030801;2.澳大利亞聯邦科學與工業研究組織農業與食品部,文布利西澳6913)
梨小食心蟲Grapholita molesta(Busck)屬鱗翅目卷蛾科小食心蟲屬,在全世界廣泛分布,主要為害桃、梨等經濟作物,導致果品產量和質量嚴重下降[1-6]。目前,常用的防治手段為化學防治,但長時間的不合理用藥導致梨小食心蟲對某些殺蟲劑敏感度降低[7]。已有不少研究表明,細胞色素P450 在昆蟲對殺蟲劑的代謝中起著非常重要的作用。
細胞色素P450(Cytochrome P450,CYP)是一類能夠代謝昆蟲內源化合物(如激素等)和外源性化合物(如植物次生物質、殺蟲劑等)的酶系,在昆蟲的解毒、細胞代謝和平衡中起著關鍵作用[8]。CYP 酶的誘導或抑制是昆蟲與藥物相互作用的基礎,它可以使昆蟲產生拒食、趨避、中毒等癥狀。細胞色素P450 具有多個進化支,包括CYP2、CYP3、CYP4 以及線粒體CYP 簇[9],其中CYP3 簇最大,包含CYP6、CYP9 家族在內的多個家族。CYP6 家族屬于昆蟲特有的家族,不少研究證明,該家族基因參與外源物質的代謝。例如飛蝗CYP6 家族參與了外源殺蟲劑的代謝[10],其中CYP6HC1與CYP6HCL1在氨基甲酸酯類和擬除蟲菊酯類殺蟲劑代謝中起關鍵性作用[8];岡比亞按蚊CYP6M2基因可對多種不同類別的殺蟲劑產生耐藥性[11]。此外,目前常用的殺蟲劑有阿維菌素、吡蟲啉以及高效氯氟氰菊酯等。其中,阿維菌素因其殺蟲效果好、對人畜毒性低等優勢被廣泛應用于梨小食心蟲的防治[12]。
本研究從前期構建的轉錄組數據庫進行BLAST 檢索,獲得一個細胞色素P450 基因序列。通過克隆獲得cDNA 全長序列,提交P450 國際命名委員會命名為CYP6AE123,并對其進行生物信息學分析和基因表達模式分析,以明確其不同發育階段和不同組織部位的表達特性,及其對不同濃度阿維菌素的響應,為進一步研究其功能奠定基礎。
1 材料和方法
1.1 試驗材料
本試驗所用梨小食心蟲采自山西省晉中市太谷區西山底桃園,飼養于山西農業大學生物安全與生物防治研究基地人工培養箱中(MGC-450HP),經室內人工飼養50 代以上,光周期15 L∶9 D,相對濕度70%~80%;幼蟲飼喂人工飼料,成蟲飼喂10%蜂蜜水[13],期間未接觸任何殺蟲劑。
收集梨小食心蟲不同發育階段樣品,成蟲3 頭、蛹5 頭、五齡幼蟲5 頭、四齡幼蟲5 頭、三齡幼蟲10 頭、二齡幼蟲30 頭、一齡幼蟲50 頭以及卵100 粒[14]。不同組織選取羽化30 h 之內的健康梨小食心蟲成蟲,分別取頭、前腸、中腸、后腸、脂肪體、馬氏管、精巢、卵巢[15],每個重復取60 頭成蟲。采用阿維菌素LC1(019.82 μg/mL)、LC3(072.90 μg/mL)、LC5(0179.66 μg/mL)分別處理健康的梨小食心蟲成蟲,24 h 后收集存活的成蟲樣品。以上材料均放置于液氮中速凍,存于-80 ℃冰箱中備用,以上樣品均設置3 次生物學重復。
