魚-菜共生養殖模式對羅非魚生長性能、水質和浮游生物的影響
2021-05-06 13:06:53劉邦輝方彰勝丁悅秀
江蘇農業科學 2021年6期
劉邦輝 方彰勝 丁悅秀
摘要:以尼羅羅非魚為研究對象,采用魚-菜共生養殖技術對6口池塘(A組:試驗組,3口池塘;B組:對照組,3口池塘)羅非魚生長性能、水質影響和養殖效益進行研究。結果顯示,在196 d養殖周期內,魚-菜共生模式(A組)下羅非魚的生長速度、成活率、肥滿度分別顯著提高18.9%、13.3%和37.0%(P<0.05),飼料系數降低10.6%(P<005);在水質改良方面,試驗組COD、濁度、亞硝酸鹽、總氨氮及溶解性正磷酸鹽的含量與對照組(B組)相比均具有顯著差異(P<005),A組池塘浮游植物密度和浮游動物密度均顯著低于B組(P<0.05),但浮游動物生物量顯著高于B組(P<005)。同時,A組浮游植物Shannon-Weaver多樣性指數(H)和Pielou均勻度指數(J)均顯著高于對照池塘(P<005)。表明魚-菜共生養殖技術,具有提高羅非魚生長性能、提高養殖池塘浮游生物多樣性、改善養殖水質、降低水體污染和增產增值等優點,不失為一種生態健康高效的精準池塘養殖模式。
關鍵詞:羅非魚;魚菜共生;精準池塘;養殖模式
多年來,作為傳統的池塘養殖國家,我國池塘養殖面積和水產品產量均居世界第一[1],依靠技術進步和模式升級,高密度集約化養殖已成為我國池塘養殖的主要養殖模式[2-3]。隨之產生大量的池塘沉積有機物,進而致使池塘水體呈富營養態,排出后甚至會導致附近水源或水資源惡化[4-5]。因此,探索具有增值、高效、環保的新型養殖模式以維持水產養殖增產生態協同發展已成為近年來科技工作者新的研究課題[6-7]。
魚-菜共生系統是指將高密度水產養殖與水耕栽培(hydroponics)有機結合,形成生物浮床,利用水生植物生長過程中需要吸收大量氮、磷等元素的原理,達到去除或降低池塘水體中的沉余有機物含量以改善水質收獲蔬菜的目的,進而形成營養鹽良性循環的養殖-種植系統[8]。已有研究表明,魚-菜共生可以使養殖生物、植物和微生物等系統生物與生態因子達到一種平衡狀態,促使養殖水體呈現穩態,進而為養殖水體的原位修復提供有效途徑[9-10]。
羅非魚是我國重要的池塘養殖經濟魚類之一,其肉質鮮美、刺少、蛋白質含量高,近年已成為我國養殖、加工、出口的優質魚類[11]。本研究以羅非魚為研究對象,采用魚菜共生-生物浮床技術對6口池塘(A組:試驗組;B組:對照組)羅非魚生長性能、水質和浮游生物進行研究,以期為魚菜共生-生物浮床技術在水產養殖中的應用提供技術參數。
