Prpf40b 基因缺失誘發大鼠心臟結構異常
2021-05-18 05:55:58劉夢迪董偉張旭陳煒高翔高珊張連峰呂
中國比較醫學雜志 2021年4期
關鍵詞:實驗
劉夢迪董 偉張 旭陳 煒高 翔高 珊張連峰呂 丹*
(1.中國醫學科學院醫學實驗動物研究所,北京協和醫學院比較醫學中心,北京市人類重大疾病實驗動物模型工程技術研究中心,北京 100021; 2.中國醫學科學院醫學實驗動物研究所,北京協和醫學院比較醫學中心,國家衛生健康委員會人類疾病比較醫學重點實驗室,北京 100021)
剪接體在真核細胞新生RNA 轉錄物的加工中起重要作用。 剪接體是小核RNA 和蛋白質的復合物,可從初級轉錄本(pre-mRNA)中去除(剪接)內含子和非常規外顯子,從而將識別的外顯子組裝(剪接)成成熟的mRNA。 它主要是基因特異性的,并且是由于順式調控序列中的突變指導了外顯子的剪接識別[1]。 但是,已知有越來越多的突變靶向剪接體的組分,因此潛在地產生了全局而非基因特異性的RNA 剪接變化。 剪接位點的識別可以通過跨越內含子以及跨越外顯子兩種機制進行,該方式可極大的增加蛋白質的多樣性和基因表達的復雜程度[2-3]。
近十年來,已有多篇研究報道剪接復合體相關基因突變與多種臨床疾病相關[4-5]。 而PRPF40B是已發現的編碼剪接體化合物的基因之一,該類基因還包括, SF3B1, SRSF2, U2AF1, ZRSR2, SF1,SF3A1 及U2AF2。 前mRNA 加工因子(Pre-mRNA processing factor,PRPF)是研究發現的一種參與前mRNA 剪接的蛋白質。 人PRPF 基因編碼一個1007個氨基酸的蛋白質,包含一個N-末端340 個氨基酸的富含Arg/Ser 的結構域,該結構域通常存在于前mRNA 剪接因子中。 PRPF 的催化結構域與細胞周期蛋白依賴性激酶和絲裂原活化蛋白激酶具有同源性[6]。
PRPF40B 屬于U2 核糖核蛋白體依賴型剪接復合體相關基因,目前,已有研究表明PRPF40B 基因突變, 與骨髓增生異常綜合征(myelodysplastic syndromes,MDS),慢性淋巴細胞白血病(chronic lymphocytic leukemia,CLL)和急性粒細胞白血病(acute myeloid leukemia,AML)有關[7-10]。 在患有骨髓惡性腫瘤的患者中,已經描述了八個核心剪接體基因中的體細胞突變-SF3B1, SRSF2, U2AF35( U2AF1 ), ZRSR2, SF3A1, PRPF40B, U2AF65(U2AF2),其中PRPF40B 的突變發生頻率為1%左右[1,11-12]。 此外,有研究表明PRPF40B 基因突變還與多種實體腫瘤發生相關,包括乳腺癌,肝癌,胰腺癌和黑色素瘤等[2,13]。
所以, 為深入研究剪接復合體相關基因PRPF40B 的生物學作用,動物模型是重要的工具,本實驗室依托研究所和基因工程技術平臺,利用CRISPR/Cas9 技術首次構建了PRPF40B 基因敲除大鼠模型,為開展該基因的生物學功能研究,以及其在多種疾病病理進程中所扮演角色的深入研究,提供了體內動物模型工具。 并且在初步的研究分析中,發現該基因敲除可誘發心臟結構形態異常改變。
1 材料和方法
