禾谷鐮孢核孔蛋白基因FgNup42的功能分析
2021-05-12 15:01:14張承啟廖露露齊永霞丁克堅陳莉
中國農業科學 2021年9期
張承啟,廖露露,齊永霞,丁克堅,陳莉
禾谷鐮孢核孔蛋白基因的功能分析
張承啟,廖露露,齊永霞,丁克堅,陳莉
安徽農業大學植物保護學院/作物有害生物綜合治理安徽省重點實驗室/植物病蟲害生物學與綠色防控安徽普通高校重點實驗室,合肥 230036
【】核孔蛋白Nup42在真核生物基因表達調控以及mRNA加工運輸等生物學過程中發揮著重要作用,本研究旨在分析禾谷鐮孢()中核孔蛋白基因在病原菌生長發育、逆境脅迫、致病和產毒等生物學過程中的功能。通過融合PCR(double-joint PCR)和酵母空隙修復(gap repair)技術分別構建基因敲除和回補載體,再利用PEG介導的原生質體轉化的方法獲得基因敲除突變體ΔFgNup42和回補體ΔFgNup42-C。觀察測定基因敲除突變體ΔFgNup42在營養生長、無性繁殖和有性生殖過程中的變化,同時測定突變體ΔFgNup42對滲透、殺菌劑以及細胞壁脅迫因子的敏感性。將突變體ΔFgNup42進行田間麥穗和室內玉米花絲接種試驗明確其致病力情況。通過液相色譜-串聯質譜(LC-MS/MS)檢測ΔFgNup42的產毒能力,同時利用qRT-PCR比較分析參與單端孢霉烯族毒素生物合成的7個在野生型PH-1和基因敲除突變體ΔFgNup42中的相對表達量。表型測定發現,基因敲除突變體ΔFgNup42的生長速率只有野生型PH-1的50%,菌落邊緣菌絲分枝變多且致密。顯微觀察分生孢子形成情況,發現敲除突變體ΔFgNup42相較野生型PH-1分生孢子產量降低了85.45%,并且隔膜數在0—2的分生孢子比例明顯增多。有性生殖誘導結果顯示ΔFgNup42的有性生殖能力增強,較野生型產生了更多的子囊殼。突變體ΔFgNup42對滲透脅迫因子NaCl和KCl,細胞壁脅迫因子剛果紅,以及殺菌劑戊唑醇和氰烯菌酯的敏感性減弱。致病力分析發現敲除基因后菌體在麥穗和玉米花絲上的致病力嚴重降低。此外,與野生型相比,ΔFgNup42中毒素DON、3ADON和15ADON的合成量明顯減少。核孔蛋白基因在禾谷鐮孢生長發育、抵御逆境以及致病和產毒過程中發揮著重要作用。
禾谷鐮孢;;生長發育;環境脅迫;致病力
0 引言
【研究意義】由禾谷鐮孢復合種(species complex)引起的赤霉病嚴重威脅麥類作物的生產安全[1-2],除了造成嚴重的產量損失外,病原菌還能夠在谷物籽粒中產生一系列的鐮刀菌毒素,包括脫氧雪腐鐮刀菌烯醇(deoxynivalenol,DON)及其乙酰化衍生物3ADON和15ADON,雪腐鐮刀菌烯醇(NIV)及其乙酰化衍生物玉米赤霉烯酮(zearalenone,ZEN),嚴重威脅食品安全和人畜生命健康[3-4]。自公布了禾谷鐮孢基因組數據以來[5],該病原菌中涉及致病、產毒相關基因的分子生物學功能得到了充分的解析,隨著組學層面的研究以及A-to-I的RNA編輯的發現,禾谷鐮孢的生長發育、致病和毒素生物合成調控機制得到了進一步的闡釋[6-9]。真核生物細胞功能的發揮取決于mRNA準確、有效地通過嵌在核膜上的核孔復合體從細胞核內輸出到細胞質中[10]。核孔復合體除了在細胞核與細胞質之間進行物質交換運輸外,該復合體中元件還扮演著與運輸無關的重要角色,包括基因表達調控、染色質組裝、DNA修復和mRNA加工等生物學過程[11]。解析核孔復合體中相關蛋白在禾谷鐮孢形態建成、致病以及毒素生物合成過程中的作用,對揭示該病原菌生長發育、抵御逆境、產毒和致病的分子機理,以及進一步探索小麥赤霉病的綜合防控技術具有重要意義。【前人研究進展】遺傳信息的順利傳遞需要新轉錄和加工的mRNA通過核孔復合體(nuclear pore complexes,NPC)從細胞核輸出到細胞質中。核孔復合體是貫穿核膜的大分子運輸機器,每個核孔復合體約由1 000個蛋白質亞基(統稱為核孔蛋白,nucleoporins)組成[12]。核孔復合體融合貫穿在核膜中形成孔道,并同時生成由富含苯丙氨酸-甘氨酸重復序列(phenylalanine-glycine repeats,FG)組成的被動擴散屏障;每個核孔復合體由約60 MDa的對稱核心組成,該核心在胞質面和核質面分別被不同的蛋白質進行修飾[13]。最早發現核孔復合體具有調控基因表達功能的試驗證據來自于釀酒酵母(),Menon等首次證明了核孔復合體中支架蛋白Nup84本身可以充當轉錄激活因子[14],隨后的研究發現釀酒酵母中其他的核孔蛋白也同樣參與了基因轉錄激活過程[15-16]。