美麗異木棉復合體的花色變化規律研究
2021-05-17 05:56:08吳欣黃頌誼黃桂蓮
廣東園林 2021年2期
吳欣 黃頌誼 黃桂蓮
吉貝屬Ceiba隸屬于錦葵科(APGIV 系統),原分布于中南美洲和非洲西部,包括17 個物種[1]。該屬大部分物種因觀賞價值較高而被廣泛引種栽培于世界各地[2~3]。據記載,我國于20 世紀60 年代開始引種“美麗異木棉”于海南尖峰嶺,現今在兩廣地區、福建、海南和香港等地廣泛栽培[2~3]。國內引進的“美麗異木棉”作為園林常用樹種,不僅花期長、花色美、觀賞價值高,也具有易繁殖、生長快和抗性強的特點[4~6]。根據近期的文獻報道,基于形態和分子數據分析表明,目前我國華南地區栽培的“美麗異木棉”包括了吉貝屬內至少3 個物種以及它們的雜種后代,其中涉及的物種有美麗異木棉C. speciosa、C. insignis和C.crispiflora[7~8]。由于國內園林常用的“美麗異木棉”并非為一個物種,因此本研究將吉貝屬多個物種及其雜種稱為“美麗異木棉復合體”。
美麗異木棉復合體的花瓣顏色組成較為復雜,花瓣先端顏色分為白色(淺灰色)、淺紅色、紅色、深紅色和紫紅色,花瓣基部顏色分為白色、黃白色、淺黃色和黃色[9]。目前,美麗異木棉復合體在園林應用上個體顏色參差不齊,難以形成具有群體效應的景觀,對花色變化研究有利于選育出觀賞價值高、花色統一的美麗異木棉復合體優良種質,形成具有群體效應的景觀。有文獻將美麗異木棉復合體的花瓣先端顏色和基部顏色進行分類統計[7~9],但并沒有進行顏色變化相關的研究。而國內有關美麗異木棉復合體的文獻報道,主要涉及形態特征[9]、光合特性[10~11]、栽培繁殖[12~18]、病理學[19]、種質分析[7~8,20]等方面。本研究對美麗異木棉復合體的花色變化規律進行觀察分析,以期為美麗異木棉復合體的品種選育提供參考。
1 材料與方法
1.1 試驗地點及對象
試驗地點在廣州市白云區的陳田花園,觀測對象為來自8 株不同花色的美麗異木棉復合體個體的108 朵花,編號1~108,花色類型涵蓋了粉白、粉紅和紫紅色3 種。為減少環境因素如溫差、降雨、氣流等變化的影響,觀測對象均為在同一天(2019 年12月11 日)開放的花朵。觀測時段為2019 年12 月11—20 日(截止時間為最后一個觀測對象脫落),具體的觀測時間為上午8:00—12:00。
