白藜蘆醇對脂多糖誘導人視網膜色素上皮ARPE-19細胞損傷的保護機制研究
2021-05-23 03:34:40宋慧王英豪
中國中醫眼科雜志 2021年4期
宋慧,王英豪
視網膜炎性反應可導致多種眼科疾病的發生與發展,主要包括葡萄膜炎、糖尿病性視網膜病變和年齡相關性黃斑變性[1-2]。脂多糖(lipopolysaccharides,LPS)在葡萄膜炎和細菌性眼內炎等眼科炎性疾病中可導致視網膜色素上皮(retinal pigment epithelium,RPE)細胞的炎癥反應[3-4],進而誘導線粒體損傷,導致細胞凋亡[5]。因此,尋找能降低RPE細胞炎癥程度、改善線粒體功能損傷的藥物對于眼部疾病的治療至關重要。白藜蘆醇(resveratrol,RES)具有廣泛的藥理活性,在抗炎性反應、抗細胞凋亡、抗腫瘤等方面效果顯著[6-8]。RES屬于一種無毒的天然植物化合物,通過激活線粒體凋亡途徑,抑制視網膜母細胞瘤細胞增殖并誘導細胞凋亡,可作為抗該腫瘤的佐劑[9]。近期一項研究[10]提出,RES可以抑制過氧化應激誘導的人視網膜色素上皮(human retinal pigment epithelial,hRPE)細胞凋亡,其作用機制可能與調節該細胞的周期有關。然而,RES對hRPE細胞損傷保護的分子機制尚不明確。hRPE細胞中豐富的線粒體對于維持細胞穩態至關重要。為了適應環境變化,線粒體可以迅速改變能量的來源以維持細胞代謝能力,最終通過釋放參與細胞死亡途徑的蛋白質而導致細胞凋亡[11]。細胞有一個綜合的機制以抵消線粒體的損害,其中重要的部分是移除功能失調的線粒體,即線粒體自噬或吞噬[12]。早期研究[13]提出,Aldolase/AMPK信號通路參與線粒體的自噬過程,其活化可促使自噬,進而維持細胞內環境穩定。目前為止,RES是否能通過調節線粒體功能改善hRPE細胞損傷尚不清楚。因此,本文從RES對LPS誘導下hRPE ARPE-19細胞活力和凋亡的影響角度進行研究,以期為視網膜疾病的預防和治療尋找新的思路。
1 材料與方法
1.1 材料與儀器
ARPE-19細胞購自武漢巴菲爾公司(來自上海ATCC細胞庫);DMEM培養基購自Invitrogen公司,批號:12100-500;胰蛋白酶及胎牛血清均購自Gibco公司,批號分別為80871和C10 268951;RES購自上海寶曼生物科技有限公司,批號:111535- 200502;SDS-PAGE試劑盒購自武漢谷歌生物有限公司,批號:B1027;細胞計數試劑盒8(cell counting kit 8,CCK8)購自上海工程生物公司,批號:P0012A;Annexin V-FITC/PI細胞凋亡檢測試劑盒購自江蘇凱基生物有限公司,批號:C1062S;Hoechst染色試劑盒購自上海Thermo Fisher Scientific公司,批號:C0003;二氫脫氧嗎啡(compound C,CC)試劑購自上海Selleck公司,批號:ab 120843;LPS購自上海寶曼生物科技有限公司,批號:623311;磷酸緩沖鹽溶液(phosphate