Nup42(又名Rip1)作為釀酒酵母中最早被鑒定的核孔蛋白之一,其與具有穿梭功能的RNA結合蛋白Rev的核輸出信號(nuclear export signal,NES)互作,促進了mRNA前體的剪接和輸出[17]。在釀酒酵母中,Nup42與mRNA輸出因子Gle1互作促使成熟的mRNA經過核孔,由具有ATPase酶活性的DEAD-box蛋白Dbp5水解ATP提供能量,進而運輸至細胞質[18-20]。在酵母和人類細胞中研究發現,Nup42具有兩個獨立功能的結構域,即FG結構域和CTD(carboxy-terminal domain)結構域,其中FG結構域與運輸受體Mex67-Mtr2互作招募成熟的mRNA[21-22];CTD結構域與mRNA輸出因子Gle1互作,可以向細胞質中運輸熱激蛋白成熟的轉錄本[23-24]。當人體細胞中同時缺失基因以及參與肌醇六磷酸合成的激酶基因時,熱激蛋白基因轉錄的mRNA則不能從細胞核輸出至細胞質中,從而導致細胞出現對溫度敏感的缺陷[25]。當在雙突變體ΔΔ中只轉入并表達Nup42的CTD結構域時,能夠實現熱激蛋白基因轉錄的mRNA正常輸出到細胞質中,并且恢復了突變體ΔΔ對溫度敏感的缺陷[25-26],但在細胞中只表達Nup42的FG結構域時并不能恢復突變體Δ對溫度敏感的缺陷[21]。關于核孔蛋白Nup42的功能研究目前僅限于人類、哺乳動物和酵母等模式生物細胞,在植物病原真菌中未見報道。【本研究切入點】禾谷鐮孢中核孔蛋白Nup42由基因(FGSG_06067)編碼,核孔蛋白在真核生物基因表達調控以及mRNA加工運輸等生物學過程中至關重要,但目前沒有相關研究報道禾谷鐮孢中核孔蛋白基因的生物學功能。【擬解決的關鍵問題】基于同源重組原理和PEG介導的原生質體轉化的方法,獲得禾谷鐮孢基因敲除突變體ΔFgNup42以及回補突變體ΔFgNup42-C,明確在禾谷鐮孢生長發育、抵抗逆境、產毒和致病等方面的作用,為進一步探究禾谷鐮孢致病和毒素生物合成調控提供一定的理論依據。
1 材料與方法
試驗于2018—2020年在安徽農業大學國家農作物品種審定特性鑒定站完成。
1.1 菌株及培養條件
禾谷鐮孢野生型菌株PH-1(NRRL 31084)、基因敲除突變體ΔFgNup42和回補突變體ΔFgNup42-C、質粒PBS、neo-PYF11、酵母菌株XK-125以及大腸桿菌DH5均保存在安徽農業大學國家農作物品種審定特性鑒定站。供試馬鈴薯瓊脂培養基(potato dextrose agar,PDA)、完全培養基(complete medium,CM)、基本培養基(minimal medium,MM)、分生孢子誘導培養基CMC(carboxymethyl cellulose)等的配制方法參考鐮孢菌試驗手冊(laboratory manual)[27]。
1.2 化學試劑和儀器
DNA聚合酶、反轉錄試劑盒、質粒小量提取試劑盒均購自南京諾唯贊生物科技有限公司;酵母轉化試劑盒購自MP Biomedicals公司;酵母質粒提取試劑盒購自Solarbio公司;潮霉素B購自Roche公司;引物、膠回收試劑盒、G418等常規生化試劑均采購于上海生工生物工程有限公司。
PCR儀,C1000 Touch,Bio-Rad公司;電泳儀,Powerpac HV,Bio-Rad公司;凝膠成像儀,Chemidoc,Bio-Rad公司;熒光倒置顯微鏡,Ti-S,尼康;移液器及臺式離心機,Eppendorf公司。
1.3 基因敲除及回補突變體的獲得
在禾谷鐮孢基因組數據庫網站(https://fungidb.org/fungidb/)檢索基因的上下游片段序列以及編碼區序列,并設計相關引物(表1)。首先PCR擴增出基因編碼區上下游序列各約800 bp,以質粒PBS為模板擴增潮霉素磷酸轉移酶基因();然后通過融合PCR(double-joint PCR)技術,按照上游、、下游的順序連接成一個重組DNA片段,從而構建出基因敲除打靶片段[28];最后利用同源重組原理和原生質體轉化的方法[29],將重組DNA片段轉入野生型PH-1中,在含有100 μg·mL-1潮霉素B的PDA培養基中篩選轉化子并用表1中的鑒定引物鑒定出敲除轉化子。
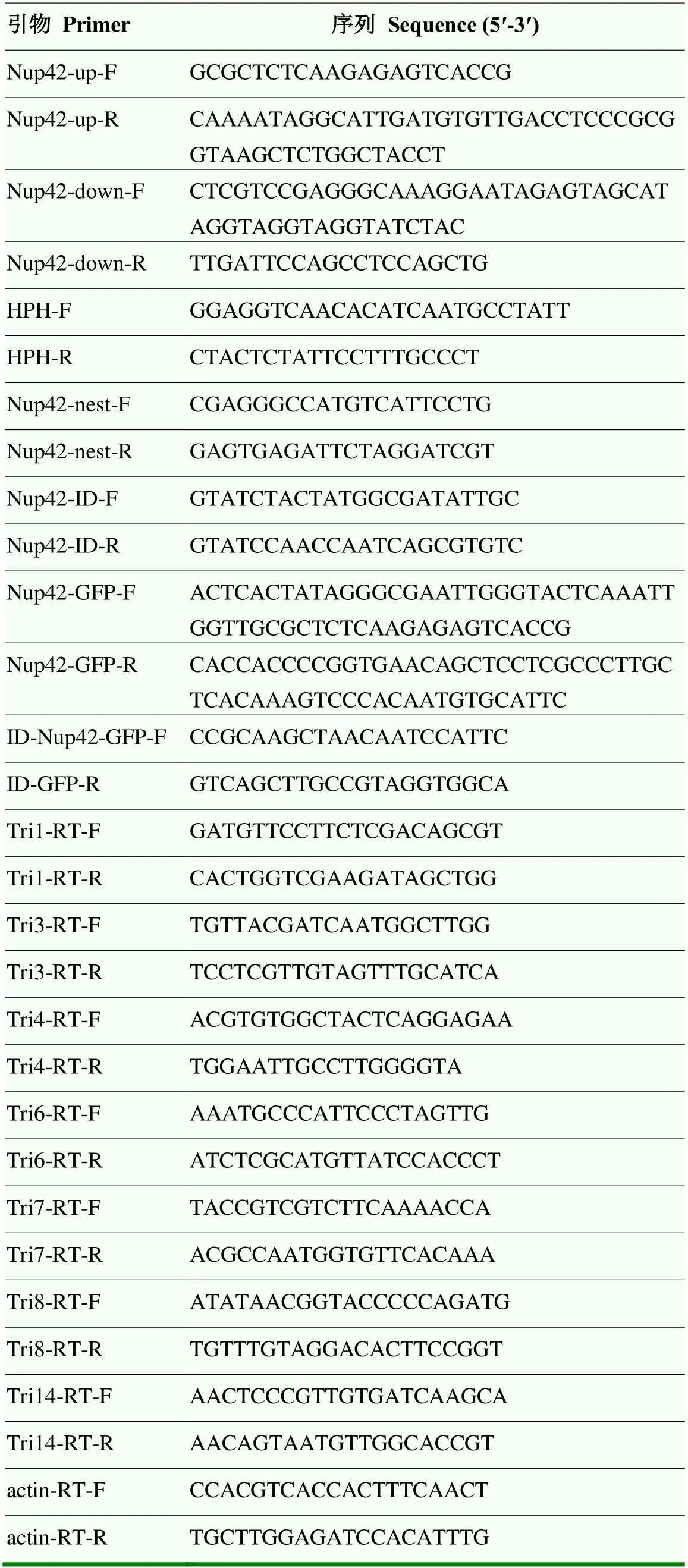
表1 本研究所用到的引物
