蔗糖、硼酸、Ca2+對大白杜鵑花粉萌發的影響
2021-05-06 13:06:53王波周蘭英夏華梅
江蘇農業科學 2021年6期
王波 周蘭英 夏華梅
摘要:探究大白杜鵑花粉萌發所需的營養液濃度組合,為大白杜鵑的資源保存及雜交育種提供科學依據。以大白杜鵑新鮮花粉為試驗材料,利用單因子試驗比較固體培養基中不同濃度的蔗糖、硼酸、Ca2+對其萌發的影響,在此基礎上進行正交試驗,探討大白杜鵑花粉萌發的最佳蔗糖、硼酸、Ca2+濃度組合。結果表明,在一定質量濃度范圍內,蔗糖、硼酸、Ca2+對大白杜鵑花粉萌發均起到促進作用,但是超過一定質量濃度時則起抑制作用,最適濃度分別為10%、0005%、0.002%。蔗糖、硼酸和Ca2+這3個因素的綜合作用對大白杜鵑花粉萌發有顯著影響,大白杜鵑花粉的最適培養基為10%蔗糖+0.005%硼酸+0.002%Ca2+。
關鍵詞:大白杜鵑;花粉;萌發;培養基
大白杜鵑(Rhododendron decorum Franch.),別稱大白花杜鵑,是杜鵑花科(Ericaceae)杜鵑花屬(Rhododendron L.)木本植物,在我國主要分布在四川、云南、貴州、西藏等地,在四川省會理縣生長于龍肘山海拔1 900~2 100 m處,種群成片分布。大白杜鵑花簇多花,形如繡球,色澤艷麗;是杜鵑花中少有的香花品種,具有很高的觀賞價值;分布廣泛、海拔較低,是香花育種的理想親本。張敬麗等研究發現,馬纓花(Rhododendron delavayi Franch.)與大白花杜鵑進行雜交后,雜交種能夠結出可育的種子[1]。此外,大白杜鵑還可供食用和藥用[2-3],開發利用前景廣闊。但是,人為干預使其生境遭到嚴重破壞,資源日趨受到威脅,環境干旱使野生杜鵑結實率降低[4],大白杜鵑物種資源的保存迫在眉睫。