buffer saline,PBS)購自上海Thermo Fisher Scientific公司,批號:1209-32;甘油購自江蘇凱基生物有限公司,批號:HY-0904;三磷酸腺苷(adenosine triphosphate,ATP)試劑盒購自江蘇凱基生物有限公司,批號:1209-83;二辛喹酸(bicinchoninic acid,BCA)試劑盒購自上海Thermo Fisher Scientific公司,批號:CY- 201906;四甲基羅達明乙酯購自上海Thermo Fisher Scientific公司,批號:HB-20 180812;Tris緩沖液(tris buffer,TBS)購自江蘇凱基生物有限公司,批號:3980-431;增強化學發光顯色液(enhanced chemiluminecence,ECL)購自上海Thermo Fisher Scientific公司,批號:BP-20 190321;Lipofectamine2000購自上海Invitrogen公司,批號:120M-4017V;兔抗Cleaved caspase-3、p-AMPK、aldolase、α-tubulin和β-actin抗體均購自美國CST公司,批號分別為ab102986、ab190863、ab1339086、ab184736和ab1109376。
1.2 細胞培養
將凍存于液氮中的ARPE-19細胞株進行復蘇,待細胞貼壁融合至適當密度后進行傳代培養。具體操作如下:從培養箱中取出培養瓶至超凈工作臺,無菌PBS清洗2遍,加入適量濃度的胰蛋白酶消化至細胞變圓,然后加入新鮮培養液終止消化,將細胞吹散后轉移至離心管中,1,000 rpm離心5 min,加入含10%胎牛血清的完全培養液吹散分裝于2個培養瓶中,置于5%CO2、37℃恒溫條件下培養。
1.3 細胞分組及處理
將對數生長期細胞分為5組。(1)對照組(NC組):采用正常完全培養液培養。(2)LPS組:采用含100 ng/mL的LPS培養液培養。(3)LPS+RES組(LR組):采用100 ng/mL的LPS和40 μM的RES共同處理。(4)LPS+CC組(LC組):采用100 ng/mL的LPS和10 μM的CC共同處理。(5)LPS+CC+RES組(LCR組):采用100 ng/mL的LPS和10 μM的CC和40 μM的RES共同處理。每組細胞實驗設置5個復孔,均培養24 h。
1.4 細胞活力檢測
把對數生長期細胞消化、重懸、接種于96孔板中,每孔加入200 μL細胞混懸液,邊緣孔用200 μL PBS填滿后培養過夜。細胞貼壁后,按照“1.3”項處理細胞后,取出96孔板,棄上清,每孔加入10 μL CCK8溶液,孵育1.5 h后采用酶標儀檢測450 nm處的光密度(optical delnsity,OD)并計算細胞活力。細胞活力(%)=OD實驗組/OD對照組×100%。
1.5 Hoechst染色