1 材料與方法
1.1 試驗材料
試驗在廣東省江門市新會區開源農業有限公司羅非魚養殖場進行,時間為2018年4月17日至10月30日。試驗分為2組,每組3個重復,總計池塘6口,每口池塘面積為2 000 m2,養殖水源為西江水,池塘水深為1.8~2.0 m,所有試驗養殖池塘均配置2臺葉輪式增氧機,功率為1.5 kW。試驗用魚為佛山市梁氏集團提供的無病無傷尼羅羅非魚(Oreochromis niloticus),平均體質量為(20.0±12) g/尾,試驗用菜為江門市新會市開源農業有限公司提供的蕹菜(Ipomoea aquatica Forsk.),又稱空心菜。
1.2 試驗方法
試驗分為2個組、各3個重復試驗(A組、試驗組;B組、對照組),每組試驗魚放養規格和密度分別為(20±1.2)g/尾和(19.0±0.1)×104尾/hm2,組間無顯著差異。其中,A組池塘水面每隔4 m放置2 m寬浮排結構的裝置并種植蕹菜(圖1、圖2),共6排,B組池塘無放置。
1.2.1 養殖管理 試驗期間,尼羅羅非魚養殖、投料及日常管理參照開源農業有限公司養殖場羅非魚池塘養殖企業標準進行,試驗飼料由恒興飼料提供,每天06:00—08:00、14:00—18:00各投喂飼料1次,投喂量為魚質量的3%,試驗周期為126 d。試驗期間,記錄各池塘放養規格、放養量、飼料投喂量等。試驗完成時,統計魚體規格、總質量、成活率、飼料利用率等。
1.2.2 養殖水質因子測定 每周一07:00—08:00采用ET 9908便攜式多參數水質測定儀對試驗養殖水體的溫度(T)、pH值、溶解氧(DO)含量、濁度(Tur)進行定點測定;透明度(Scc)采用薩氏盤法進行測定。采集各試驗池塘30 cm處水體用于水質因子測定,其中,采用納氏試劑分光光度法(HJ 535—2009)測定TAN含量;采用分光光度法(GB 7493—1987)測定NO-2含量;采用紫外分光光度法(HJ/T 346—2007)測定NO-3含量;采用重鉻酸鹽法(GB 11914—1989)測定COD含量;采用磷鉬藍分光光度法(HJ593—2010)測定溶解性正磷酸鹽(SRP)含量。