1.1 實驗動物
12 月齡雄性SPF 級Prpf40b 基因敲除SD 大鼠及同窩對照大鼠各6 只,體重550 ~720 g,由中國醫學科學院醫學實驗動物研究所實驗動物資源研究中心建立及培育,同時敲除大鼠培育過程中使用的SD 大鼠,均購自北京維通利華實驗動物技術有限公司[SCXK(京)2019-0011]。 大鼠飼養于中國醫學科學院醫學實驗動物研究所屏障環境動物房[SYXK(京)2019-0014],同時動物飼養間采用12 h交替明暗照明,動物自由飲水攝食。 同時,動物實驗中涉及的動物操作程序已得到中國醫學科學院醫學實驗動物研究所實驗動物使用及管理委員會(IACUC)的批準(ZLF18003),實驗過程中遵循了3R 原則。
1.2 主要試劑與儀器
PCR 引物由北京天一輝遠生物科技有限公司合成;4%甲醛溶液購自北京益利精細化學品有限公司;DNA 提取試劑盒購自北京全式金生物技術有限公司;瓊脂糖購自西班牙Biowest Agarose 公司;凝膠電泳儀購自美國伯樂公司;組織脫水機、石蠟包埋機、石蠟切片機購自德國徠卡公司;PCR 儀購自美國伯樂公司;氣體麻醉機購自英國斯特普科技有限公司;病理切片掃描機購自德國徠卡公司;小動物成像系統Vevo3100 購自加拿大VisualSonics 公司。
1.3 實驗方法
1.3.1 載體構建及基因型鑒定(vector construction and genotyping)
利用本實驗室已經建立的成熟的CRISPR/Cas9技術操作平臺[14],構建Prpf40b 基因敲除大鼠。 首先研究人員依據Prpf40b 基因第二外顯子,設計sgRNA,并利用MEGAshortscript T7 轉錄試劑盒(AM1354,Ambion,美國)。 將Cas9 蛋白(30 ng/μL)和sgRNA(皆10 ng/μL)混合后,經顯微注射到受精卵、移植至假孕母子子宮內,手術完成后常規飼養。
對出生后仔鼠,制備組織基因組,經PCR 鑒定其基 因 型, PCR 引 物: 5’ TCCTCGACAGCCACA TACTCAG3’ 及5’ GTCTTCACTTCGACCTGGTTCA G3’。 PCR 條件:94℃預變性4 min,94℃變性20 s,60℃退火1 min,72℃延伸1.5 min,35 個循環。 野生條帶和敲除條帶分別是1890 bp 和809 bp。
1.3.2 超聲影像分析技術(echocardiography analysis)
大鼠經異氟烷(1.5%~2.5%)麻醉,胸前區脫毛,仰臥位固定于加熱板上,保持體溫恒定,同時四肢固定,待呼吸平穩后,利用16 兆赫茲探頭(Vevo3100, 加拿大),經胸前區長軸和短軸兩個切面,采集B-mode 和M-mode 信號,經3100 信號及數據分析系統,導出超聲參數,包括左室收縮末期內徑 ( LVESD, left ventricular ( LV ) end-systole diameter); 左室 舒張 末 期內 徑( LVEDD, left ventricular(LV)end-diastole diameter);左室收縮末期容積(LVESV, left ventricular (LV) end-systole volume); 左室舒張末期容積( LVEDV, left ventricular(LV)end-diastole volume);每搏輸出量(SV, stroke volume)等。