利用表1中引物Nup42-GFP-F/Nup42-GFP-R擴增基因的啟動子至開放閱讀框的DNA片段,使用酵母轉化試劑盒將所得的PCR產物與經Ⅰ線性化的質粒neo-pYF11共轉化進入XK1-25酵母中構建重組質粒[30-31];然后,鑒定并提取陽性酵母菌株中的質粒并將其轉化至大腸桿菌DH5中;最后,提取大腸桿菌質粒經PEG介導的原生質體轉化至敲除突變體ΔFgNup42中,通過G418(100 μg·mL-1)篩選并經PCR鑒定回補轉化子。
1.4 菌落形態、無性繁殖及有性生殖觀測
菌落形態觀察:菌株于PDA平板上培養3 d,打孔器取直徑5 mm的菌碟分別接種于PDA、MM、CM的9 cm平板中,25℃培養3 d拍照,每次重復3皿,試驗重復3次。
邊緣菌絲觀察:將滅菌潔凈的載玻片浸潤在融化狀態的CM培養基中迅速取出放置暗盒中使其表面培養基凝固,取直徑3 mm的菌碟于載玻片上,每個菌株重復5個玻片,25℃培養16—20 h后取出載玻片于顯微鏡下觀察邊緣菌絲形態并拍照。
分生孢子產量及隔膜數統計:從新鮮活化的菌落邊緣打孔取5個直徑5 mm的菌碟接種到分裝有30 mL CMC培養液的50 mL三角瓶中,每個菌株3瓶,25℃,180 r/min搖培7 d。血球計數板統計孢子數量,試驗重復3次。吸取CMC培養液中的分生孢子,熒光增白劑(calcofluor white,CFW)染色,顯微鏡下統計不同隔膜數孢子的數量,每個菌株分別統計300個分生孢子。
有性生殖觀測:將菌碟接種于胡蘿卜培養基中,4個重復,25℃培養至所有菌株都長滿整個培養基,刮凈培養基表面的氣生菌絲,加入800 μL 0.1% Tween-20均勻涂抹在培養基表面,晾干后放置黑光燈下25℃培養20 d,觀察統計子囊殼數量及子囊孢子形態并拍照。
1.5 脅迫因子敏感性測定
從活化于PDA中生長3 d的菌落邊緣取直徑5 mm的菌碟分別接種于含1 mol·L-1NaCl、1 mol·L-1KCl、0.2 g·L-1剛果紅、0.25 μg·mL-1戊唑醇和0.25 μg·mL-1氰烯菌酯的PDA平板上,25℃培養3 d后拍照,每次重復3個皿,試驗重復3次。
1.6 致病力和毒素測定
麥穗接種:在小麥(感病品種安農8455)抽穗揚花盛期,注射10 μL孢子懸浮液(106孢子/mL)到麥穗中部小穗的花藥中,每天早晚各噴灌一次,15 d后統計發病情況并拍照記錄,每個菌株重復30株麥穗。
玉米花絲接種:將4根新鮮的玉米花絲彼此成一束,每束玉米花絲長度為10 cm,成束排列在經無菌蒸餾水潤濕的濾紙上。玉米花絲束中央放置一個直徑為5 mm的菌碟,5個重復,25℃保濕7 d,拍照。毒素的提取和測定方法參考Ji等[32]。
1.7 TRI表達分析
于PDA中生長3 d的菌落邊緣取5個直徑為5 mm的菌碟,置于分裝有30 mL誘導產毒培養基(trichothecene biosynthesis induction,TBI)的三角瓶中,錫箔紙包裹避光,25℃,180 r/min搖培48 h[33]。過濾收集TBI中的菌絲并在液氮中充分研磨,提取總RNA,使用反轉錄試劑盒進行逆轉錄。利用表1中相關引物,qRT-PCR確定的表達水平,試驗重復3次。
1.8 數據統計分析
采用Excel 2010計算菌落生長抑制率、分生孢子產量、子囊殼數量以及毒素含量,數據采用Fisher’s最小顯著差數法進行分析(=0.05)。
2 結果
2.1 敲除FgNup42對禾谷鐮孢營養生長的影響
基因缺失突變體ΔFgNup42在培養基PDA、MM和CM上的生長速率與野生型PH-1、回補突變體ΔFgNup42-C相比降低了約50%(圖1-a、1-b)。通過觀察菌落邊緣形態特征發現,敲除突變體ΔFgNup42相較野生型,邊緣菌絲分枝變多且致密(圖1-c),結果表明的缺失嚴重影響了禾谷鐮孢的生長速率以及菌落的形態特征。
2.2 FgNup42參與禾谷鐮孢分生孢子的發育
經熒光增白劑(CFW)染色,顯微鏡觀察誘導產孢培養基CMC中分生孢子的形態發現,基因敲除突變體ΔFgNup42產生的分生孢子隔膜數與野生型相比明顯變少(圖2-a);通過統計分析不同隔膜數分生孢子所占的比例發現,突變體ΔFgNup42中0—2個隔膜的分生孢子比例較野生型和回補體明顯增加(圖2-b)。血球計數板統計各菌株的分生孢子產量發現,敲除突變體ΔFgNup42的產孢量顯著降低,只有野生型PH-1的14.55%(圖2-c)。