目前,國內外對大白杜鵑的研究主要集中在生態分布、染色質組成[5]、營養成分[2]和組織培養[6]等方面,有關大白杜鵑花粉的儲存及其生活力、萌發條件的研究未見報道。目前已有少數其他杜鵑種花粉的研究,如徐芬芬等對毛杜鵑(Rhododendron pulchrum)花粉離體萌發特性的研究發現,毛杜鵑花粉萌發的最適硼酸質量濃度為10 mg/L,培養7 h后花粉萌發情況穩定[7]。李玉萍等以滿山紅、映山紅、馬銀花、錦繡杜鵑的新鮮花粉為試驗材料,測定不同液體培養基和儲藏方法下的花粉萌發力,發現杜鵑花花粉的適宜培養基配方為100 g/L蔗糖+20 mg/L 硼酸+20 mg/L CaCl2[8]。目前,關于其他植物花粉萌發的研究非常多,不同植物花粉萌發要求的最適培養基組合和濃度不同,但適量的蔗糖、硼酸、CaCl2會提高多數植物花粉的萌發率。目前,關于大白杜鵑花粉萌發的研究未見報道,本試驗用蔗糖、硼酸及CaCl2 3個因素組成的固體培養基研究大白杜鵑花粉的萌發力,旨在找出適合大白杜鵑花粉萌發的營養液濃度組合,為大白杜鵑的資源保存及雜交育種提供科學依據。
1 材料與方法
1.1 試驗材料
本研究所用材料于2013年5月9日采自四川省會理縣龍肘山海拔1 900~2 100 m處。選擇20株盛花期大白杜鵑植株上即將開放的100朵花,用濕毛巾包裹后放入裝有冰塊的潔凈紙盒內冷藏并迅速帶回實驗室,任其花藥自然開裂,收集花粉,并將花粉充分混勻,避免個體植株花藥不同引起的花粉生活力差異。將收集的花粉裝入花粉瓶中,于 4 ℃ 低溫冷藏。
1.2 培養基配方和制作方法