1.2 梨小食心蟲CYP6AE123 克隆
以云杉卷葉蛾CYP6AE83基因為參考序列,對梨小食心蟲轉錄組數據檢索、BLAST 比對,獲得P450 基因的全長cDNA 序列。利用Primer Preimer 5.0 設計特異性引物(表1)。
根據RNAisoTMPlus 試劑盒說明書提取成蟲總RNA,并取1 μg 反轉錄成cDNA,作為PCR 擴增的模板。PCR 反應體系(50 μL):5×TransStartRFast Pfu Buffer 10 μL、2.5 mmol/L dNTPs 4 μL、上下游引物各1 μL、cDNA(1×)2 μL、TransStartRFast Pfu DNA polymerase 1 μL、ddH2O 31 μL。反應程序:95 ℃2 min;95 ℃20 s,60 ℃20 s,72 ℃1 min,40 個循環;72 ℃5 min。反應產物經1%瓊脂糖凝膠電泳后于紫外光下切下目的條帶回收純化。將目的基因連接至pEASY-Blunt 載體中,轉化至大腸桿菌DH5α感受態細胞。將其涂布于LB 固體培養基中,挑取單克隆驗證,并進行LB 液體培養基過夜培養。菌液驗證后送上海生工生物有限公司測序,提交P450 命名委員會進行命名。

表1 引物
1.3 梨小食心蟲CYP6AE123 生物信息學分析
P450基因的開放閱讀框(openreadingframe,ORF)及氨基酸序列使用ORF Finder(http://www.ncbi.nlm.nih.gov/orffinder)預測;采用ExPASy 的在線工具Prot-Param (http://web.expasy.org/protparam/)預測CYP6AE123 蛋白的理化性質;采用SignalP 4.1 Server(http://www.cbs.dtu.-dk/services/SignalP/)分析信號肽;用NPS@_HNNsecondary[15](https://npsa-prabi.ibcp.fr/cgi-bin/secpred_consensus.pl)預測蛋白質的二級結構;采用SWISS-MODEL(https://swissmodel.expasy.org/interactive)進行蛋白質的三級結構同源建模。
根據NCBI BLASTX(https://blast.ncbi.nlm.nih.gov/Blast.cgi)的比對結果找出同源性較高的其他物種的氨基酸序列,在DNAMAN 進行序列比對;采用MEGA 7.0 軟件并運用鄰接法(Neighbor Joining,NJ)構建進化樹,抽樣次數(Bootstrap)設置1 000 次。
1.4 梨小食心蟲CYP6AE123 表達水平分析