1.3.3 病理組織學分析(histological analysis)
大鼠經安樂死后,摘取心臟,經生理鹽水清洗后,固定于4%甲醛中固定48 h。 隨后經脫水后的心臟組織經智能石蠟包埋機包埋制備成蠟塊,推拉式切片機進行切片(切片厚度4 μm),恒溫烤箱將切片烤干后,經全自動染色機完成HE(hematoxylineosin stain)染色,自動封片機完成封片。 切片掃描(徠卡TCS SP2, 德國) 后經Aperio ImageScope v8.2.5 軟件分析并導出病理圖片。
1.4 統計學方法
數據采用GraphPad Prism 7 統計軟件處理,數據以平均數±標準誤差(±s)來表示,均數間采用Student’st-tests 分析,P<0.05 為有顯著性差異。
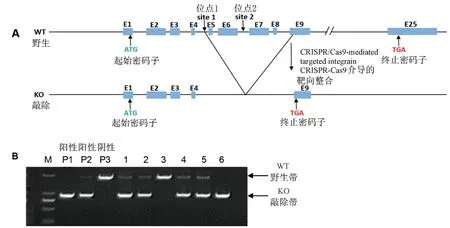
注:A:利用CRISPR/Cas9 技術,Prpf40b 基因敲除大鼠構建示意圖,site1、site2:靶點設計位置。 B:Prpf40b 基因敲除大鼠基因型PCR 鑒定電泳凝膠圖像,P1~P3 分別是敲除純合子,雜合子及同窩野生型對照孔;1、2、4、5:敲除雜合子大鼠基因型條帶;3:同窩野生型基因型條帶;6:敲除純合子大鼠基因型條帶。圖1 Prpf40b 基因敲除大鼠構建及鑒定Note. A, Schematic diagram of Prpf40b knockout rat with CRISPR/Cas9 technology. site1/site2,target design sites.B,Genotyping identification of Prpf40b knockout rat through PCR. P1 ~P3, Sample for reference of homozygosis,heterozygosis and wild-type. 1,2,4 and 5, Heterozygosis band. 3, Wild-type band. 6, Homozygosis band.Figure 1 Establishment and genotyping of Prpf40b knockout rat
2 結果
2.1 Prpf40b 基因敲除大鼠的建立及繁育
利用CRISPR/Cas9 技術,建立Prpf40b 基因敲除大鼠,經測序及PCR 技術鑒定,敲除大鼠第5、6、7、8 以及部分第9 外顯子已敲除,構建示意圖見圖1A。
將首建鼠與野生型SD 大鼠進行雜交,PCR 技術鑒定仔代鼠基因型,仔代敲除雜合子大鼠間再進行第二輪雜交,仔代大鼠即可獲得敲除純合子,PCR鑒定示意圖見圖1B。
觀察至12 月齡時,Prpf40b 基因敲除雜合子和純合子大鼠外觀形態及體重等,未見明顯異常。 隨后的心臟超聲影像學及病理組織學實驗,皆選取Prpf40b 基因敲除純合子大鼠及同窩陰性大鼠進行觀察及分析。
2.2 Prpf40b 基因敲除大鼠心臟結構及功能分析