1.2 試驗方法
觀察觀測對象從花苞開裂至花苞脫落的過程,記錄其開花狀態、花瓣先端顏色、基部顏色、瓣爪顏色等,具體方法如下:
1)開花狀態:根據花瓣的質感和花朵的狀態,將開花的階段劃分為初綻、盛放、軟化、下垂和脫落。記錄數據并拍照。
2)花瓣先端顏色:使用英國皇家園林比色卡(Royal Horticultural Society, Sixth Edition 2015)記錄花瓣先端顏色,比色的位置為花瓣先端的頂部1/3 位置處。記錄數據并拍照。
3)花瓣基部:肉眼觀察花瓣基部(花瓣底部1/3 位置處)是否出現褐化,拍照并記錄數據。
4)瓣爪顏色:將花瓣中部位置的黃/白色區域定義為瓣爪區域,使用英國皇家園林比色卡記錄瓣爪顏色。黃色與白色的劃分以比色卡的顏色描述為準。最后記錄數據并拍照。
5)數據分析:本研究將對盛放狀態下的花朵的花瓣先端顏色進行聚類分析,因此需將比色卡RHS 顏色轉換成RGB 數值,在SPSS v.19 中進行系統聚類分析。RHS與RGB 的顏色轉換標準參照美國杜鵑花協會官方網站(https://www.azaleas.org/)提供的數據。其他數據使用excel 進行分析或圖表繪制。
2 結果與分析
2.1 花期分布情況
根據統計,超過80%的觀測對象花期為7~8 d,約95%的觀測對象花期為6~9 d(表1)。
2.2 開花階段變化規律
根據統計,95.4%的觀測對象均經歷了初綻、盛放、軟化、下垂、和脫落5 個階段(圖1)。所有花朵均在初綻當天即進入盛放階段,均在花期最后一天發生脫落。不同的開花階段情況如表2 所示。
2.3 花瓣顏色變化規律
108 個觀測對象的花瓣顏色均隨著開花時間的推移而有所變化。花瓣主要分為先端、瓣爪和基部3個顏色區域,三者的顏色均發生變化(圖2~3)。
2.3.1 花瓣先端顏色變化規律
以花朵盛放時的花瓣先端顏色為準,通過對觀測對象的花瓣先端顏色的聚類分析,可將主要花色劃分為粉白、粉紅和紫紅三大類別(圖4)。根據觀察,隨著開花時間推移,觀測對象均顯示出花瓣先端顏色發生變化的情況,變化趨勢均為逐漸弱化、淡化,如粉白色花的變化趨勢為粉白色逐漸淡化為白色;粉紅色花的顏色慢慢轉變為更淺的粉色;深紫紅色花則變化較廣,最終顏色為淡紫紅色或粉紅色。