將對數生長期細胞以每孔5×104個接種于6孔板上培養24 h,按照“1.3”項處理細胞后,收集細胞制備成懸液,用醋酸-乙醇固定液固定,0.01 M的PBS漂洗5 min,Hoechst工作液室溫染色15 min,0.01 M的PBS漂洗3次,每次5 min,最后用甘油與PBS比例為1:9的混合液封片,熒光顯微鏡觀察。
1.6 細胞凋亡檢測
收集各組處理后的細胞,懸浮細胞直接轉移至離心管,貼壁則用胰酶消化后轉移至離心管,再用PBS清洗細胞3次,5000 r/min離心6 min,收集細胞,按照Annexin V-FITC/PI試劑盒說明書檢測細胞凋亡情況。
1.7 ATP含量檢測
收集各組處理后的細胞,然后裂解細胞、離心、取上清,采用BCA試劑盒檢測總蛋白濃度。利用ATP試劑盒檢測各組細胞中ATP含量,采用化學發光儀測定吸光度值。
1.8 線粒體膜電壓測定
線粒體膜電壓(mitochondrial membrane potentials,MMP)通過收集各組細胞,采用10 μM四甲基羅達明乙酯染色細胞測定,在37℃條件下反應15min,再利用流式細胞儀檢測熒光強度。
1.9 蛋白免疫印跡實驗
待藥物處理完成后,PBS清洗細胞2次,每孔加入200 μL的裂解液,置于冰上充分裂解30 min,收集細胞裂解液,4 ℃條件下離心12,000 rpm,收集上清液。BCA法檢測上清中總蛋白濃度,每孔上樣40 μg進行電泳分離,恒壓70V,電泳3h。然后在恒流275mA條件下電轉70 min,5%脫脂牛奶封閉1 h后,將目的條帶放入對應的一抗溶液(稀釋比1:1000)中,4℃搖床孵育過夜。TBS洗膜3次,每次10 min,再把條帶放入盛二抗(稀釋比1:3000)的平皿中室溫孵育1 h,TBS洗膜3次,每次10 min。加入4 mL的ECL顯影液顯色3 min,凝膠成像系統曝光。
1.10 統計學方法
采用SPSS 19.0統計軟件進行統計分析,計量資料用均值±標準差()表示,多組比較采用單因素方差分析,各組間兩兩比較用LSD-t檢驗。以P<0.05為差異具有統計學意義。
2 結果
2.1 RES誘導ARPE-19細胞增殖
Hoechst染色發現,各組Hoechst染色陽性ARPE-19細胞數分別為:NC組(798.32±34.54)個、LPS組(108.32±24.21)個、LR組(578.54±46.21)個。3組之間比較,差異具有統計學意義(F=108.412,P=0.000)。與NC組比較,LPS組和LR組陽性細胞數明顯減少(tLPS=36.579、tLR=8.518,均P=0.000);與LPS組比較,LR組陽性細胞數明顯增多(t=20.155,P=0.000),差異均具有統計學意義(圖1)。
2.2 RES降低LPS誘導的ARPE-19細胞凋亡
各組細胞凋亡率分別為:NC組(2.20±1.03)%、LPS組(26.48±3.92)%、LR組(13.83±3.02)%。3組之間比較,差異具有統計學意義(F=43.437,P=0.000)。與NC組比較,LPS組和LR組增高(tLPS=13.395,tLR=8.150,均P=0.000);與LPS組比較,LR組降低(t=5.716,P=0.000),差異均有統計學意義(圖2)。