a:PH-1、敲除突變體ΔFgNup42和回補體ΔFgNup42-C在培養基PDA、CM和MM中25℃生長3 d PH-1, ΔFgNup42 deletion mutant and ΔFgNup42-C complemented strains were grown on PDA, CM and MM at 25℃ for 3 d;b:菌株PH-1、ΔFgNup42和ΔFgNup42-C在PDA培養基中的生長速率比較Comparison of mycelial growth rates among PH-1, ΔFgNup42 and ΔFgNup42-C strains on PDA medium;c:PH-1、ΔFgNup42和ΔFgNup42-C的菌落邊緣菌絲生長情況Hyphal growth at the edges of PH-1, ΔFgNup42 and ΔFgNup42-C colonies
2.3 敲除基因FgNup42對有性生殖的影響
在禾谷鐮孢侵染小麥的過程中,子囊孢子作為重要的初侵染源在病害循環中發揮著至關重要的作用[34]。因此,筆者觀測了野生型PH-1、基因缺失突變體ΔFgNup42以及回補突變體ΔFgNup42-C子囊殼以及子囊孢子的形成狀況。經過20 d的有性生殖誘導,敲除突變體ΔFgNup42產生的子囊殼數量顯著多于野生型和回補體(圖3-a、3-b)。于載玻片上壓開子囊殼后觀察發現,野生型PH-1、敲除突變體ΔFgNup42和回補突變體ΔFgNup42-C在子囊以及子囊孢子的形態特征上沒有明顯差異(圖3-c),說明基因的缺失不影響子囊以及子囊孢子的形態。
2.4 FgNup42參與禾谷鐮孢對滲透、細胞壁以及殺菌劑脅迫的敏感性
Nup42作為細胞核孔的重要組分,而核孔又是物質進出細胞核與細胞質的重要通道[13],因此測定了基因敲除突變體ΔFgNup42對滲透脅迫(1 mol·L-1NaCl和1 mol·L-1KCl)、細胞壁脅迫(0.2 g·L-1剛果紅)以及殺菌劑(0.25 μg·mL-1戊唑醇和0.25 μg·mL-1氰烯菌酯)的敏感性,發現ΔFgNup42相較野生型對NaCl、KCl、剛果紅、戊唑醇以及氰烯菌酯的抗性顯著增加(圖4)。
2.5 FgNup42的缺失對禾谷鐮孢致病和產毒的影響
為了明確在禾谷鐮孢致病過程中的功能,對野生型PH-1、敲除突變體ΔFgNup42以及回補突變體ΔFgNup42-C在麥穗和玉米花絲上進行了致病力測定。田間麥穗接種發現突變體ΔFgNup42的致病力顯著下降,僅局限于小穗發病(圖5-a);通過室內接種玉米花絲發現,ΔFgNup42在玉米花絲上的侵染擴展程度相比野生型和回補體同樣明顯下降(圖5-b)。DON毒素及其衍生物是禾谷鐮孢的關鍵致病因子[35],由于敲除基因后導致病原菌致病力嚴重下降,因此筆者測定了PH-1、ΔFgNup42以及ΔFgNup42-C的產毒能力。結果顯示,ΔFgNup42產生的毒素量DON、3ADON以及15ADON顯著低于野生型和回補菌體(圖5-c)。為了進一步證實該結果,測定了參與DON毒素生物合成途徑中各的表達量,與野生型相比,ΔFgNup42中7個基因的轉錄水平明顯降低(圖5-d)。上述結果表明,FgNup42通過調節禾谷鐮孢中的表達來調控毒素的生物合成。

a:PH-1、ΔFgNup42和ΔFgNup42-C在添加1 mol·L-1 NaCl、1 mol·L-1 KCl、0.2 g·L-1剛果紅、0.25 μg·mL-1戊唑醇和0.25 μg·mL-1氰烯菌酯的PDA培養基中25℃生長3 d的比較Comparison of PH-1, ΔFgNup42 and ΔFgNup42-C following incubation at 25℃ for 3 d on PDA plates supplemented with 1 mol·L-1NaCl, 1 mol·L-1KCl, or 0.2 g·L-1Congo red, 0.25 μg·mL-1 tebuconazole and 0.25 μg·mL-1phenamacril;b:菌株在含有NaCl、KCl、剛果紅、戊唑醇和氰烯菌酯的PDA培養基中生長3 d相較對照組的菌絲生長抑制率Mycelial growth inhibition compared with non-treated controls following incubation for 3 d on PDA containing NaCl, KCl, Congo red, tebuconazole and phenamacril
3 討論
核孔復合體控制著生物大分子在細胞核與細胞質之間的運輸,在mRNA輸出至細胞質的最后一步,DEAD-box解旋酶Dbp5(人類細胞中叫做DDX19)被核孔蛋白Nup42、Gle1和Nup214激活,可從mRNA顆粒中去除RNA結合蛋白Nxf1·Nxt1,從而使成熟的mRNA順利到達細胞質完成翻譯[13]。編碼一個核孔蛋白,該蛋白最早被鑒定到與釀酒酵母中表達的HIV-1 Rev的效應子互作[17,36]。雖然有關Nup42的生化功能在釀酒酵母和人類細胞中研究得比較透徹[11,37],但是有關分子遺傳學方面的功能尚未在植物病原真菌中研究報道。
