一水肌酸及共軛亞油酸對大鼠肌纖維相關基因和酶活性的影響及機理
2021-05-19 02:22:18李慧姣侯艷茹趙雅娟白艷蘋趙麗華
食品科學 2021年7期
關鍵詞:劑量
蘇 琳,李慧姣,侯艷茹,趙雅娟,白艷蘋,孫 冰,趙麗華,靳 燁
(內蒙古農業大學食品科學與工程學院,內蒙古 呼和浩特 010018)
肌纖維的生長發育是決定畜肉產量和品質的重要因素,而纖維類型的異質性是動物肌肉組織的重要特征[1]。根據肌球蛋白重鏈(myosin heavy chain,MyHC)的多態性可將肌纖維劃分為4 種肌纖維類型。I型(慢速氧化型肌纖維)肌纖維線粒體數量較多,具有較高的肌紅蛋白含量及有氧代謝酶活力,但ATPase活性較低,故收縮速率慢且持久;IIa型(快速氧化型肌纖維)肌纖維含有一定數量的肌紅蛋白,糖原含量較高,具有有氧代謝和糖酵解代謝兩種供能途徑;IIb型(快速酵解型肌纖維)肌纖維線粒體數量少,糖原含量高,ATPase活性高,糖酵解酶活力高,收縮速率快且短;IIx型(中間型肌纖維)肌纖維的線粒體數量、肌紅蛋白含量、一系列酶活性以及代謝和收縮特性均介于IIa型和IIb型肌纖維之間。因此當氧化型肌纖維比例高時,肌肉的肉色、pH值、大理石紋評分和肌內脂肪含量均較高,肌肉細嫩,保水性能較高,肉質好[2-3]。
一水肌酸(creatine monohydrate,CMH)一般在動物的肝臟或胰腺中由甘氨酸、精氨酸和蛋氨酸合成,機體中約95%的肌酸儲存在骨骼肌中。夏偉光等[4]認為在肉雞飼糧中補充肌酸對肉雞的生長發育尤其是骨骼肌的生長發育具有重要作用;其主要機理是飼糧中添加CMH對肉雞骨骼肌能量代謝的影響可能與一磷酸腺苷活化蛋白激酶(AMP-activated protein kinase,AMPK)的激活有關。而共軛亞油酸(conjugated linoleic acid,CLA)是一類具有共軛雙鍵亞油酸各種構象和位置異構體的總稱。有研究者發現,在妊娠母豬日糧中添加CLA,所產仔豬慢肌纖維比例更高,MyHC I、MyoG和MyoD基因表達量升高,推測CLA影響豬生長發育早期肌肉形成和肌纖維類型[5]。
AMPK被稱為細胞的“能量監測器”,能促進線粒體生物合成及氧化型肌肉表型的代謝變化[6]。目前,AMPK激活的途徑主要有以下3 種:運動激活、激活劑以及特殊飼料誘導激活[7]。營養是保證動物肌肉發育的基礎,如何通過飼糧營養途徑改善肉質是人們較為關注的問題。因此,本實驗用CMH和CLA飼喂大鼠,并對AMPK活力及肌纖維相關基因mRNA相對表達量進行測定,以探尋飼糧添加CMH和CLA對大鼠肌纖維類型組成的影響,及其與AMPK代謝通路的關系。
1 材料與方法
1.1 材料與試劑
CMH、CLA(異構體c9,t11和t10,c12質量分數分別為28.5%和30%) 青島澳海生物有限公司。