經基因型鑒定后,選用2 月齡Prpf40b 基因敲除大鼠和同窩陰性對照大鼠,進行心臟超聲影像分析,結果顯示,與同窩陰性大鼠相比,敲除大鼠心臟結構及形態各參數未見明顯異常(數據未列出)。
隨后選用12 月齡敲除大鼠和同窩陰性對照大鼠,進行心臟超聲影像分析,同時采集B-mode 和Mmode 影像。 通過M-mode 影像觀察,12 月齡敲除大鼠,左室內徑明顯減小(圖2)。
隨后通過Veov3100 分析系統分析并導出各超聲參數,結果顯示,左室舒張末期內徑(LVEDD, left ventricular(LV)end-diastole diameter)顯著變小,減少了15.6%(圖3A,P=0.0069);左室收縮末期內徑 ( LVESD, left ventricular ( LV ) end-systole diameter)顯著變小,減少了22.2%(圖3B,P=0.0134); 左室舒張末期容積( LVEDV, left ventricular(LV)end-diastole volume)顯著降低,降低了31.5%(圖3C,P=0.0068);左室收縮末期容積(LVESV, left ventricular(LV)end-systole volume)顯著降低,降低了44.1%(圖3D,P=0.0057);同時,每搏輸出量(SV, stroke volume)和心輸出量(CO,cardiac output)顯著降低,分別降低了26.1%(圖3E,P= 0.0229)和25.9%(圖3F,P= 0.0306)。
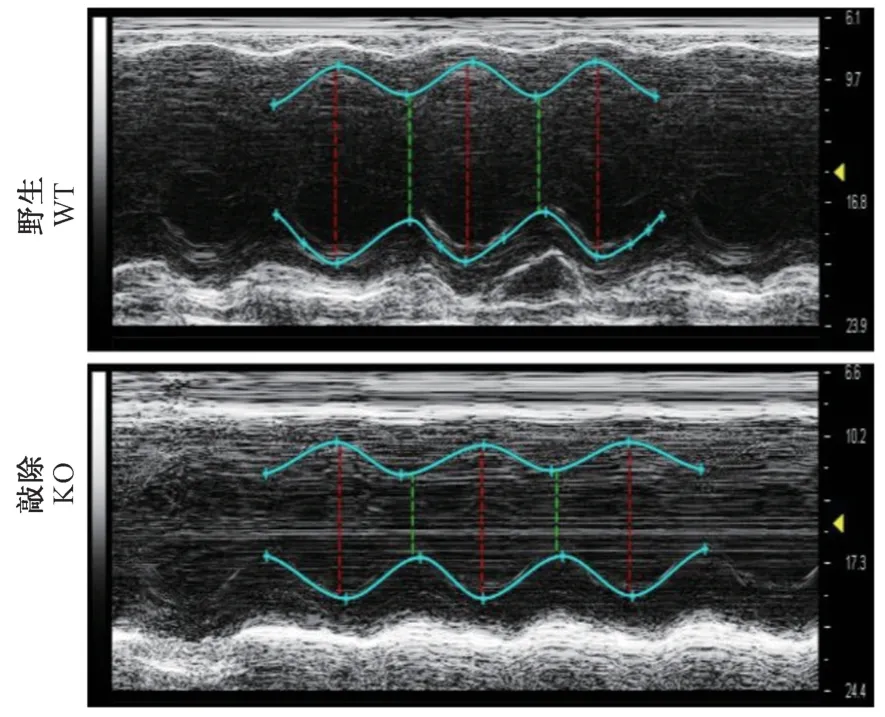
圖2 Prpf40b 基因敲除大鼠超聲影像分析M-mode 截圖Figure 2 M-mode screenshot from echographic analysis in Prpf40b knockout rat
2.3 Prpf40b 基因敲除大鼠心肌組織學觀察
通過超聲影像分析,12 月齡Prpf40b 基因敲除大鼠心臟結構形態出現異常,隨后我們選取12 月齡敲除大鼠,經摘取心臟,常規固定并制備心肌組織石蠟切片,隨后進行HE 染色,切片掃描后經Aperio分析心肌組織學改變。
與同窩陰性對照大鼠相比,敲除大鼠心臟整體變小,心室腔變小(圖4A),這與敲除大鼠通過超聲影像學觀察到的結果一致。 在顯微水平,與同窩陰性對照大鼠相比,敲除大鼠心肌纖維出現排列不齊,心肌纖維粗細不均一,同時,心肌纖維間肌漿網明顯擴張,以及心肌纖維染色著色不均一等現象(圖4B)。
同時,我們通過統計12 月齡敲除大鼠心臟濕重與體重比值,即心重體重比(HW/BW, heart weight/body weight),進一步確認心臟形態學改變。 結果我們發現,敲除大鼠心重體重比顯著下降,下降了15.6%(圖4C,P=0.0348),與超聲影像學及病理組織學觀察結果一致。
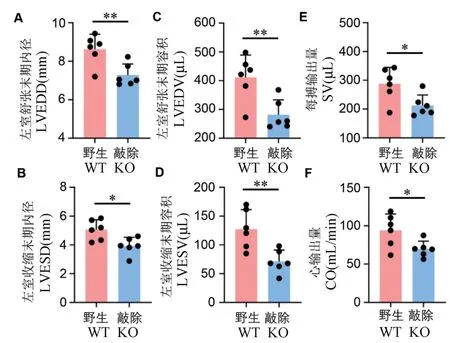