本研究發現禾谷鐮孢基因缺失突變體ΔFgNup42生長速率與野生型相比明顯減慢,但是在釀酒酵母中并非其生長所必需[36]。此外,突變體ΔFgNup42在無性繁殖過程中出現嚴重缺陷,分生孢子產量顯著減少并且不同隔膜的分生孢子比例分布發生了明顯的分化。但ΔFgNup42的有性生殖能力增強了,相較野生型菌株產生了更多的子囊殼,同時子囊孢子的形態正常,可能因為敲除基因致使禾谷鐮孢有絲分裂相關基因的表達受到抑制,而減數分裂過程得到了促進,此現象背后的調控機制有待進一步研究。突變體ΔFgNup42對外界滲透壓、細胞壁脅迫以及殺菌劑的敏感性發生了變化,禾谷鐮孢參與菌體抵御外界環境脅迫,很可能由于基因的缺失導致相關信號通路的轉錄因子Hog1[38]、Slt2[39]、FgSR[40]以及FgTfml[41]從細胞質進入細胞核受阻,這與Nup42在釀酒酵母中同源基因缺失突變體Δ抵抗逆境脅迫因子類似[11,26]。除了在生長發育過程中出現的缺陷外,ΔFgNup42在寄主植物上的致病力嚴重降低,雖然敲除了基因后使菌體的生長速率減慢了50%,但是突變體ΔFgNup42在麥穗上的致病僅局限于單個小穗,沒有向周圍小穗擴展,說明ΔFgNup42致病力的缺陷并不是由生長速率降低引起的。通過測定菌株產毒能力的變化發現,ΔFgNup42在發病麥粒中3種毒素(DON、3ADON和15ADON)合成量顯著減少,而毒素是禾谷鐮孢關鍵的致病因子,幫助其在寄主中擴展蔓延[35,42],因此可以推斷ΔFgNup42致病力的降低主要是其毒素合成能力的下降所導致的。突變體ΔFgNup42中參與DON毒素生物合成的表達量的顯著降低更加證實了其毒素合成能力的缺陷,關于核孔蛋白FgNup42調控DON毒素生物合成的轉錄及出核運輸機制有待于進一步研究。

a:PH-1、ΔFgNup42和ΔFgNup42-C在麥穗上的發病癥狀Disease symptoms on wheat heads caused by PH-1, ΔFgNup42 and ΔFgNup42-C;b:PH-1、ΔFgNup42和ΔFgNup42-C接種玉米花絲致病情況Maize silks inoculated with PH-1, ΔFgNup42 and ΔFgNup42-C;c:各菌株于接種侵染的麥穗籽粒中單端孢霉烯族毒素的產量Levels of trichothecene mycotoxins produced by each strain in infected spikelets collected from inoculated wheat heads;d:7個TRI在PH-1和ΔFgNup42中的相對表達量Relative transcription levels of 7 TRIs in PH-1 and ΔFgNup42
4 結論
通過基因敲除與回補的方法研究了禾谷鐮孢中基因的功能,的缺失導致禾谷鐮孢生長速率降低,分生孢子產量減少,有性生殖能力變強,對滲透、細胞壁以及殺菌劑的脅迫敏感性變弱,毒素合成能力下降同時致病力嚴重降低。
[1] 陳云, 王建強, 楊榮明, 馬忠華. 小麥赤霉病發生危害形勢及防控對策. 植物保護, 2017, 43(5): 11-17.
CHEN Y, WANG J Q, YANG R M, MA Z H. Current situation and management strategies of Fusarium head blight in China. Plant Protection, 2017, 43(5): 11-17. (in Chinese)
[2] MCMULLEN M, JONES R, GALLENBERG D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Disease, 1997, 81(12): 1340-1348.
[3] PESTKA J J, SMOLINSKI A T. Deoxynivalenol: Toxicology and potential effects on humans. Journal of Toxicology and Environmental Health, Part B Critical Reviews, 2005, 8(1): 39-69.