RNAiso Plus RNA提取裂解液、PrimeScript? RT Reagent Kit with gDNA Eraser (Perfect Real Time)反轉錄試劑盒、SYBR?Premix ExTaq? II實時熒光定量檢測試劑盒 寶生物工程(大連)有限公司;DNase/RNase-Free無菌水 天根生物技術(北京)有限責任公司;Tris-HCl(分析純) 美國Amresco公司;乳酸脫氫酶(lactic dehydrogenase,LDH)、琥珀酸脫氫酶(succinate dehydrogenase,SDH)、蘋果酸脫氫酶(malate dehydrogenase,MDH)活力檢測試劑盒 南京建成生物工程研究所。
1.2 儀器與設備
CFX96?實時定量聚合酶鏈式反應(polymerase chain reaction,PCR)儀、凝膠成像分析系統 美國Bio-Rad公司;NanoDrop2000核酸蛋白分析儀 美國Thermo Fisher Scientific公司;Veriti96 Well Thermal Cycler PCR儀美國Applied Biosystems公司;水平電泳槽、BG-power 3500型穩壓穩流電泳儀 北京百晶生物技術有限公司;5417R低溫臺式冷凍離心機 德國Eppendorf生物公司;TU-1810紫外-可見分光光度計 北京普析通用儀器有限責任公司;LRH-250生化培養箱 上海一恒科學儀器有限公司;XHF-DY高速分散器 寧波新芝生物科技股份有限公司;PB-10 pH計 北京賽多利斯儀器系統有限公司;ELX800型酶標儀 美國Bio-Tek Instruments公司。
1.3 方法
1.3.1 實驗動物分組與樣品采集
選取符合實驗要求的3 周齡健康雌性Wistar大鼠40 只(購自內蒙古大學動物實驗中心,生產許可證號:SCXK(蒙)2016-0001,使用許可證號:SYXK(蒙)2020-0002),分5 組進行飼養。分組如下:對照組,飼喂基礎飼料;CMH低劑量組,飼喂0.5%(以基礎飼料質量計,下同)CMH+基礎飼料;CMH高劑量組,飼喂1% CMH+基礎飼料;CLA低劑量組,飼喂0.5% CLA+基礎飼料;CLA高劑量組,飼喂1% CLA+基礎飼料。將大鼠分別置于40 個籠中,于相對濕度(55±5)%、溫度(22±2)℃,每12 h明暗交替的環境中,持續飼喂4 周。
大鼠日糧基礎飼料組成配比為:玉米43.15%(質量分數,下同)、小麥粉23.59%、大豆粕18.21%、麩皮11.78%、食鹽0.28%、石粉0.83%、豆油0.49%、酵母0.22%、預混料1.00%。主要營養水平為:粗蛋白質量分數9%、粗脂肪質量分數23%、碳水化合物質量分數68%、代謝能計算值為14.21 MJ/kg。