提取不同發育階段、不同組織及不同濃度阿維菌素處理后的梨小食心蟲總RNA,并取1 μg 為模板反轉錄為cDNA。以β-actin為內參基因,RT-qPCR檢測CYP6AE123基因的相對表達量。每組樣品均設置3 次技術重復。反應體系(20 μL):SYBR qPCR Master Mix 10 μL、10 μmol/L 的上下游引物(表1)各0.4 μL、稀釋10 倍的cDNA 模板1 μL、去離子水8.2 μL。擴增條件為:94 ℃30 s;94 ℃10 s,50 ℃30 s,72 ℃,30 s,于72 ℃收集熒光,共40 個循環。采用2-ΔΔCT 法[16]計算相對表達量。
1.5 數據分析
使用SPSS 22.0 軟件進行單因素方差分析(one way-ANOVA),采用Tukey 法進行差異顯著性分析,采用Excel 2016 進行數據處理和繪圖。
2 結果與分析
2.1 梨小食心蟲CYP6AE123 的克隆及序列分析
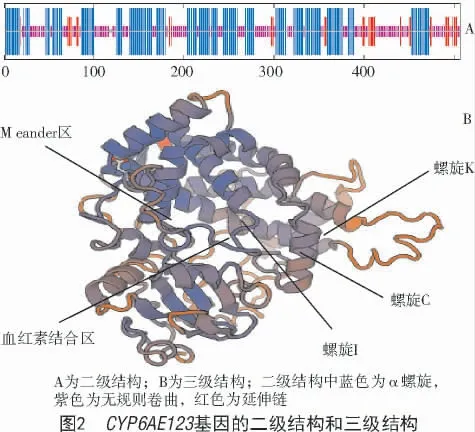
通過檢索、比對、克隆和測序,獲得梨小食心蟲細胞色素P450CYP6AE123的cDNA 全長序列(圖1),提交P450 命名委員會,命名為CYP6AE123。ORF 長度為1 524 bp,編碼507 個氨基酸;蛋白分子質量為58 176.72 u,理論等電點為7.07,不穩定指數為37.50,故該基因編碼的蛋白是穩定的;脂肪系數為89.41,總平均親水性為-0.110,屬疏水性蛋白。二級結構表明,CYP6AE123編碼的蛋白主要由α 螺旋(46.55 %),無規則卷曲(42.80%)以及延伸鏈(8.48 %)組成,三級結構同源建模模擬了P450CYP6AE123的特征序列和空間結構(圖2)。

2.2 梨小食心蟲CYP6AE123 基因的系統發育分析
BLAST 比對結果表明,梨小食心蟲與鱗翅目昆蟲蘋果蠹蛾Cydia pomonella(Cp)、小線角木蠹蛾Streltzoviella insularis(Si)、棉鈴蟲Helicoverpa armigera(Ha) 的氨基酸序列一致性分別高達83.67%、57.39%、52.61%(表2)。

表2 梨小食心蟲CYP6AE123 的BLAST 比對
多序列比對如圖3 所示,梨小食心蟲CYP6AE123具有細胞色素P450 的特征區域,包括螺旋I、螺旋C、螺旋K、Meander 區以及血紅素結合區。

系統發育分析結果顯示(圖4),進化樹中多為各物種的CYP6AE 家族,表明克隆得到的基因屬于CYP6AE 亞家族;且梨小食心蟲CYP6AE123與煙草天蛾CYP6AE129聚在一起,表明二者親緣關系最近,可能具有相似功能。

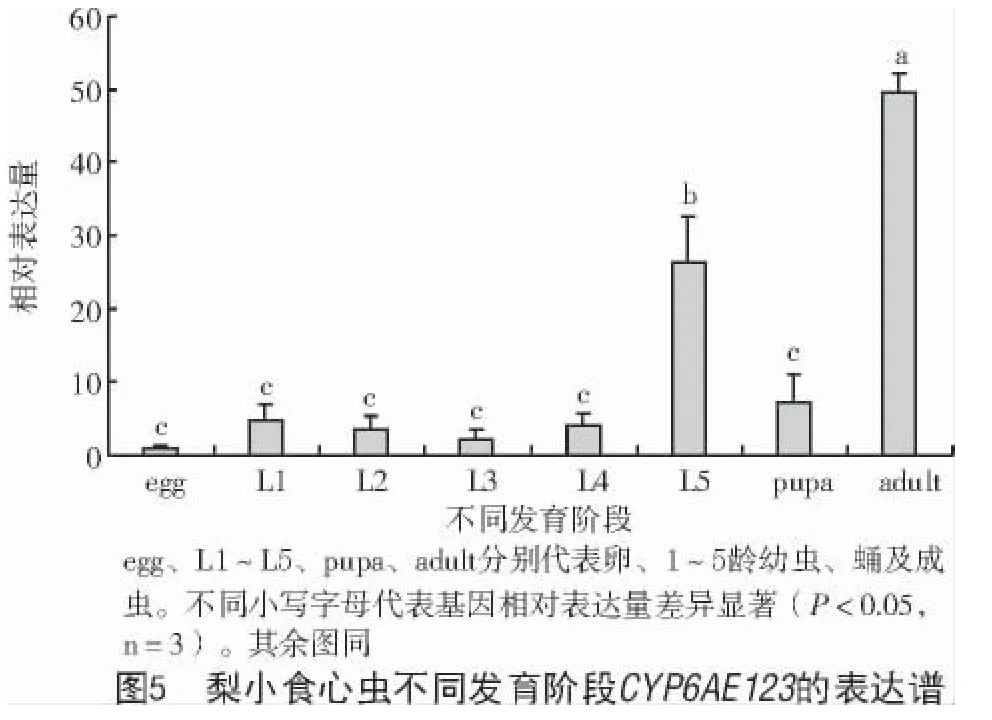
不同發育階段表達模式如圖5 所示,CYP6AE123在梨小食心蟲所有發育階段均有表達,成蟲期和五齡幼蟲相對較高,且差異達顯著水平(P<0.05);其他齡期表達相對較低,且各階段之間無明顯差異。其中,卵期表達量最低,成蟲期表達量是卵期的49.20 倍,五齡幼蟲時期表達量是卵期的26.92 倍。