[4] 史建榮, 劉馨, 仇劍波, 祭芳, 徐劍宏, 董飛, 殷憲超, 冉軍艦. 小麥中鐮刀菌毒素脫氧雪腐鐮刀菌烯醇污染現狀與防控研究進展. 中國農業科學, 2014, 47(18): 3641-3654.
SHI J R, LIU X, QIU J B, JI F, XU J H, DONG F, YIN X C, RAN J J. Deoxynivalenol contamination in wheat and its management. Scientia Agricultura Sinica, 2014, 47(18): 3641-3654. (in Chinese)
[5] CUOMO C A, GUELDENER U, XU J R, TRAIL F, TURGEON B G, DI PIETRO A, WALTON J D, MA L J, BAKER S E, REP M,. Thegenome reveals a link between localized polymorphism and pathogen specialization. Science, 2007, 317(5843): 1400-1402.
[6] SON H, SEO Y S, MIN K, PARK A R, LEE J, JIN J M, LIN Y, CAO P, HONG S Y, KIM E K,. A phenome-based functional analysis of transcription factors in the cereal head blight fungus,. Plos Pathogens, 2011, 7(10): e1002310.
[7] WANG C F, ZHANG S J, HOU R, ZHAO Z T, ZHENG Q, XU Q J, ZHENG D W, WANG G H, LIU H Q, GAO X L, MA J W, KISTLER H C, KANG Z S, XU J R. Functional analysis of the kinome of the wheat scab fungus. Plos Pathogens, 2011, 7(12): e1002460.
[8] YUN Y Z, LIU Z Y, YIN Y N, JIANG J H, CHEN Y, XU J R, MA Z H. Functional analysis of thephosphatome. New Phytologist, 2015, 207(1): 119-134.
[9] LIU H, WANG Q H, HE Y, CHEN L F, HAO C F, JIANG C, LI Y, DAI Y F, KANG Z S, XU J R. Genome-wide A-to-I RNA editing in fungi independent of ADAR enzymes. Genome Research, 2016, 26(4): 499-509.
[10] CASEY A K, WENTE S R. Nuclear transport: shifting gears in fungal nuclear and cytoplasmic organization. Current Biology, 2012, 22(19): R846-R848.
[11] RAICES M, D’ANGELO M A. Nuclear pore complexes and regulation of gene expression. Current Opinion in Cell Biology, 2017, 46: 26-32.
[12] LIN D H, HOELZ A. The structure of the nuclear pore complex (An update). Annual Review of Biochemistry, 2019, 88: 725-783.
[13] LIN D H, CORREIA A R, CAI S W, HUBER F M, JETTE C A, HOELZ A. Structural and functional analysis of mRNA export regulation by the nuclear pore complex. Nature Communications, 2018, 9: 2319.