1.2.3 浮游生物水樣采集與測定 浮游生物的采樣時間和次數與水質因子測定水樣一致。采用《內陸水域漁業自然資源調查手冊》方法進行浮游生物采樣、計數及種類鑒定。采集前配制魯哥氏液用以固定水樣。在各試驗池塘選擇5個采樣點(2對角線上選擇4個點和中心點)分別采集1 000 mL水體,取500 mL混合水樣用于測定浮游植物、原生動物和輪蟲等等浮游生物量,取1 000 mL混合水樣經浮游生物過濾網過濾后用于枝角類和橈足類測定。所有樣品經沉淀濃縮后在顯微鏡下觀察計數。浮游生物多樣性按以下公式計算:
1.3 計算公式
1.4 數據處理
試驗數據均采用SPSS 17.0處理,采用“平均值±標準差”表示。在單因子方差分析(A-NOVA)基礎上采用t檢驗進行分析比較,取95%置信區間(P<0.05)或99%置信區間(P<0.01)。
2 結果與分析
2.1 羅非魚生長特性及養殖效益分析
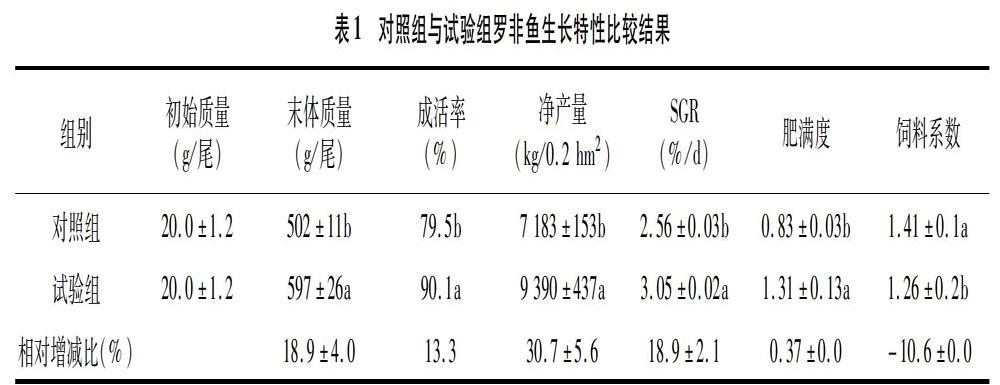
由表1可知,與對照組相比,試驗組的特定生長率、肥滿度、成活率和凈產量分別提高了18.90%、0.37%、13.30%和30.70%,飼料系數降低了1060%,均呈現顯著差異(P<0.05)。魚-菜共生養殖設施費用0.3萬元,使用周期為3年,平均每年折舊費0.1萬元。試驗組比對照組凈增值0.5萬元/1 000 m2(魚時價9.8 元/kg,飼料時價6.2元/kg)。結果顯示,魚-菜共生養殖技術不僅可提高羅非魚養殖成活率和特定生長率,還可顯著提高飼料利用率進而促進增產增值,不失為一種生態、高效的養殖方式。
2.2 池塘水質因子濃度變化
由表2可知,試驗周期內,與對照組相比,試驗組養殖水體水溫和硝酸鹽平均濃度無顯著差異(P>0.05),試驗組養殖水體pH值、Scc、DO等水環境因子數值均顯著高于對照組(P<0.05),試驗組養殖水體Tur、TAN、NO-2、COD和SRP等水質因子分別降低了34.34%、18.31%、52.24%、39.7%和38.69%,也均呈現出顯著差異(P<0.05)。試驗結果表明,與對照組相比,試驗組在相同飼養管理的前提下,魚-菜共生對羅非魚池塘養殖水體水質因子濃度水平的調節和生態系統的穩定具有較高的效率。
2.3 浮游生物種類組成及百分比
在養殖試驗周期內,通過對池塘養殖水體浮游生物種類的鑒定,大致檢測出浮游動物24種(屬),其中原生動物為10種(屬)、輪蟲為10種(屬)、枝角類為2種(屬)、橈足類為2種(屬)。檢測出浮游植物48種(屬),其中綠藻門為19種(屬)、硅藻門為12種(屬)、藍藻門為8種(屬)、裸藻門為4種(屬)、隱藻門為2種(屬)、甲藻門為3種(屬),2組池塘的優勢種群基本相同。