大鼠飼喂4 周后,麻醉、斷頸處死,3 min內迅速脫皮,取完整的腓腸肌切分成若干份迅速放入標記好的無酶無菌凍存管中,并立即放入液氮速凍,置于-80 ℃冰箱保存備用。
1.3.2 實驗大鼠生長性能的測定
整個實驗周期記錄大鼠體質量增長情況和日攝入量,并計算大鼠日增體質量。
1.3.3 AMPK活力的測定
采用生物素雙抗體夾心酶聯免疫吸附法測定樣品中p-AMPK蛋白含量,反映AMPK磷酸化水平。AMPK活力以p-AMPK蛋白質量濃度表示,即p-AMPK蛋白質量濃度越高,磷酸化程度就越高,相應AMPK活力就越高。
1.3.4 肌纖維相關基因mRNA相對表達量的測定
1.3.4.1 總RNA提取及引物設計
用RNAiso Plus RNA提取裂解液對肌肉組織的總RNA進行提取。利用核酸蛋白分析儀檢測所提取總RNA濃度和純度(A260nm/A280nm);采用質量分數1%的瓊脂糖凝膠電泳檢測完整性。將提取的RNA質量濃度稀釋為500 ng/μL,按照反轉錄試劑盒說明書進行反轉錄操作,合成cDNA,置于-20 ℃冰箱備用。實驗所需大鼠引物序列參照文獻[8-9],由艾博生物技術有限公司設計合成。具體序列如表1所示。

表1 實時定量PCR引物Table 1 Primer sequences used for real-time polymerase chain reaction
1.3.4.2 實時熒光定量PCR檢測
采用嵌合熒光法,以合成的cDNA為模板,使用實時熒光定量試劑盒進行擴增。管家基因和目的基因分別做3 個平行和2 個陰性對照。反應體系:SYBR?Premix ExTaq? II 12.5 μL;上下游引物各1.0 μL;DNA模板(10 ng/μL)2.0 μL;dH2O 8.5 μL;共25.0 μL。實時熒光定量PCR條件:95 ℃預變性30 s;95 ℃變性5 s,60 ℃退火30 s,72 ℃延伸30 s,共35 個循環;72 ℃延伸10 min。產物保存于4 ℃冰箱。采用2-ΔΔCt法計算目的基因相對表達量,計算公式[10]如下。

1.3.5 相關代謝酶活力的測定
LDH、SDH、MDH活力的測定分別按照相應檢測試劑盒說明書步驟進行操作,結果以蛋白質量計。
1.4 數據統計與分析
利用SPSS 20.0軟件進行數據統計分析,結果均以平均值±標準差表示。用單因素方差分析法進行顯著性差異分析,用Preason’s法進行相關性分析,采用t檢驗評估相關系數。當P<0.05時,表示差異顯著。
2 結果與分析
2.1 飼糧添加CMH和CLA對大鼠生長性能的影響