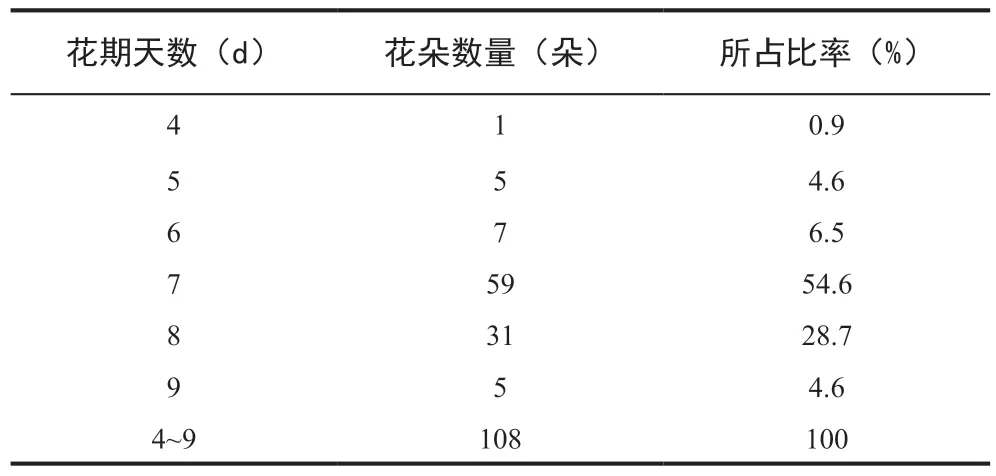
表1 觀測對象的花期情況

圖1 同一觀測對象在完整花期內不同開花階段的表現
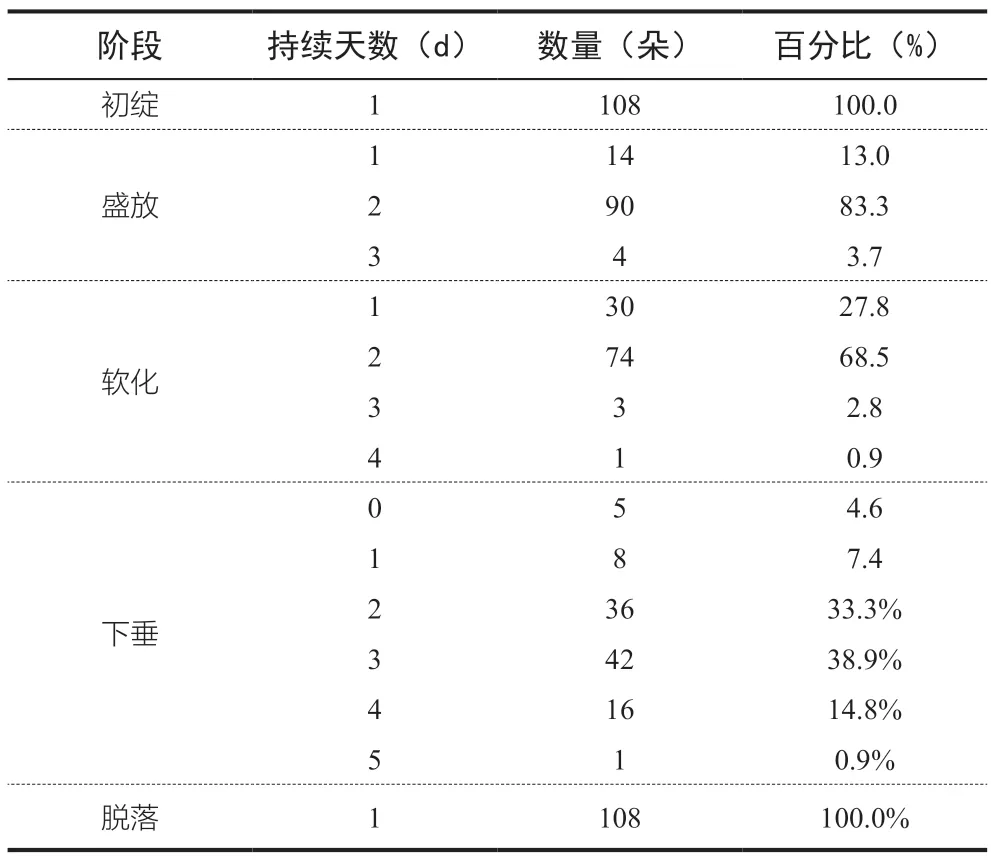
表2 觀測對象開花的各階段情況

圖2 開花前期的花瓣
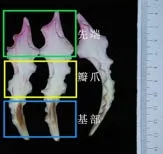
圖3 開花后期的花瓣
2.3.2 瓣爪顏色變化規律
據觀測,瓣爪顏色主要有黃色和白色2 種,所有觀測對象的瓣爪初始顏色均為黃色,隨著時間推移而逐漸淡化為白色。根據記錄,觀測對象的黃瓣爪時期均為開花后的1~3 d,較為集中;而白瓣爪時期的持續時間差異較大,為0~7 d(表3)。據統計,大部分觀測對象的黃瓣爪時期占其花期的20%~40%,平均黃瓣爪時期占比為30%;白瓣爪時期的占比則為50%~70%,平均白瓣爪時期占比為56%(圖5)。
2.3.3 花瓣基部顏色變化規律
在瓣爪顏色發生變化的同時,花瓣基部的顏色也發生改變。據觀察,觀測對象的花瓣基部均出現褐化現象,但花瓣基部開始出現褐化的時間不一。據統計,花瓣基部褐化通常在開花后的2~4 d 開始出現,出現的時間主要為第3 d(圖6)。結合瓣爪由黃色轉變為白色的時間節點,發現71%的觀測對象瓣爪顏色變為白色和花瓣基部開始褐化的時間節點重疊。當花瓣基部開始出現褐化之后,褐化(顏色)程度逐漸加深,同時褐化范圍逐漸由花瓣基部向瓣爪部位延伸,直至花瓣脫落。
2.3.4 不同開花階段的花瓣顏色變化
研究顯示,美麗異木棉復合體花瓣瓣爪和花瓣基部在開花階段中出現明顯的顏色變化,并且顏色變化與開花階段有較明顯的相關性,而花瓣頂端顏色變化相對不明顯。所有觀測對象的花瓣瓣爪在開花階段早期(初綻和盛放)呈現黃色,開花階段中期(軟化)顏色逐漸淡化,開花后期(下垂)完全變成白色。因此,觀測對象處于初綻和盛放階段時,花瓣瓣爪的顏色為黃色,軟化階段可能為黃色或白色,下垂階段則為白色。此外,所有觀測對象的花瓣基部在開花階段早期(初綻和盛放)沒有褐化,開花階段中期(軟化)逐漸褐化,開花后期(下垂)完全褐化。因此,觀測對象處于初綻和盛放階段時,花瓣基部沒有褐化,軟化階段為逐漸褐化,下垂階段則為完全褐化。

圖4 美麗異木棉復合體(盛放時)花瓣先端顏色的聚類分析