通過對2組池塘養殖水體浮游生物統計(表3)得知,2組羅非魚養殖池塘內的浮游植物均以藍藻、綠藻和硅藻為主,3種藻類總和分別占浮游植物的88.2%和89.1%,在各組藻間除綠藻、甲藻百分比無顯著差異外(P>0.05),其余藻類均有顯著差異(P<005);浮游動物以原生動物為主,試驗組和對照組分別占總浮游生物數量的91.1%和85.4%,在各組原生動物間,除原生動物和枝角類間百分比無顯著差異外(P>0.05),橈足類和鹵蟲所占百分比較小,但2組間呈顯著差異(P<0.05)。試驗結果表明,魚-菜共生模式不但有效地抑制了藍藻的繁殖,促進可被羅非魚有效利用的硅藻種群數量,還提高了維持養殖水體穩態的原生動物的種群數量,進一步提高了養殖水體的生態平衡。
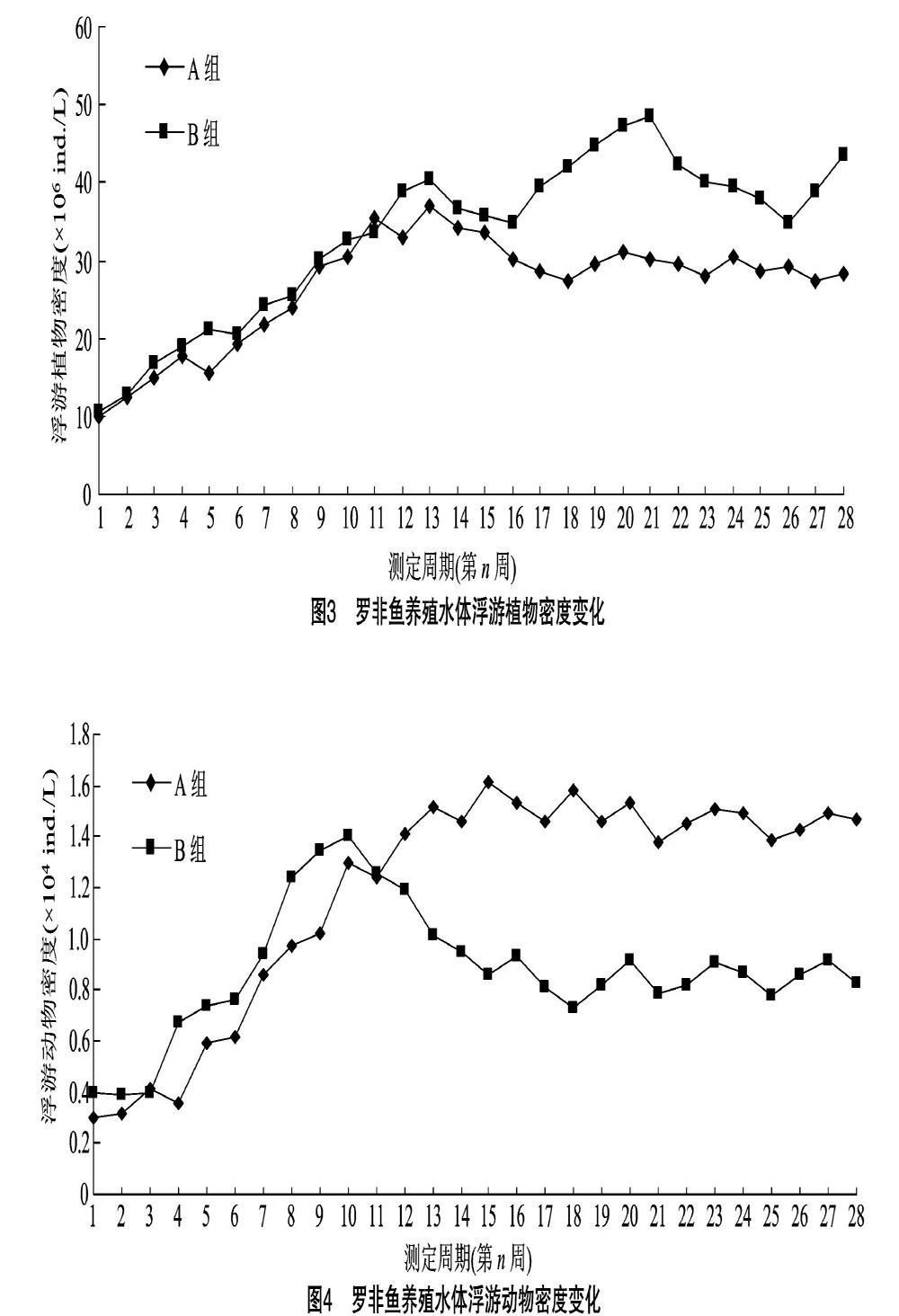
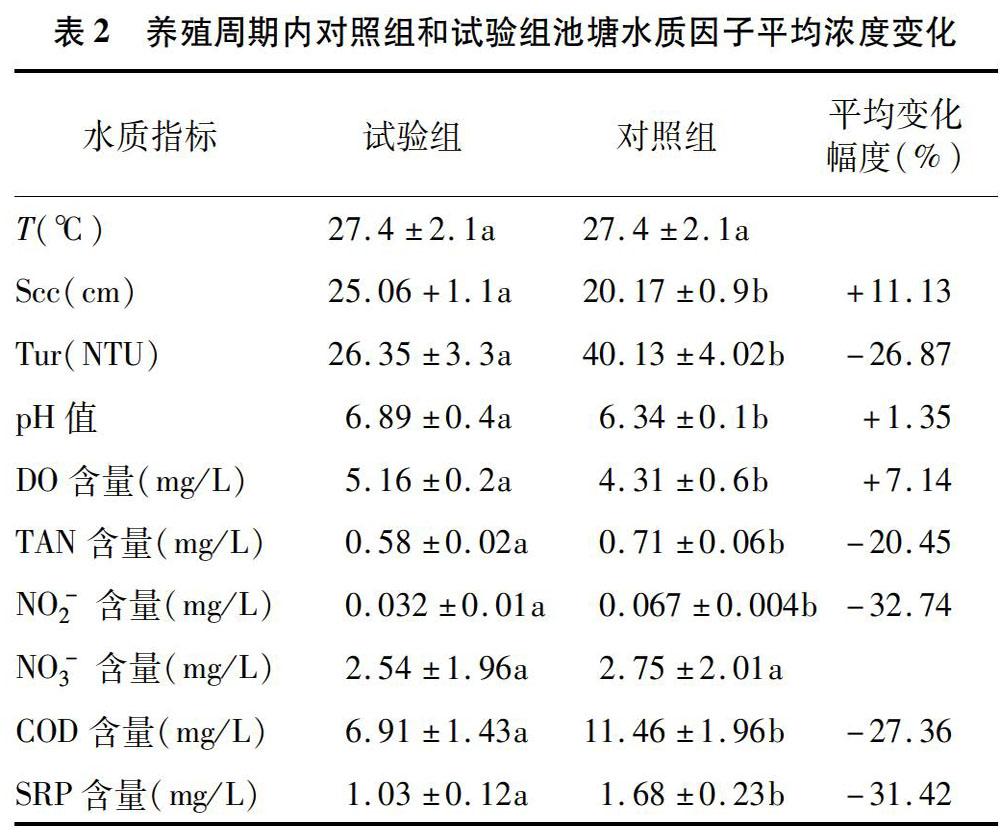
2.4 浮游生物密度與生物量變化
試驗周期內2組池塘養殖水體浮游植物密度變化見圖3。由圖3可見,在試驗前期,隨著氣溫升高和養殖飼料投喂量的增加,2組池塘養殖水體中浮游植物密度除在前5~6周時出現較小的下降變化外,整體增長較快并在第13周達到增長拐點。在試驗周期13~28周內,試驗組和對照組養殖水體浮游植物密度分別在27.3×106~31.2×106 ind./L 和34.8×106~48.6×106 ind./L之間波動,養殖水體的浮游植物密度均值分別為30.525×106 ind./L 和39.125×106 ind./L。試驗結果表明,魚-菜共生養殖模式對羅非魚養殖水體浮游植物密度有較好的穩定作用,進而促進形成穩態養殖水體。
試驗周期內2組池塘養殖水體浮游動物密度變化見圖4。由圖4可知,在試驗期內,隨著氣溫升高和養殖飼料投喂量的增加,2組池塘養殖水體中浮游動物密度除在前4周內無顯著變化外,整體呈線性增長并在第9周達到增長變化臨界點,這可能與原生動物的數量變化有關。在試驗周期10~28周內,試驗組和對照組養殖水體浮游動物密度呈現相反的變化波動,且兩者的變化分別在1.02×104~158×104 ind./L和0.73×104~1.4×104 ind./L 之間波動。該結果表明,魚-菜共生養殖模式可間接改變養殖水體浮游動物種群數量變化。
2.5 浮游生物多樣性的變化