由圖1可知,飼糧添加CMH和CLA后,CMH低、高劑量組大鼠的生長速率高于對照組,而CLA低、高劑量組則低于對照組,但各組間無明顯差異。由圖2可知,飼糧添加CMH和CLA后,大鼠的日攝入量較對照組均顯著降低(P<0.05),但CMH低、高劑量組和CLA低、高劑量組間無顯著差異(P>0.05)。已有研究發現,飼糧添加CLA對豬平均日增體質量無顯著影響,但總體來說,日攝入量隨著CLA添加量的增加呈線性下降[11],這與本實驗研究結果一致,可能是因為隨著CLA添加量的增加,飼糧能量也在增加,當飼糧能量大于大鼠需要量時,會影響采食量,進而使得日攝入量降低。
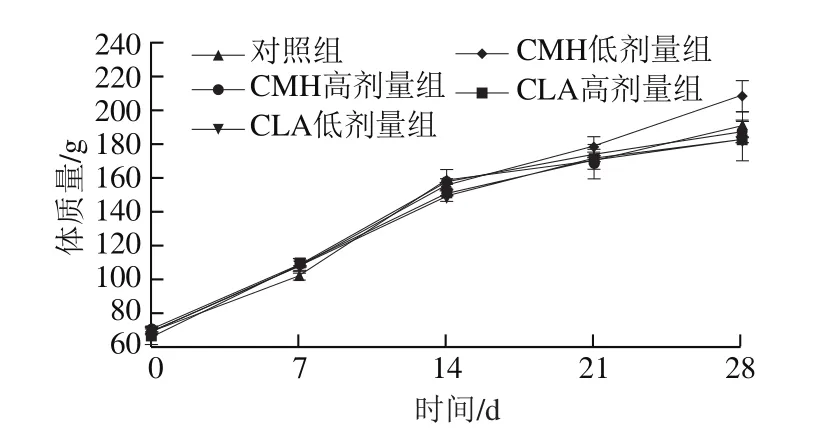
圖1 飼糧添加CMH和CLA對大鼠體質量的影響Fig.1 Effect of CMH and CLA on body mass of rats
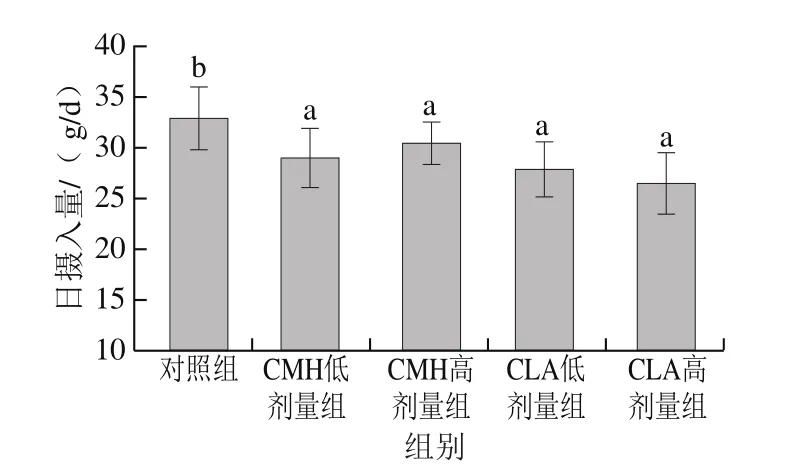
圖2 飼糧添加CMH和CLA對大鼠日攝入量的影響Fig.2 Effects of CMH and CLA on daily feed intake in rats
2.2 飼糧添加CMH和CLA對大鼠肌纖維類型組成的影響
Chang等[12]根據肌纖維特有的MyHC類型將豬肌纖維分為4 種,并研究了豬骨骼肌MyHC I、MyHC IIa、MyHC IIb和MyHC IIxmRNA的表達。建立在MyHC表達基礎上的分子分型較組織化學評定更為準確、可靠[13]。因此,本實驗在大鼠基礎飼料中添加不同劑量CMH和CLA,研究大鼠腓腸肌中肌纖維MyHCmRNA相對表達量變化。
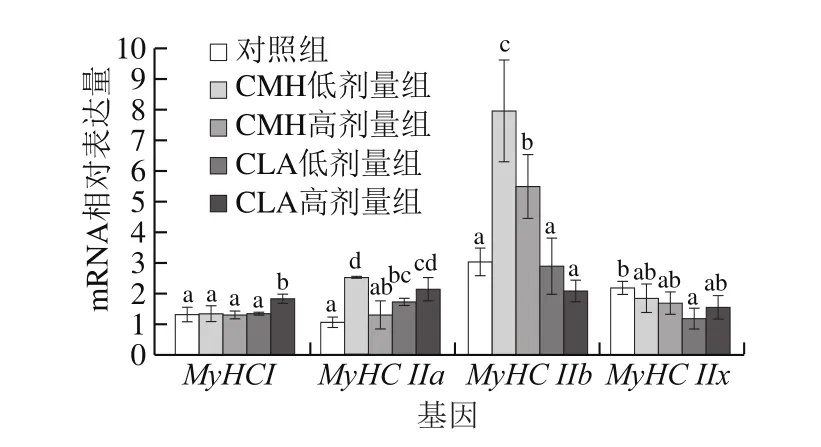
圖3 飼糧添加CMH和CLA對大鼠腓腸肌MyHC mRNA相對表達量的影響Fig.3 Effects of CMH and CLA on MyHC mRNA expression in rat gastrocnemius
由圖3可知,與對照組相比,CMH低劑量組MyHC IIa及CMH低、高劑量組MyHC IIbmRNA相對表達量顯著增加(P<0.05);同時各實驗組MyHC IIxmRNA相對表達量降低,且CMH低、高劑量組間差異不顯著(P>0.05)。因此,飼糧添加CMH增加了大鼠腓腸肌MyHC IIa、MyHC IIbmRNA相對表達量,且低劑量的CMH對肌纖維類型組成的影響較高劑量更為明顯。門小明等[14]研究表明,日糧中添加0.5% CMH能顯著降低豬背最長肌MyHC IIamRNA相對表達量,與本實驗結果相反,造成此差異的原因可能是因為動物的品種、添加量、肌肉部位等因素的不同。