單因素試驗中各因素濃度水平[9-10]如表1所示,以同樣濃度的空白培養基作為對照。在進行單因素試驗后,總結分析各因素濃度對花粉萌發的影響,并在此基礎上進行L16(43)正交試驗。按濃度配方配制100 mL溶液,加入搪瓷碗中,煮沸后向其中加入1 g瓊脂粉,攪拌至充分溶解,冷卻后作為花粉接種用固體培養基。
1.3 花粉培養觀測方法
在雙孔凹型玻片孔內滴1滴清水以保持濕度,在蓋玻片上滴入適量培養基,將花粉均勻播種在培養基上,再將有培養基的一面朝下蓋在玻片孔上,每個培養基設5個重復。將玻片放入25 ℃人工培養箱中,在黑暗條件下培養。每隔2 h觀測1次,發現花粉萌發后,每隔1 h觀測1次,最終從2次測定萌發率基本相同開始測定萌發率。大白杜鵑花粉為四合體,單粒花粉為圓球形[11-12],在統計花粉萌發數時,只要有1個花粉萌發即視為萌發,以花粉管長度超過花粉直徑作為判斷萌發的標準。每個玻片觀察5個視野,每個視野觀察的花粉數不少于30粒,記錄花粉的萌發數量,花粉萌發率的計算公式如下:
花粉萌發率=已萌發的花粉粒數/測定的花粉粒總數×100%。
1.4 數據統計分析
用Excel軟件進行數據整理,并用SPSS軟件對正交試驗數據進行單因變量多因素方差分析,若差異顯著,則在方差分析的基礎上用新復極差法(SSR)進行因素水平間的多重比較。
2 結果與分析
2.1 大白杜鵑花粉萌發的基本情況
將大白杜鵑花粉于25 ℃黑暗條件下處理3 h后,四合體吸水膨大,極面觀為三裂球形;6 h后花粉管從寬大的萌發溝內長出,并逐漸增長;10 h后萌發基本趨于穩定,花粉管的長度達到最長。在按試驗對照條件處理12 h后,花粉的萌發效果見圖1。
2.2 單因素試驗中不同濃度蔗糖、硼酸、CaCl2對大白杜鵑花粉萌發的影響
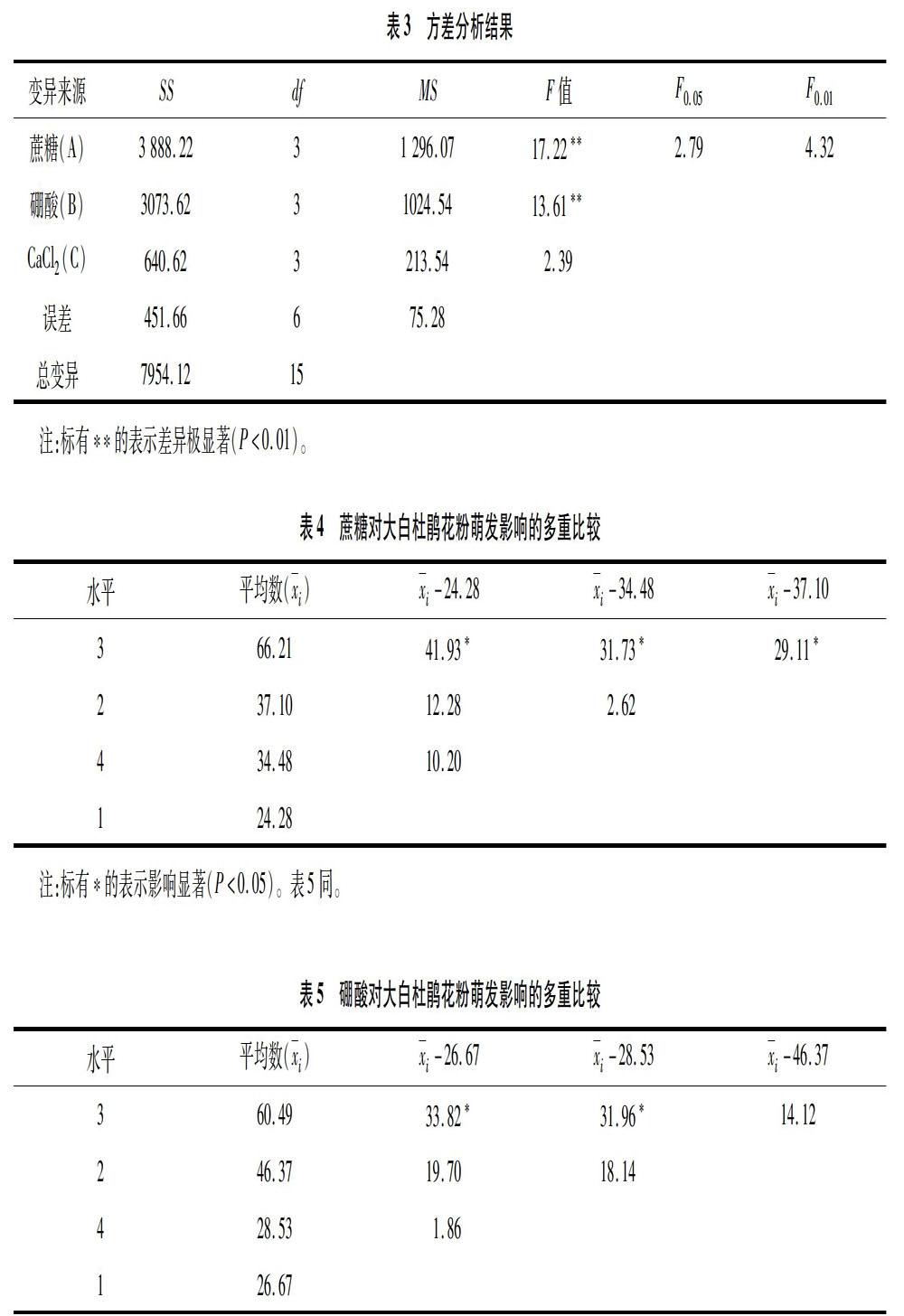