試驗周期內2組養殖水體浮游生物多樣性指數見表4。由表4可知,試驗前期,2組池塘養殖水體浮游生物多樣性及均勻度指數變化不明顯,試驗中后期變化差異較大。整個試驗周期內,除浮游動物的Pielou 均勻度指數(J)無顯著差異外(P>0.05),試驗組浮游植物Shannon-Weaver多樣性指數(H)與 Pielou 均勻度指數(J)和浮游動物 Shannon-Weaver多樣性指數(H)均顯著高于對照組(P<005)。
3 討論
(1)魚-菜共生池塘養殖模式對羅非魚生長特性的影響。陳家長等研究表明,魚-菜共生池塘養殖模式對養殖池塘水質改善和經濟效益提高方面具有顯著促進作用[8,10],隨著研究的深入,魚-菜共生池塘養殖模式在養殖水體富營養化治理、水體健康評價和水質生物監測等方面具有非常廣闊的應用前景。該試驗數據顯示,與普通組相比,魚-菜共生試驗組羅非魚特定生長率、肥滿度、成活率和凈產量分別提高了18.90%、0.37%、13.30%和3070%,飼料系數降低了10.60%,同時凈增值 0.5萬元/1 000 m2[12-16]。本試驗數據顯示,魚-菜共生養殖技術不僅可提高羅非魚養殖成活率和特定生長率,還可顯著提高飼料利用率進而達到環保、增值效果,這與陳家長等[10,17]等對養殖池塘研究結果一致,不失為一種高效健康養殖模式。
(2)魚-菜共生養殖模式對羅非魚養殖水體水質的調控。大量研究表明,生物浮床或者有根植物能夠有效去除水體中總氮、BOD、SS、總磷、氨氮和重金屬等污染物[18-21]。本試驗魚菜共生-生物浮床研究結果顯示,在對照組與試驗組之間除養殖水體水溫和硝酸鹽平均濃度無顯著差異外(P>0.05),試驗組養殖水體pH值、透明度、溶解氧含量等水質因子均不同程度地顯著高于對照組(P<0.05),試驗組養殖水體濁度、氨氮、亞硝酸鹽、化學需氧量和溶解性正磷酸鹽等水質因子均顯著降低(P<0.05)。試驗研究結果表明,在相同飼養管理的前提下,對池塘養殖水體水質因子濃度的調節和生態系統的穩定具有較高的效率,該結果與曹煜成等[11,15]有關魚菜生物浮床對養殖水體調控的研究基本一致。