2.3 RES降低LPS誘導的ARPE-19細胞線粒體損傷
3組細胞ATP比較,差異具有統計學意義(F=46.803,P=0.000)。與NC組比較,LPS組和LR組明顯降低;與LPS組比較,LR組明顯升高(表1、2)。

圖1 各組存活ARPE-19細胞的核染色圖(×200倍)。1A NC組;1B LPS組;1C LR組
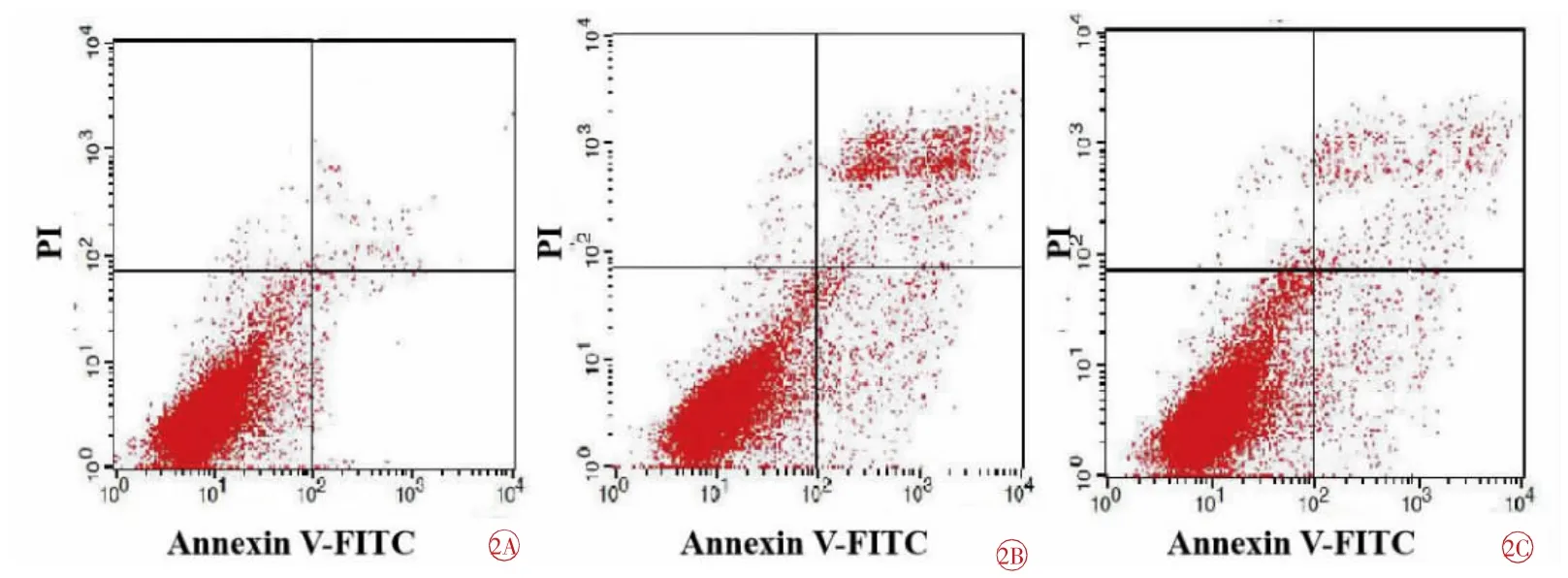
圖2 各組細胞凋亡率的比較。2A NC組;2B LPS組;2C LR組
3組細胞MMP比較,差異具有統計學意義(F=57.815,P=0.000)。與NC組比較,LPS組明顯降低,LR組降低不顯著;與LPS組比較,LR組明顯升高(表1、2)。
表1 各組ARPE-19細胞ATP水平和MMP(,n=5)

表1 各組ARPE-19細胞ATP水平和MMP(,n=5)
注:NC對照組;LPS脂多糖組;LR脂多糖+白藜蘆醇組;ATP三磷酸腺苷;MMP線粒體膜電壓
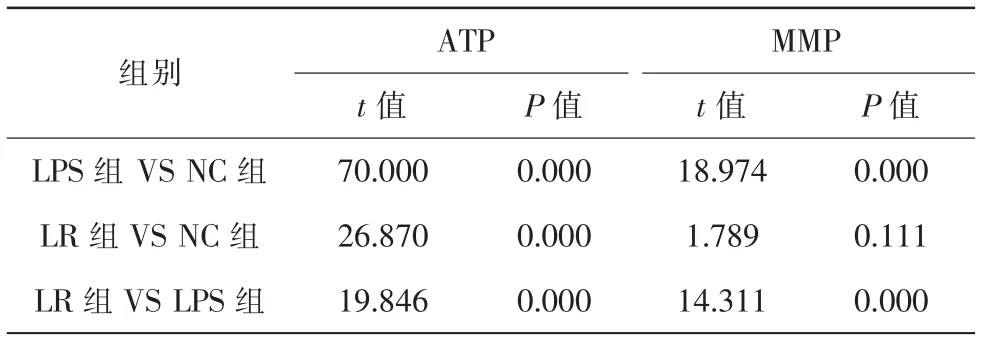
表2 ARPE-19細胞ATP水平和MMP比較統計值
2.4 RES通過AMPK信號通路改善ARPE-19細胞損傷
實驗觀察了CC對p-AMPK蛋白表達的影響。各組細胞p-AMPK蛋白表達水平比較,差異具有統計學意義(F=19.804,P=0.000)。與NC組比較,LPS組、LR組和LC組升高,LCR組升高不明顯。與LPS組比較,LR組升高,LC組和LCR組降低。與LR組比較,LC組和LCR組降低。與LC組比較,LCR組升高不明顯(圖3,表3、4)。
實驗進一步檢測了RES和CC對細胞增殖活力的影響(圖4B)。各組細胞相對活力比較,差異具有統計學意義(F=31.209,P=0.000)。與NC組比較,LPS組、LR組、LC組和LCR組細胞活力降低。與LPS組比較,LR組升高,LC組降低,LCR組升高不明顯。與LR組比較,LC組和LCR組均降低。與LC組比較,LCR組升高(表3、4)。

圖3 p-AMPK蛋白條帶圖
表3 各組ARPE-19細胞p-AMPK表達水平和細胞活力(,n=5)

表3 各組ARPE-19細胞p-AMPK表達水平和細胞活力(,n=5)
注:NC對照組;LPS脂多糖組;LR脂多糖+白藜蘆醇組;LC脂多糖+AMPK抑制劑組;LCR脂多糖+白藜蘆醇和AMPK抑制組;OD光密度
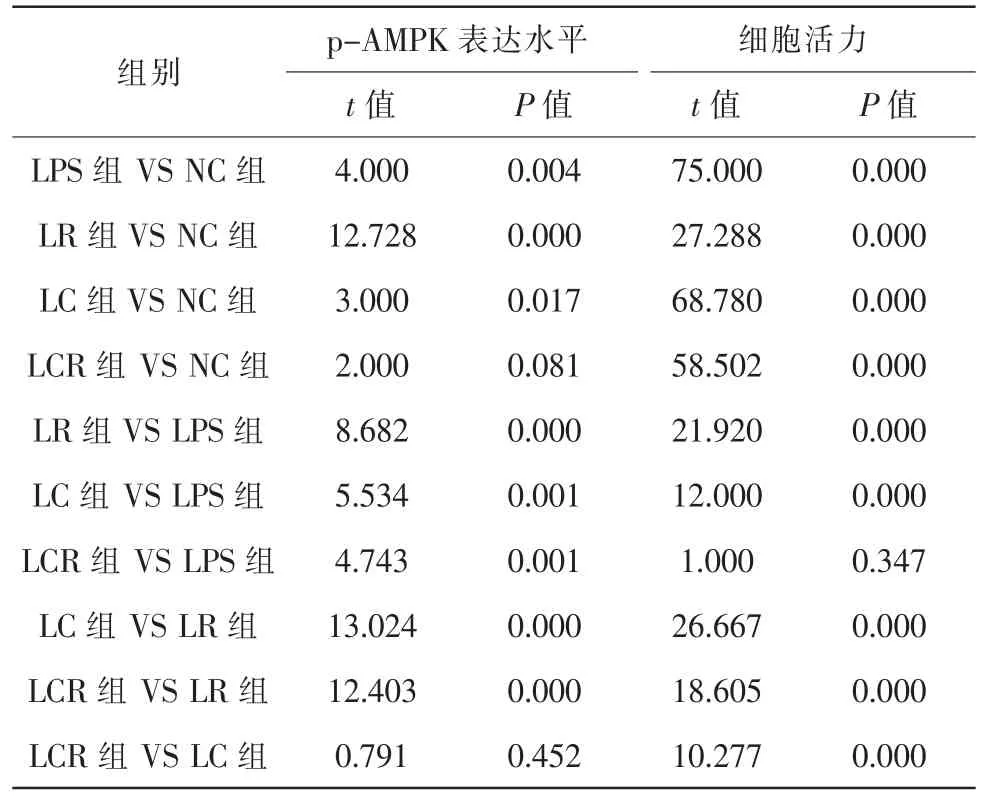
表4 ARPE-19細胞p-AMPK表達水平和細胞活力比較統計值
2.5 RES通過Aldolase激活AMPK信號通路誘導ARPE-19細胞增殖