2.2.1 蔗糖對大白杜鵑花粉萌發的影響 由圖2可知,不同濃度蔗糖對大白杜鵑花粉萌發有不同程度的影響。當蔗糖濃度為0、2%、5%、10%時,花粉萌發率隨著蔗糖濃度的升高而提高,分別為688%、20.77%、54.91%、74.23%,最大萌發率為74.23%;但當蔗糖濃度超過10%以后,花粉萌發率隨著蔗糖濃度的增加而下降。綜上,大白杜鵑花粉萌發的適宜蔗糖濃度為10%。
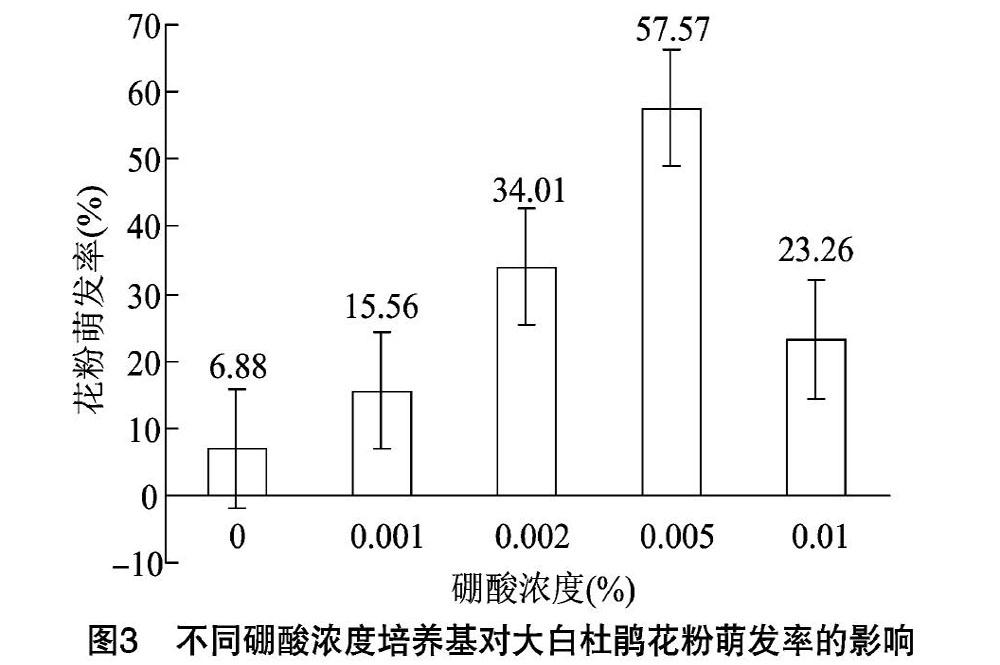
2.2.2 硼酸對大白杜鵑花粉萌發的影響 由圖3可知,不同濃度硼酸對大白杜鵑花粉萌發有不同程度的影響。當硼酸濃度為0~0.005%時,花粉萌發率隨著硼酸濃度的升高而提高,最大萌發率為5757%;但當硼酸濃度超過0.005%以后,花粉萌發率隨著硼酸濃度的增加而下降。綜上,大白杜鵑花粉萌發的適宜硼酸濃度為0.005%。
2.2.3 CaCl2對大白杜鵑花粉萌發的影響 由圖4可知,不同濃度CaCl2對大白杜鵑花粉萌發有不同程度的影響。當CaCl2濃度為0~0.002%時,花粉萌發率隨著CaCl2濃度的升高而提高,最大萌發率為39.47%; 但當CaCl2濃度超過0.002%以后, 花粉萌發率隨著CaCl2濃度的增加而下降。綜上,大白杜鵑花粉萌發的適宜CaCl2濃度為0.002%,并且高濃度的CaCl2會明顯抑制大白杜鵑的花粉萌發。
2.3 正交試驗結果分析
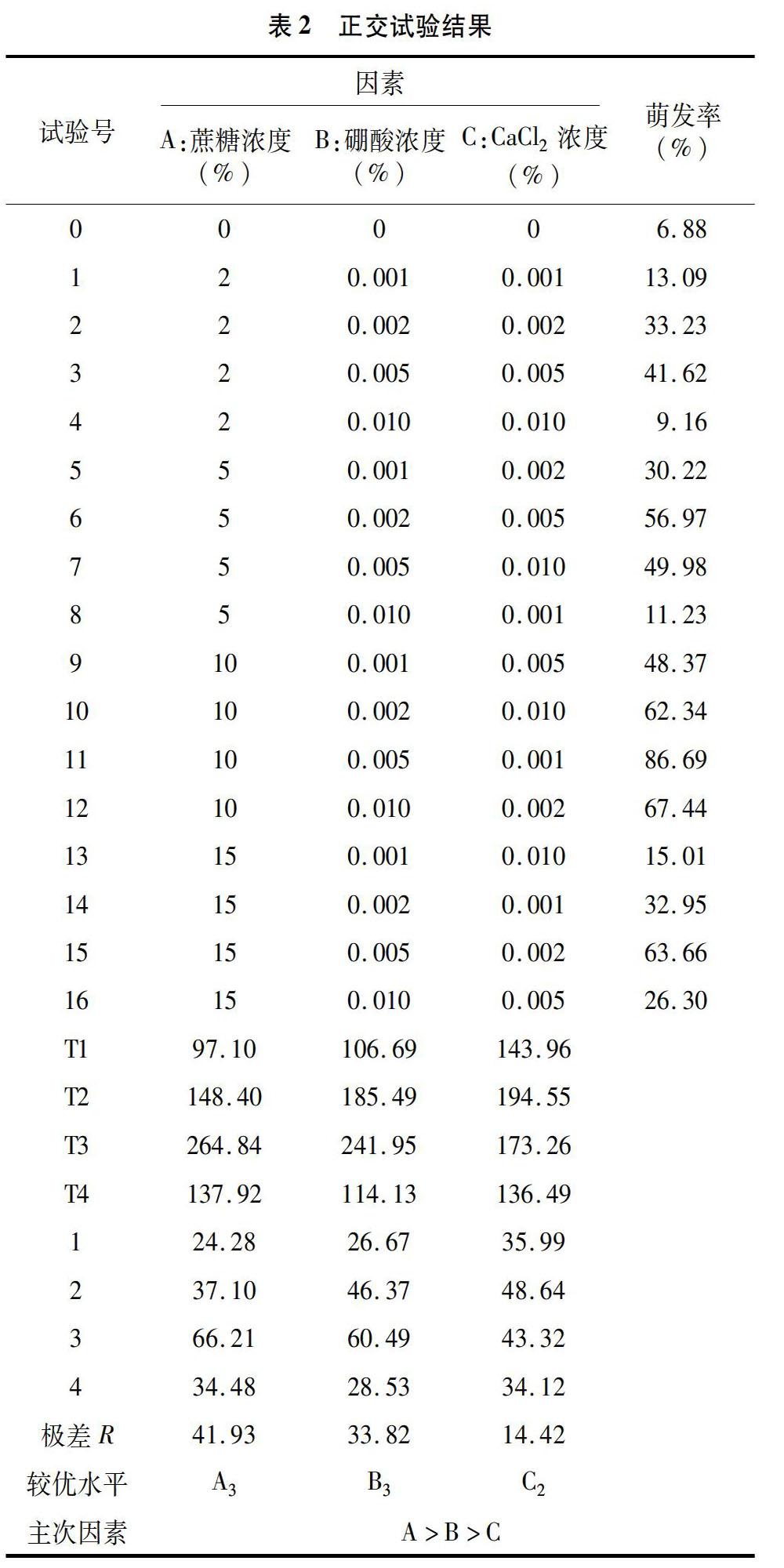