已有學者研究表明,因生物浮床栽培的水生植物生長在水面,影響水體氣液相接觸面積,進而降低養殖池塘水體空氣溶解量[22]。此外,植物的遮陽作用,阻礙水體藻類等沉水植物光合作用的同時,營呼吸作用的植物根系及其周圍營硝化作用的微生物仍會消耗氧氣,進一步導致養殖水體“氧源”減少[23]。也有部分學者認為,當生物浮床覆蓋面積小于池塘養殖水體面積的20%時,對池塘養殖水體溶解氧含量的變化有影響,但不顯著,覆蓋面積降低至7.5%時,養殖池塘水體的溶氧量無差異[10-11]。本試驗結果顯示,試驗養殖池塘中水體的溶解氧含量有所增加,與上述研究結果相悖,其可能原因是在低覆蓋率條件下,魚-菜共生系統改變了養殖池塘水體中微生物的種群和密度,促使水體生態系統形成微生態平衡,進而達到水體穩態。這種“綜合效應”進一步改善了養殖水體的營養鹽流通,降低了含氮化合物含量及溶解氧消耗,具體原理仍需進一步研究。
(3)魚-菜共生養殖模式對羅非魚養殖水體浮游動植物密度的影響。浮游生物的種類組成、群落結構和豐度變化,直接影響水體水質、系統內物質能量流和生物資源演替[24],池塘系統內浮游生物群落由多種原生動物和浮游植物種類構成[25],浮游生物群落種群能隨外界環境的改變而做出規律性的變化[26-30],形成復雜和多樣性的群落組合,呈現出不同的生物多樣性指數[25-27],進而促進養殖池塘系統內的能量流動、物質循環和信息傳遞[31-32]。所以,養殖池塘浮游生物的種類組成、密度及多樣性特征可以作為養殖水體健康狀況的指示[33-34]。
浮游生物多樣性指數通常用以反映養殖水體浮游生物群落結構組成,或富營養化對養殖水生生物的影響[27-29]。Shannon-Wiener 指數表示浮游生物群落組成,指數值越大預示著浮游生物群落結構越復雜,養殖池塘水體穩定性也越大[35]。均勻度是多樣性指數實際與理論的比值,描述水體中浮游生物種類、數量的均勻程度,多用于水體浮游生物多樣性評估[36]。優勢種種類和密度對水體群落結構的穩定性有重要作用,優勢種種類多且優勢度越小,則群落結構相對穩定[37]。本試驗結果表明,2組池塘中原生動物的密度均呈現出先上升后下降、最終趨于穩定的形態,試驗組池塘浮游動物多樣性顯著高于普通組,這與已有的研究結果基本一致。浮游動植物多樣性指數和均勻性指數均高于普通組,這與鄧金釵等羅非魚混養池塘研究中浮游動植物多樣性指數[38]有部分差異,可能是由于單養和混養主體生物的差異引起,具體原因仍需進一步研究。
魚-菜共生模式能夠在一定程度上影響池塘浮游生物物群落結構,豐富池塘浮游生物多樣性,改善養殖水質,促進養殖水體趨于穩定和健康,不失為一種生態、高效的池塘養殖模式。
參考文獻:
[1]徐 皓,劉興國,吳 凡. 池塘養殖系統模式構建主要技術與改造模式[J]. 中國水產,2009,9(8):7-9.