[14] MENON B B, SARMA N J, PASULA S, DEMINOFF S J, WILLIS K A, BARBARA K E, ANDREWS B, SANTANGELO G M. Reverse recruitment: The Nup84 nuclear pore subcomplex mediates Rap1/ Gcr1/Gcr2 transcriptional activation. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(16): 5749-5754.
[15] CASOLARI J M, BROWN C R, KOMILI S, WEST J, HIERONYMUS H, SILVER P A. Genome-wide localization of the nuclear transport machinery couples transcriptional status and nuclear organization. Cell, 2004, 117(4): 427-439.
[16] SCHMID M, ARIB G, LAEMMLI C, NISHIKAWA J, DURUSSEL T, LAEMMLI U K. Nup-PI: The nucleopore-promoter interaction of genes in yeast. Molecular Cell, 2006, 21(3): 379-391.
[17] STUTZ F, NEVILLE M, ROSBASH M. Identification of a novel nuclear pore-associated protein as a functional target of the HIV-1 Rev protein in yeast. Cell, 1995, 82(3): 495-506.
[18] MURPHY R, WENTE S R. An RNA-export mediator with an essential nuclear export signal. Nature, 1996, 383(6598): 357-360.
[19] BOLGER T A, FOLKMANN A W, TRAN E J, WENTE S R. The mRNA export factor Gle1 and inositol hexakisphosphate regulate distinct stages of translation. Cell, 2008, 134(4): 624-633.
[20] GROSS T, SIEPMANN A, STURM D, WINDGASSEN M, SCARCELLI J J, SEEDORF M, COLE C N, KREBBER H. The DEAD-box RNA helicase Dbp5 functions in translation termination. Science, 2007, 315(5812): 646-649.
[21] ADAMS R L, TERRY L J, WENTE S R. Nucleoporin FG domains facilitate mRNP remodeling at the cytoplasmic face of the nuclear pore complex. Genetics, 2014, 197(4): 1213-1224.
[22] STRASSER K, BASSLER J, HURT E. Binding of the Mex67p/Mtr2p heterodimer to FXFG, GLFG, and FG repeat nucleoporins is essential for nuclear mRNA export. The Journal of Cell Biology, 2000, 150(4): 695-706.
[23] KENDIRGI F, REXER D J, ALCAZAR-ROMAN A R, ONISHKO H M, WENTE S R. Interaction between the shuttling mRNA export factor Gle1 and the nucleoporin hCG1: A conserved mechanism in the export of Hsp70 mRNA. Molecular Biology of the Cell, 2005, 16(9): 4304-4315.
[24] SAAVEDRA C A, HAMMELL C M, HEATH C V, COLE C N. Yeast heat shock mRNAs are exported through a distinct pathway defined by Rip1p. Genes and Development, 1997, 11(21): 2845-2856.
[25] MILLER A L, SUNTHARALINGAM M, JOHNSON S L, AUDHYA A, EMR S D, WENTE S R. Cytoplasmic inositol hexakisphosphate production is sufficient for mediating the Gle1-mRNA export pathway. The Journal of Biological Chemistry, 2004, 279(49): 51022-51032.
[26] STUTZ F, KANTOR J, ZHANG D, MCCARTHY T, NEVILLE M, ROSBASH M. The yeast nucleoporin rip1p contributes to multiple export pathways with no essential role for its FG-repeat region. Genes and development, 1997, 11(21): 2857-2868.
[27] NICHOLSON P. Thelaboratory manual. Plant Pathology, 2007, 56(6): 1037.
[28] YU J H, HAMARI Z, HAN K H, SEO J A, REYES-DOMINGUEZ Y, SCAZZOCCHIO C. Double-joint PCR: a PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genetics and Biology, 2004, 41(11): 973-981.
[29] PROCTOR R H, HOHN T M, MCCORMICK S P. Reduced virulence ofcaused by disruption of a trichothecene toxin biosynthetic gene. Molecular Plant-Microbe Interactions, 1995, 8(4): 593-601.
[30] BRUNO K S, TENJO F, LI L, HAMER J E, XU J R. Cellular localization and role of kinase activity ofin. Eukaryotic Cell, 2004, 3(6): 1525-1532.
[31] CHEN Y, ZHENG S Y, JU Z, ZHANG C Q, TANG G F, WANG J, WEN Z Y, CHEN W, MA Z H. Contribution of peroxisomal docking machinery to mycotoxin biosynthesis, pathogenicity and pexophagy in the plant pathogenic fungus. Environmental Microbiology, 2018, 20(9): 3224-3245.
[32] JI F, XU J H, LIU X, YIN X C, SHI J R. Natural occurrence of deoxynivalenol and zearalenone in wheat from Jiangsu province, China. Food Chemistry, 2014, 157: 393-397.
[33] MENKE J, DONG Y H, KISTLER H C.Tri12p influences virulence to wheat and trichothecene accumulation. Molecular Plant-Microbe Interactions, 2012, 25(11): 1408-1418.
[34] TRAIL F. For blighted waves of grain:in the postgenomics era. Plant Physiology, 2009, 149(1): 103-110.