飼糧添加CLA有增加MyHC ImRNA相對表達量的趨勢,其中CLA高劑量組顯著高于對照組(P<0.05)。CLA低、高劑量組MyHC IIamRNA相對表達量均顯著高于對照組(P<0.05)。與對照組相比,CLA低、高劑量組MyHC IIb、MyHC IIxmRNA相對表達量均有所降低,但僅CLA低劑量組MyHC IIxmRNA相對表達量發生顯著變化(P<0.05),CLA低、高劑量組間MyHC IIa、MyHC IIbmRNA相對表達量差異不顯著(P>0.05)。因此,飼糧添加CLA可以增加大鼠腓腸肌氧化型肌纖維的表達,降低酵解型肌纖維的表達,提示飼糧添加CLA可以促進肌纖維由酵解型向氧化型轉變,并且與添加CMH相反,高劑量的CLA對于肌纖維類型組成的影響較低劑量更明顯。有研究指出,在生長育肥豬日糧中添加1.5%的CLA,可顯著提高MyHC I和MyHC IIa的mRNA相對表達量,同時顯著降低MyHC IIb和MyHC IIx的mRNA相對表達量[15]。Huang等[16]也發現,飼糧添加CLA能顯著提高育肥豬肌肉中MyHC I和MyHC IIa的mRNA相對表達量,降低MyHC IIxmRNA相對表達量,但對MyHC IIbmRNA相對表達量影響不顯著。以上研究結果與本實驗研究結果一致,即飼糧添加CLA對MyHC I和MyHC IIa型肌纖維有積極影響。
2.3 飼糧添加CMH和CLA對大鼠腓腸肌LDH、SDH、MDH活力的影響
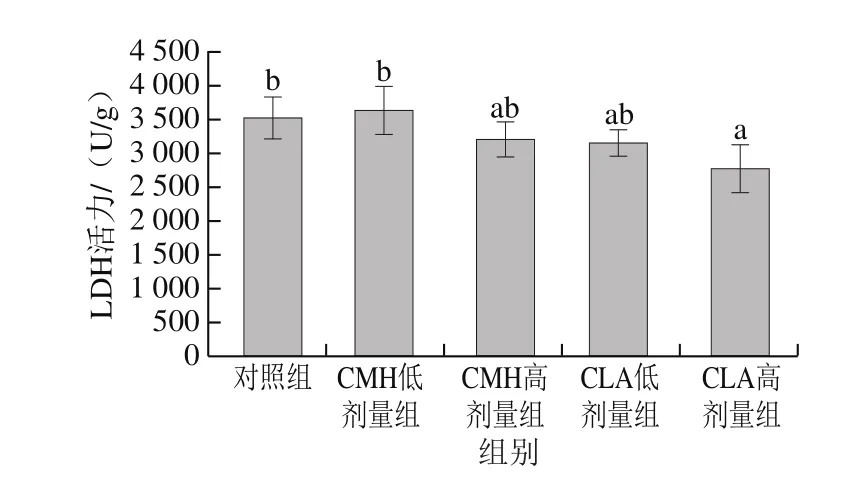
圖4 飼糧添加CMH和CLA對大鼠腓腸肌LDH活力的影響Fig.4 Effects of CMH and CLA on LDH activity in rat gastrocnemius
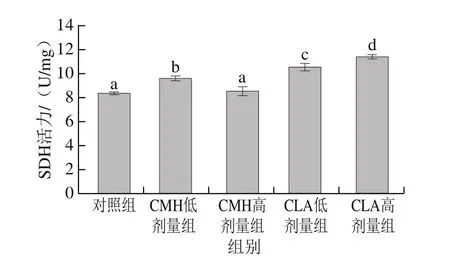
圖5 飼糧添加CMH和CLA對大鼠腓腸肌SDH活力的影響Fig.5 Effects of CMH and CLA on SDH activity in rat gastrocnemius
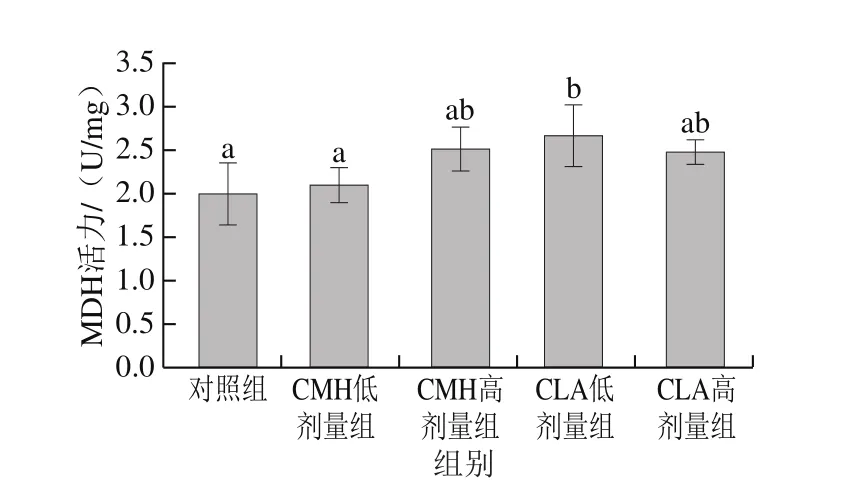
圖6 飼糧添加CMH和CLA對大鼠腓腸肌MDH活力的影響Fig.6 Effects of CMH and CLA on MDH activity in rat gastrocnemius
不同肌纖維類型組成的肌肉,其能量代謝類型也存在差異,氧化型肌纖維中線粒體含量豐富,有較高活性的有氧代謝酶,主要通過有氧代謝途徑供能;酵解型肌纖維中線粒體含量較少,但含有較高活性的酵解型酶,主要通過糖酵解方式供能[17-18]。LDH在糖酵解過程中起著重要作用,是糖酵解產生乳酸的催化酶,其活性高低能夠反映細胞內無氧酵解的活躍程度[19]。由圖4可知,與對照組相比,飼糧添加高劑量CMH、CLA可以降低大鼠腓腸肌中LDH活力,其中CLA高劑量組與對照組差異顯著(P<0.05)。