[2]Piedrahita R H. Reducing the potential environmental impact of tank aquaculture effluents through intensification and recirculation[J]. Aquaculture,2003,226(1):35-44.
[3]董雙林. 高效低碳——中國水產養殖業發展的必由之路[J]. 水產學報,2011,35(10):1595-1600.
[4]王 峰,雷霽霖,高淳仁,等. 國內外工廠化循環水養殖研究進展[J]. 中國水產科學,2013,20(5):1100-1111.
[5]王彥波,許梓榮. 微生物對水產養殖環境的生物修復作用[J]. 飼料研究,2004(12):42-43.
[6]Barak Y,Cytryn E,Gelfand I,et al. Phosphorus removal in a Marine prototype,recirculating aquaculture system[J]. Aquaculture,2003,220(1):313-326.
[7]Pedersen L F,Meinelt T,David L. Straus peracetic acid degradation in freshwater aquaculture systems and possible practical implications[J]. Aquacult Eng,2013,53:65-71.
[8]丁永良,張明華,張建華,等. 魚菜共生系統的研究[J]. 中國水產科學,1997,4(增刊1):71-76.
[9]常會慶,寇太記,喬鮮花,等. 幾種植物去除污染水體中養分效果研究[J]. 水土保持通報,2009(5):118-122.
[10]陳家長,孟順龍,胡庚東,等. 空心菜浮床栽培對集約化養殖魚塘水質的影響[J]. 生態與農村環境學報,2010,26(2):155-159.
[11]賀艷輝,張紅燕,龔贇翀,等. 我國羅非魚出口面臨的機遇與挑戰[J]. 中國漁業經濟,2009,27(3):85-89.
[12]賈 悅,李秀珍,唐瑩瑩,等. 不同采收方式對富養化河道浮床空心菜生物產出的影響[J]. 生態學雜志,2011,30(6):1091-1099.
[13]周小平,王建國,薛利紅,等. 浮床植物系統對富營養化水體中氮、磷凈化特征的初步研究[J]. 應用生態學報,2005,16(11):2199-2203.
[14]由文輝,劉淑媛,錢曉燕. 水生經濟植物凈化受污染水體研究[J]. 華東師范大學學報(自然科學版),2000,3(1):99-102.
[15]楊曉玲,郭金耀. 水蕹菜對富營養化養殖水的凈化作用研究[J]. 作物雜志,2012(1):49-52.
[16]陳玉紅,劉忠良. 豬場厭氧廢水用于空心菜水培試驗研究[J]. 農業環境與發展,2011(3):87-90.
[17]谷孝鴻. 不同養殖類型池塘浮游生物群落結構的初步分析[J]. 湖泊科學,1994,6(3):276-282.