注:A:LVEDD,左室舒張末期內徑;B:LVESD,左室收縮末期內徑;C:LVEDV,左室舒張末期容積;D:LVESV,左室收縮末期容積;E:SV,每搏輸出量;F:CO,心輸出量。 與野生對照組比較,*P<0.05,**P<0.01。圖3 Prpf40b 基因敲除大鼠超聲影像參數分析Note. A, LVEDD, left ventricular(LV)end-diastole diameter. B,LVESD,left ventricular(LV)end-systole diameter. C, LVEDV, left ventricular(LV)end-diastole volume. D, LVESV, left ventricular(LV)end-systole volume. E, SV, stroke volume. F, CO, Cardiac output. Compared with wild type rats, *P<0.05, **P<0.01.Figure 3 Echography parameter analysis in Prpf40b knockout rat
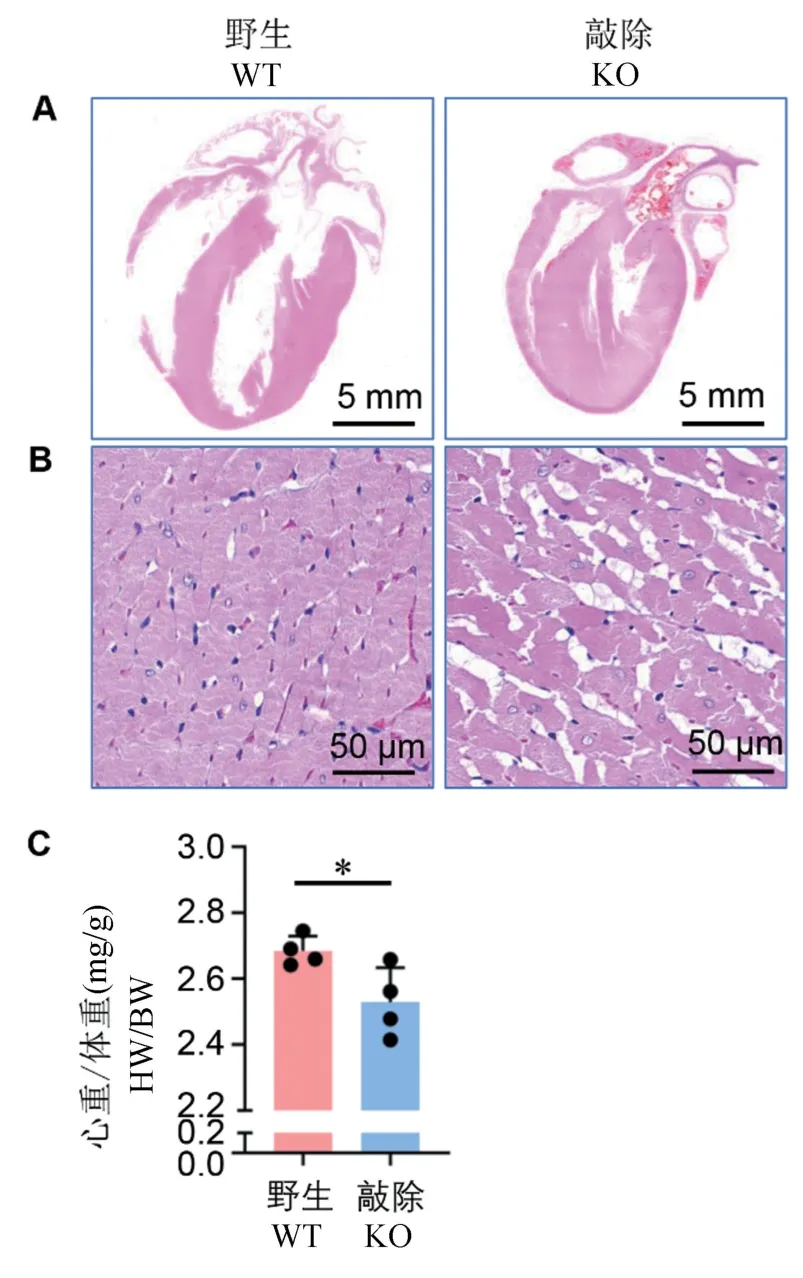
注:A:12 月齡Prpf40b 敲除大鼠心臟長軸整體切片HE 染色;B:敲除大鼠心臟長軸整體切片HE 染色高倍截圖;C:敲除大鼠心重體重比。 與野生對照組比較,*P<0.05。圖4 Prpf40b 基因敲除大鼠病理組織學分析Note. A, The whole-heart longitudinal sections with HE staining from Prpf40b knockout rat at 12 months old. B, Magnification of the whole-heart longitudinal sections with HE staining from Prpf40b knockout rat. C, Ratio of heart weight/ body weight.Compared with wild type rats, *P<0.05.Figure 4 Histological analysis in myocardial of Prpf40b knockout rat
3 討論
酵母剪接因子Prp40 是一種必需的U1-snRNP相關蛋白,作為支架參與剪接體復合物形成的早期步驟。 Prp40 的兩種哺乳動物同源基因包括PRPF40A,也被稱為HYPA(亨廷頓相互作用蛋白A) 或 FBP11 ( formin-binding protein 11 ) 和PRPF40B,即已知的HYPC(亨廷頓相互作用蛋白C)[15]。
研究表明PRPF40A 和PRPF40B 參與剪接體的早期組裝,但功能尚不清楚。 PRPF40B 與神經系統疾病的發病機制有關,包括亨廷頓病和Rett 綜合征。 此外, 近幾年骨髓增生異常綜合征(myelodysplastic syndromes,MDS)中檢測到參與剪接位點識別的剪接體成分的突變[16]。 PRPF40B 是剪接體突變檢測到的靶點之一,在整個開放閱讀框架中,PRPF40B 的變化以錯義突變的形式出現,提示這些突變可能導致功能喪失。 PRPF40B 定位于富含剪接因子的核斑點,結合SF1 和U2AF65,并在體內調節多種剪接事件。 研究顯示PRPF40B 缺失增加了Fas/CD95 受體數量和細胞凋亡,表明PRPF40B 能夠改變關鍵凋亡基因的選擇性剪接來調節細胞存活。 這些結果支持了PRPF40B 在導致外顯子選擇性剪接的早期事件中的作用,并可能為PRPF40B 相關疾病的分子機制提供重要參考[17]。