說明飼糧添加CMH和CLA后,大鼠腓腸肌酵解酶活力減弱。SDH和MDH是葡萄糖有氧氧化過程中的限速酶,可以反映線粒體有氧代謝程度,評價肌肉氧化活性[20]。由圖5可知,CMH低劑量組和CLA低、高劑量組SDH活力均顯著高于對照組(P<0.05),由圖6可知,CLA低劑量組MDH活力顯著高于對照組(P<0.05),總體來看,飼糧添加CMH和CLA后,大鼠腓腸肌SDH和MDH活力均有不同程度的提高,這說明大鼠腓腸肌氧化酶活力升高。這與添加CMH和CLA后氧化型肌纖維比例升高、酵解型肌纖維比例下降這一結果一致。
2.4 飼糧添加CMH和CLA對大鼠腓腸肌AMPK活力的影響
有研究表明,飼糧添加CMH可以激活AMPK信號通路,并降低糖酵解酶活性[21],添加不飽和脂肪酸能夠提高豬肌肉中AMPK的表達,增加氧化型肌纖維的比例[8]。AMPK在肌肉能量代謝方面發揮重要作用,與肌纖維類型轉化密切相關。AMPK基因缺陷型小鼠肌纖維類型在由酵解型向氧化型轉變時會受到阻礙[22],這表明AMPK對氧化型肌纖維有正反饋作用。侯艷茹等[23]研究表明不同的飼喂環境會導致蘇尼特羊背最長肌AMPK活性不同。也有研究表明,不同肌肉中(如比目魚肌、腓腸肌、四頭肌)因I、IIa、IIb型和IId/x型肌纖維比例差別較大,AMPK活性也有較大差異[24]。因此,在大鼠基礎飼料中添加不同劑量的CMH和CLA后,測定大鼠腓腸肌AMPK活力變化。
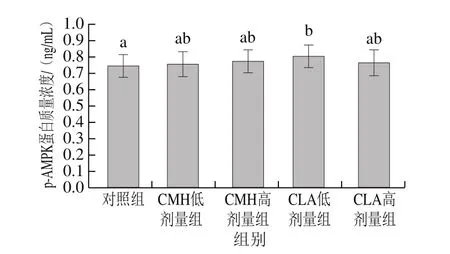
圖7 飼糧添加CMH和CLA對大鼠腓腸肌AMPK活力的影響Fig.7 Effects of CMH and CLA on AMPK activity in rat gastrocnemius
由圖7可知,飼糧添加CMH和CLA后,CMH和CLA低、高劑量組大鼠腓腸肌AMPK活力均高于對照組,且CLA低劑量組顯著高于對照組(P<0.05)。有報道稱,日糧添加多不飽和脂肪酸可以促進AMPK基因的表達[25],對小鼠飼喂不飽和脂肪酸后發現,AMPK基因表達量顯著增加[9],李蛟龍等[21]認為飼糧添加CMH對AMPKmRNA表達有顯著促進作用,這與本實驗研究結果相似。因此,飼糧添加CMH和CLA很可能激活了AMPK信號通路,進而導致大鼠腓腸肌肌纖維類型的改變。
過氧化物酶增殖體激活受體γ輔激活因子1α(peroxisome proliferator-activated receptor γ coactivator 1α,PGC-1α)已被證明是線粒體生物發生的主要調節器,很多研究表明PGC-1α基因表達可通過運動以及AMPK的激活來誘導[22]。為進一步研究飼糧添加CMH和CLA對大鼠腓腸肌肌纖維類型組成的影響機制,本實驗對肌纖維相關基因表達量進行測定。