[18]周真明,陳燦瑜,葉 青,等. 浮床植物系統對富營養化水體的凈化效果[J]. 華僑大學學報(自然科學版),2010(5):576-579.
[19]呂錫武,宋海亮. 水培蔬菜法對富營養化水體中氮磷的去除特性研究[J]. 江蘇環境科技,2004,6(2):1-3.
[20]操家順,李欲如,陳 娟. 水蕹菜對重污染河道凈化及克藻功能[J]. 水資源保護,2006,22(2):36-38,41.
[21]Yin Z,Chen J,Zeng L,et al. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight[J]. Molecular Plant-microbe Interactions:MPMI,2000,13(8):869-876.
[22]任照陽,鄧春光. 生態浮床技術應用研究進展[J]. 農業環境科學學報,2007(增刊1):261-263.
[23]馬井泉,周懷東,董哲仁. 我國應用生態技術修復富營養化湖泊的研究進展[J]. 中國水利水電科學研究院學報,2005,3(3):209-215.
[24]趙 文. 刺參池塘養殖生態學及健康養殖理論[M]. 北京:科學出版社,2009.
[25]李丹丹. 不同模式羅非魚養殖池塘浮游生物群落結構動態變化[D]. 南京:南京農業大學,2015:1-7.
[26]李志斐,王廣軍,陳鵬飛,等. 生物浮床技術在水產養殖中的應用概況[J]. 廣東農業科學,2013,40(3):106-108,114.
[27]李勝男,周 建,魏利軍,等. 淡水超微型浮游植物多樣性及其研究方法[J]. 生態學雜志,2015,34(4):1174-1182.
[28]Rakesh R. Yeredla,Xu H F.incorporating strong polarity minerals of tourmaline with semiconductor Titania to improve the photosplitting of water[J]. The Journal of Physical Chem-istry C,2008,112(2):532-539.
[29]Xu H L,Cao Q,Wang X Y,et al. Properties and chemical oxidation polymerization of polyaniline/neutral red/TiO2 composite electrodes[J]. Materials Science and Engineering:B,2010,171(1):104-108.
[30]Yuan Y,Ahmed J,Kim S H. Polyaniline/Carbon black com-posite-supported Iron phthalocyanine as an oxygen reduction catalyst for microbial fuel cells[J]. Journal of Power Sources,2011,196(3):1103-1106.
[31]李 今,呂 田,華江環. 人工浮床水培空心菜生長特性及其在養殖廢水凈化中的應用[J]. 湖南師范大學自然科學學報,2014,37(2):22-27.
[32]胡綿好. 水生經濟植物浮床技術改善富營養化水體水質的研究[D]. 上海:上海交通大學,2008.
[33]Tavernini S,Mura G,Rossetti G. Factors influencing the seasonal phenology and composition of zooplankton communities in mountain temporary pools[J]. International Review of Hydrobiology,2005,90(4):358-375.
[34]Echaniz S A,Vignatti A M,de Paggi S J,et al. Zooplankton seasonal abundance of South American saline shallow lakes[J]. International Review of Hydrobiology,2006,91(1):86-100.
[35]Lai B,Wang P,Li H,et al. Calcined polyaniline-iron composite as a high efficient cathodic catalyst in microbial fuel cells[J]. Bioresource Technology,2013,131(0):321-324.
[36]Rao S M,Xing Y C. Simulation of nanostructured electrodes for polymer electrolyte membrane fuel cells[J]. Journal of Power Sources,2008,185(2):1094-1100.
[37]Logan B E,Murano C,Scott K,et al. Electricity generation from cysteine in a microbial fuel cell[J]. Water Research,2005,39(5):942-952.
[38]鄧金釵,熊邦喜,張小敏,等. 池塘主養草魚三種養殖模式浮游植物群落結構的比較[J]. 淡水漁業,2012,42(5):63-67. 周 聃,崔雁娜,周冬仁,等. 不同養殖模式下加州鱸魚肉質比較分析[J]. 江蘇農業科學,2021,49(6):146-149.