各組細胞Aldolase蛋白表達水平分別為:NC組(0.09±0.01)、LPS組(0.18±0.02)、LR組(0.41±0.03)。3組之間比較,差異具有統計學意義(F=51.024,P=0.002)。與NC組比較,LPS組和LR組升高(tLPS=9.000,tLR=22.627,均P=0.000)。與LPS組比較,LR組升高(tLR=14.264,P=0.000),差異均有統計學意義(圖4)。
3 討論
視網膜病變的病原學及發病分子機制尚未闡明,既往研究[14]發現,眼部疾病與老化、氧化應激、炎癥反應及線粒體損傷等因素密切相關。隨著年齡增長,人體細胞線粒體功能也隨之減弱,RPE細胞位于脈絡膜毛細血管層與光感受器細胞層之間,容易引起其遭受炎癥因子刺激,進而導致視網膜病變[15]。本文采用LPS干預ARPE-19細胞誘導其線粒體損傷模型,實驗發現細胞ATP水平及MMP均明顯降低,另外還發現線粒體自噬相關蛋白PINK1表達水平也在下降,這說明LPS刺激可誘導ARPE-19細胞線粒體損傷,進而引起細胞活力減弱及凋亡率增高。

圖4 各組細胞Aldolase表達水平比較。4A Aldolase蛋白條帶圖;4B Aldolase相對表達水平
RES是一種屬于非黃酮類多酚化合物的植物保護素,藥理學研究[6-7]表明,其具有顯著的抗炎、抗氧化及抗凋亡的能力。另外,早期研究[16]也證實,RES可通過增強細胞線粒體自噬能力而改善疾病的進展。但有關RES對RPE細胞是否具有抗線粒體損傷活性及其作用機制尚不清楚。細胞主要依賴ATP來實現能量的代謝轉化,隨著細胞內ATP的排出,細胞的自殺機制也將被激活。生理狀態下,細胞中產生的ATP主要來源于線粒體中的氧化磷酸化[17]。在細胞生長過程中線粒體功能主要有線粒體融合、裂變和自噬調節,自噬體可對線粒體進行選擇性的隔離和降解[18]。本研究證實RES能明顯抑制LPS誘導的ARPE-19細胞,增強細胞增殖活力;RES可提升細胞中ATP水平及MMP,促使線粒體自噬能力。
已有研究[19]證實,AMPK信號通路的激活參與了細胞中線粒體自噬過程。RES可以改善線粒體功能異常并且可活化AMPK信號傳導誘導細胞的有益自噬[16]。本文實驗結果也發現,RES可促進ARPE-19細胞中p-AMPK蛋白表達,而AMPK抑制劑CC可阻斷藥物對AMPK磷酸化的激活作用并同時減弱了藥物的保護作用,提示RES對視網膜上皮細胞損傷的保護作用取決于AMPK的活性。前期已有研究[20-21]報道,RES可通過調節AMPK信號通路保護線粒體功能及細胞自噬能力。Aldolase可以作為細胞能量代謝的監視因子,在感知細胞ATP水平降低的同時可激活AMPK信號系統[22]。本實驗發現,RES能誘導ARPE-19細胞中Aldolase的表達。由此說明,RES激活AMPK信號通路和線粒體自噬是通過誘導Aldolase表達來實現的。后續實驗將進一步深入探討RES與Aldolase之間的相互作用,即RES通過何種途徑來調節ARPE-19細胞中Aldolase蛋白表達。
總之,本研究發現RES是通過激活Aldolase/AMPK信號傳導來促進細胞存活,這可能是RPE細胞對炎癥刺激的一種新的適應性反應。