2.5 添加CMH和CLA對大鼠PGC-1α、MEF2C和GLUT4 mRNA相對表達量的影響
PGC-1α是一種轉錄共激活因子[26],在線粒體生物發生中發揮重要作用[27],其表達量與氧化型肌纖維比例呈正相關,小鼠體內過表達PGC-1α后,小鼠肌肉中線粒體數量和功能均增強,同時I型肌纖維比例增加[28]。在小鼠成肌細胞中,AMPK可以直接或間接調控PGC-1αmRNA的表達[29]。肌細胞增強因子2C(myocyte enhancer factor 2C,MEF2C)是PGC-1α的一個主要輔助活化因子,可直接調控PGC-1αmRNA的表達,提高氧化代謝[30]。任陽[9]的研究結果表明,飼喂不飽和脂肪酸的小鼠比目魚肌中MEF2C基因表達顯著增加。葡萄糖轉運子4(glucose transporter 4,GLUT4)是MEF2C的上游因子,MEF2C可以和PGC-1α結合誘導GLUT4的表達,促進葡萄糖轉運[31]。

圖8 飼糧添加CMH和CLA對大鼠腓腸肌肌纖維類型轉化相關基因表達量的影響Fig.8 Effects of CMH and CLA on muscle fiber type transition-related gene expression in rat gastrocnemius
由圖8可知,CMH低劑量組和CLA低、高劑量組大鼠腓腸肌PGC-1αmRNA相對表達量顯著高于對照組(P<0.05);對于MEF2CmRNA相對表達量,與對照組相比,CMH低、高劑量組顯著增加(P<0.05),CLA高劑量組顯著下降(P<0.05);CMH低、高劑量組和CLA高劑量組與對照組相比,大鼠腓腸肌GLUT4mRNA相對表達量顯著增加(P<0.05)。總的來說,除高劑量的CLA顯著降低MEF2CmRNA相對表達量外,飼糧添加CMH和CLA均能增加PGC-1α、MEF2C和GLUT4mRNA相對表達量,與上述AMPK活力增加這一結果相一致,同時CMH對于肌纖維類型轉化相關基因的影響較CLA更顯著。有研究表明,當激活AMPK時,GLUT4和PGC-1α的表達顯著提高,并且氧化酶活性增強,糖酵解酶活性降低[32-33],這與本實驗結果一致。綜上所述,飼糧添加CMH和CLA后大鼠腓腸肌肌纖維類型的轉化很可能是由于AMPK被激活,進而促進PGC-1α的表達,PGC-1α結合并輔助激活了MEF2C,提高氧化代謝,促進GLUT4基因的表達。
2.6 MyHC表達水平與AMPK活力、肌纖維轉化相關基因表達水平及酶活力的相關性分析結果
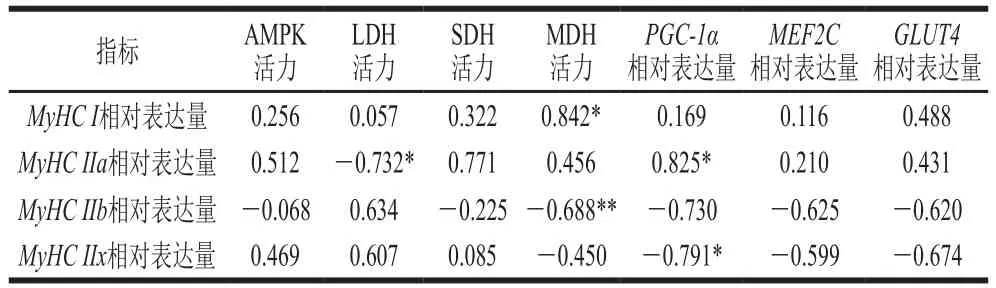