以往的研究表明,PRPF 的突變導致細胞周期中前mRNA 的積累和G1/S 轉換受損。 PRPF 對激酶、轉錄因子、染色質重塑因子、紡錘體檢查點蛋白和癌細胞生長具有不同的作用。 PRPF 被證明是U5 snRNP 相關激酶,PRPF 與BRG1 和N-CoR 相互作用,這是核激素共激活和共抑制復合物的兩個組成部分。 PRPF 與U5-snRNP 和N-CoR 脫乙酰酶復合物相關,被認為可以協調轉錄調控中的前mRNA剪接和染色質重塑事件[18-20]。 PRPF 可觸發上皮間質轉化(epithelial-mesenchymal transition, EMT),降低HCT116、PC3、B16-F10 黑色素瘤細胞的侵襲性。PRPF 可通過激活細胞生存信號通路、重新排列肌動蛋白骨架和誘導EMT 來阻斷白藜蘆醇的凋亡作用。
PRPF40B 可抑制髓系細胞缺氧,其低表達可能導致白血病的發生[21]。 目前已有報道PRP6、SWAP 和PNN 與PRPF 相互作用,并且PRPF 被證明是U5 snRNP 相關激酶。 特別是,小核糖核蛋白復合物的成員PRPF6 被證明能促進結腸癌的增殖。最近的研究表明,各種選擇性剪接事件模式的改變在結腸癌的發生和發展中起著重要的作用[6]。
Prp40 蛋白結構與人類中相對少量的蛋白(包括PRPF40A,PRPF40B 和hTCERG1)共有,它們均參與剪接和轉錄延伸的偶聯。 最接近的人類直系同源物PRPF40A 的功能與酵母Prp40 類似。PRPF40A 的WW 域對于其定位到核斑點至關重要。 PRPF40A 與SF1 和U2AF 相互作用,它存在于剪接體復合體A 和B 中,但不存在于C 中,這與Prp40 在早期剪接體組裝中的作用一致。 PRPF40A與U2-snRNP 和參與轉錄延伸的蛋白質可發生免疫共沉淀。
PRPF40A 和PRPF40B 都通過Pol II CTD 與延伸復合物相互作用調節共轉錄剪接,并且都與亨廷頓氏病大腦中擴展的亨廷頓多谷氨酰胺束結合,并參與其他神經退行性綜合征。 即使PRPF40B 不具有任何RNA 識別基序(RRM),它也可能通過橋接5′s 的U1 和與BPS 結合的SF1 來調節AS。 此外,PRPF40B 控制重要凋亡基因(例如Fas)的其他剪接位點的選擇。 PRPF40B KO 上調了低氧信號,再加上其在AML 中的低表達,提示該SF 在髓樣白血病中的作用。 在 Akt/MAPK 通路中發現了PRPF40B 調控的酶的PPI 聚集,這是已知的相互調節缺氧的途徑[21]。
在本研究中我們發現PRPF40B 基因敲除可誘發模型大鼠心臟結構形態的改變,包括整體結構形態及顯微形態的改變,其機制可能與該基因控制重要凋亡基因相關,但具體的誘導機制,還需要后續深入的分子機制研究。 同時本文建立的PRPF40B基因敲除大鼠為該基因參與多種疾病病理進程調控的作用研究,提供了體內工具動物,為剪接復合體相關基因的功能研究提供了實驗基礎。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