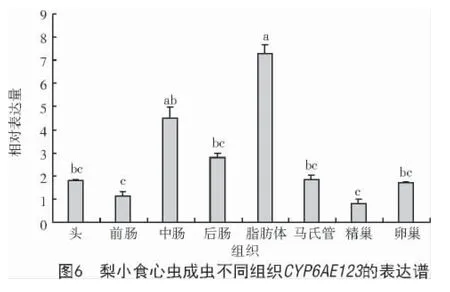
不同組織表達結果表明,CYP6AE123在成蟲的脂肪體中表達量最高,中腸次之;前腸和精巢中表達量最低;脂肪體中CYP6AE123表達量為精巢中的8.81 倍、前腸的6.35 倍;中腸表達量是精巢的5.42 倍、前腸的3.90 倍(圖6)。
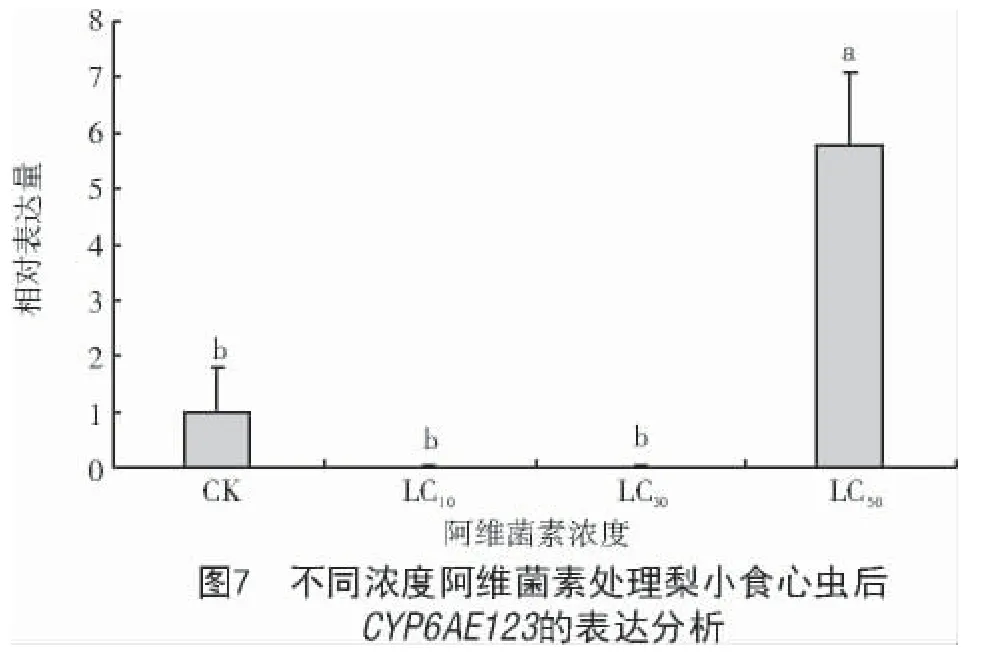
采用不同濃度的阿維菌素對梨小食心蟲成蟲進行處理,結果表明,低濃度阿維菌素(LC10、LC30)處理后,CYP6AE123表達量較CK 降低,但二者間無顯著變化;當濃度增加到LC50(179.66 μg/mL)時,CYP6AE123表達顯著增加,為CK 的5.80 倍,且二者間差異達顯著水平(P<0.05)(圖7)。
3 結論與討論
本研究對梨小食心蟲CYP6AE123的多序列比對結果表明,該基因的氨基酸序列與蘋果蠹蛾、小線角木蠹蛾、棉鈴蟲的氨基酸序列一致性分別高達83.67%、57.39%、52.61%,且均有細胞色素P450 的保守區域,這些保守區域常被用作鑒定P450 的主要特征[17]。
本研究為了進一步研究CYP6AE123的分子特性和生物功能,對不同發育階段與不同組織中CYP6AE123的表達水平進行了表達模式研究,結果顯示,梨小食心蟲CYP6AE123在成蟲期和五齡幼蟲期表達量較高,因為成蟲期是梨小食心蟲為數不多暴露于環境中的時期,也是農民防治的關鍵期[18],在這個發育階段CYP6AE123基因表達量上調,相似的結果在斜紋夜蛾中也有體現[19]。不同組織中基因表達量顯示,CYP6AE123基因在脂肪體中表達量最高,中腸次之,可能是因為脂肪體主要用于儲存能量以及對外源有毒物質的代謝[20],而中腸是昆蟲的消化器官,暗示該基因可能與梨小食心蟲外源物質代謝有關。研究發現,殺蟲劑對昆蟲的誘導表達與基因代謝外源物質密切相關。例如,在氯蟲苯甲酰胺處理甜菜夜蛾后,CYP9 家族基因被誘導,進一步驗證Unigene0035508 基因參與代謝氯蟲苯甲酰胺[21];蘋果黃蚜細胞色素P450 基因AcCYP6CY14表達可增強吡蟲啉抗性[22]。本研究發現,高濃度阿維菌素(179.66 μg/mL)誘導梨小食心蟲CYP6AE123表達,但該基因是否參與梨小食心蟲對阿維菌素的代謝尚不明確,下一步將對其進行RNAi,進一步明確其功能。
本研究克隆了梨小食心蟲CYP6 家族基因CYP6AE123,該基因屬于CYP6AE 亞家族,且包含細胞色素P450 的識別特征;表達分析研究表明,CYP6AE123在成蟲期和脂肪體高表達,且被高濃度的阿維菌素誘導表達,推測其可能參與梨小食心蟲對阿維菌素的代謝,研究結果可為進一步探究梨小食心蟲CYP6AE123基因的功能奠定基礎。