表2 MyHC表達水平與AMPK活力、肌纖維轉化相關基因表達水平及酶活力相關性Table 2 Correlation analysis of MyHC expression with AMPK activity,muscle fiber transformation-related gene expression and enzyme activities
由表2可知,AMPK活力與MyHC I、MyHC IIa和MyHC IIx相對表達量之間呈正相關,與MyHC IIb相對表達量呈負相關;LDH活力與MyHC I、MyHC IIb和MyHC IIx相對表達量之間呈正相關,與MyHC IIa相對表達量呈顯著負相關(P<0.05);SDH和MDH活力與MyHC I和MyHC IIa相對表達量之間呈正相關,與MyHC IIb相對表達量呈負相關。PGC-1α、MEF2C和GLUT均對MyHC I和MyHC IIa型肌纖維存在正向調控作用,對MyHC IIb和MyHC IIx型肌纖維存在負向調控作用。
3 結 論
通過對飼糧中添加了CMH和CLA的大鼠生長性能進行測定及分析,結果表明,飼糧中添加CMH和CLA對大鼠生長速率無顯著影響,但能顯著降低大鼠日攝入量,這說明CMH和CLA對提高飼料轉化率有積極作用。本研究結果顯示,飼糧添加CMH,上調了大鼠腓腸肌中MyHC I、IIa、IIbmRNA相對表達量,同時肌肉中氧化酶(SDH、MDH)活力及AMPK活力和PGC-1α、MEF2C、GLUT4mRNA相對表達量升高,但降低了MyHC IIxmRNA相對表達量及酵解酶(LDH)活力。飼糧添加CLA提高了大鼠腓腸肌中氧化型肌纖維比例,同時提高了MDH、SDH、AMPK活力和PGC-1α、GLUT4mRNA基因表達量,但降低了酵解型肌纖維比例和LDH活力。這說明飼糧添加CMH和CLA能夠增加AMPK活力,上調AMPK通路相關基因表達,增強肌肉氧化代謝能力,使肌纖維類型發生轉變。因此,未來畜牧業養殖過程可通過飼喂CMH和CLA,激活AMPK信號通路來調控肌纖維類型的轉化,從而提高畜肉品質。
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
今日農業(2022年4期)2022-11-16 19:42:02
鄉村科技(2021年33期)2021-03-16 02:26:54
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
藥學與臨床研究(2015年4期)2015-06-05 11:35:54
衛生職業教育(2014年24期)2014-05-20 09:05:38
同位素(2014年2期)2014-04-16 04:57:20
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