[35] SEONG K Y, PASQUALI M, ZHOU X, SONG J, HILBURN K, MCCORMICK S, DONG Y, XU J R, KISTLER H C. Global gene regulation bytranscription factorsandreveals adaptations for toxin biosynthesis. Molecular Microbiology, 2009, 72(2): 354-367.
[36] FABRE E, HURT E. Yeast genetics to dissect the nuclear pore complex and nucleocytoplasmic trafficking. Annual Review of Genetics, 1997, 31: 277-313.
[37] ADAMS R L, MASON A C, GLASS L, ADITI, WENTE S R. Nup42 and IP6coordinate Gle1 stimulation of Dbp5/DDX19B for mRNA export in yeast and human cells. Traffic, 2017, 18(12): 776-790.
[38] ZHENG D W, ZHANG S J, ZHOU X Y, WANG C F, XIANG P, ZHENG Q, XU J R. Thepathway regulates hyphal growth, stress responses, and plant infection in. Plos One, 2012, 7(11): e49495.
[39] YUN Y Z, LIU Z Y, ZHANG J Z, SHIM W B, CHEN Y, MA Z H. The MAPKK FgMkk1 ofregulates vegetative differentiation, multiple stress response, and virulence via the cell wall integrity and high-osmolarity glycerol signaling pathways. Environmental Microbiology, 2014, 16(7): 2023-2037.
[40] LIU Z Y, JIAN Y Q, CHEN Y, KISTLER H C, HE P, MA Z H, YIN Y N. A phosphorylated transcription factor regulates sterol biosynthesis in. Nature Communications, 2019, 10: 1228.
[41] LIU N, WU S Q, DAWOOD D H, TANG G F, ZHANG C Q, LIANG J T, CHEN Y, MA Z H. The b-ZIP transcription factor FgTfmI is required for the fungicide phenamacril tolerance and pathogenecity in. Pest Management Science, 2019, 75(12): 3312-3322.
[42] BAI G H, DESJARDINS A E, PLATTNER R D. Deoxynivalenol- nonproducingcauses initial infection, but does not cause disease spread in wheat spikes. Mycopathologia, 2002, 153(2): 91-98.
Functional analysis of the nucleoporin gene
ZHANG Chengqi, LIAO Lulu, QI Yongxia, DING Kejian, CHEN Li
School of Plant Protection, Anhui Agricultural University/Anhui Province Key Laboratory of Integrated Pest Management on Crops/Key Laboratory of Biology and Sustainable Management of Plant Diseases and Pests of Anhui Higher Education Institutes, Hefei 230036
【】The nucleoporin Nup42 plays important roles in gene expression regulation, nuclear mRNA processing and export. The objective of this study is to analyze the function of the nucleoporin genegrowth and development, stress, pathogenicity and mycotoxins production of.【】Targeted gene deletion and mutant complementation constructs were generated using the double-joint PCR and yeast gap repair methods, respectively. Then the gene deletion mutant ΔFgNup42 and complemented strain ΔFgNup42-C were obtained by PEG-mediated protoplast transformation. The vegetative growth, asexual and sexual reproduction of the gene disrupted mutant ΔFgNup42 were examined. Moreover, the sensitivity of ΔFgNup42 to osmotic stress, fungicides and cell wall damaging agent were investigated. Pathogenicity of the mutant ΔFgNup42 was assayed by point-inoculated the flowering wheat heads and incubation on maize silks. Mycotoxins produced by ΔFgNup42 were quantified using the liquid chromatography-tandem mass spectrometry (LC-MS/MS) method. The relative transcription levels of 7s involved in trichothecene mycotoxins biosynthesis were determined by qRT-PCR between PH-1 and ΔFgNup42.【】Phenotype analysis showed that the growth rate of ΔFgNup42 was 50% of that of wild type PH-1, but more dense in hyphal branching. Microscopic examination revealed that the conidiation of ΔFgNup42 mutant decreased by 85.45% compared with the wild type PH-1, and percentage of conidia with septum range 0 to 2 was significantly increased. The ΔFgNup42mutant produced more perithecia than the wild type strain in mating cultures suggesting improved sexual reproduction. The mutant ΔFgNup42displayed increased tolerance to osmotic stress NaCl and KCl, fungicides tebuconazole and phenamacril, also cell wall damaging agent Congo red. Pathogenicity assays exhibited that disruption of genecaused a dramatic reduction in pathogenicity by inoculated the flowering wheat heads and maize silks. Furthermore, the amount of mycotoxins DON, 3ADON and 15ADON produced by ΔFgNup42 were significantly lower than that produced by wild type.【】The nucleoporin geneplays a key role in fungal growth and development, pathogenesis, mycotoxins production and responses to various environmental stresses in
;; growth and development; environmental stress; pathogenicity

10.3864/j.issn.0578-1752.2021.09.007
2020-09-14;
2020-09-24
國家重點研發計劃(2018YFD0200500)
張承啟,E-mail:zhcq@ahau.edu.cn。通信作者陳莉,E-mail:chenlii@ahau.edu.cn
(責任編輯 岳梅)