表3 觀測對象的瓣爪顏色變化情況
3 結論與討論
3.1 花期與開花階段
本研究涉及同一天開花的108 朵美麗異木棉復合體花朵在其完整花期內的動態變化,包括花期、開花狀態、花瓣先端顏色、瓣爪顏色、基部顏色等變化過程。研究結果表明,大部分美麗異木棉復合體花朵開花時間為6~9 d。在整個花期內,開花狀態是動態變化的,主要經歷了初綻、盛放、軟化、下垂和脫落5 個階段。少部分花朵缺失了部分狀態,如下垂階段,且其花期往往相對較短,因此推斷這些花朵很可能是在其他時間段(如當天下午或夜間)處于這些階段,但沒有被觀測到,在未來的進一步研究中需要增加下午和晚上的觀測時間。
3.2 花瓣顏色變化規律
本研究顯示,美麗異木棉復合體花瓣顏色存在差異,主要有2 個因素:1)花瓣固有的顏色差異,如盛放時期花瓣先端有粉白色、粉紅色、紫紅色三大類。2)花瓣自身演變,如花瓣先端顏色可由粉白色淡化為白色(淺灰色),深紫紅色逐漸轉為紫紅色,深紅色慢慢轉化為淺紅色等;花瓣基部顏色在花朵初綻時為黃色,隨著花期逐漸淡化為淺黃色,最終變為白色;花瓣基部逐漸褐化,并且褐色不斷加深,直至花朵脫落。
本次研究發現部分觀測對象(例如2、3、10 號)不僅花期長(9 d),而且瓣爪顏色變化時間長(黃瓣爪持續4 d),褐化時間晚(第4 天開始出現褐化)。此類型的個體更具有園林觀賞價值,可作為進一步的園林應用或育種材料。
3.3 花瓣顏色與開花階段的聯系
開花階段可由花瓣的瓣爪顏色和花瓣基部是否褐化進行預判:1)如瓣爪為黃色和花瓣基部沒有褐化,則觀測對象處于開花階段的初期—初綻和盛放階段;2)瓣爪為白色和花瓣基部已經褐化,表明觀測對象處于開花階段的后期—下垂階段;3)觀測對象處于軟化階段時,花瓣基部和瓣爪顏色處于上述描述的過渡狀態。
本研究發現,美麗異木棉復合體花朵顏色組成的復雜性不僅來源于花瓣先端固有的顏色差異,還有來源于花朵在花期內的動態變化。為挖掘美麗異木棉復合體的花朵狀態及顏色變化的本質及影響因素,后續可對該類群進行分子層面的分析如qPCR、轉錄組測序、基因組重測序等,以探討其變化的內在分子機制。
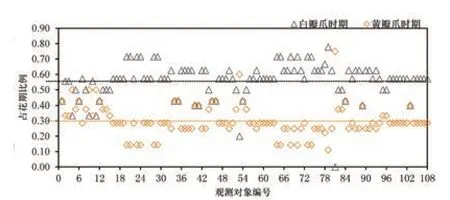
圖5 黃/白瓣爪時期占花期的比例情況

圖6 觀測對象花瓣基部褐化與瓣爪顏色變化時間的比較情況