單因素試驗結果表明,對照條件下大白杜鵑花粉有少量萌發,表明外源因素不是萌發的必要因素。如表2所示,大白杜鵑花粉在10%蔗糖+0005%硼酸+0.001%CaCl2培養基配方下的萌發率最高,達到86.69%。極差分析結果顯示,對花粉萌發率影響的因素排序為蔗糖、硼酸、CaCl2,極差依次為167.74、135.26、58.06,相應的較優水平分別為A3(10%)、B3(0.005%)、C2(0.002%),由較優水平組合而成的最佳培養基配方為10%蔗糖+0005%硼酸+0.002%CaCl2,與單因素試驗得出的正交試驗結果最高的萌發率培養基配方一致。正交試驗結果表明,蔗糖、硼酸和CaCl2在一定濃度范圍內共同作用時對大白杜鵑花粉的萌發有一定促進作用;大白杜鵑花粉萌發的最適培養基配方為10%蔗糖+0.005%硼酸+0.002%CaCl2。
3 結論與討論
很多研究發現,植物花粉離體萌發的影響因素很多,內因包括供體植株基因型、發育時期、花粉生活力等,外因如培養基固液形態、瓊脂濃度、培養溫度和濕度、光照條件、培養基營養種類和濃度等[13],其中蔗糖、硼酸及一些礦質元素如外源Ca2+等的參與是比較重要的,是誘導離體花粉萌發的基本條件。在本試驗中,對照處理的花粉也具有一定的萌發率,由此或可大膽推測,花粉可能本身就含有少量蔗糖、硼酸及Ca2+。目前已知,細胞壁是植物細胞最大的鈣庫,用X射線分析鈣在花粉粒中的分布結果也顯示,多數植物花粉壁上含有少量鈣[14]。
一般認為,蔗糖在花粉萌發過程中的作用有2個,一是為花粉萌發和花粉管的生長提供能源和碳源,二是調節培養環境滲透壓[15-16]。試驗中一定濃度的蔗糖促進大白杜鵑花粉萌發,最適宜的蔗糖濃度為10%,當蔗糖濃度過高時,花粉萌發率開始下降,這與徐芬芬等的研究結果[17]相似。這有可能是因為適當濃度的蔗糖能維持花粉與培養基之間的滲透平衡,而過高的蔗糖濃度則會導致花粉管破裂產生質壁分離現象,從而造成原生質體脫水,抑制花粉萌發生長,降低萌發率[18-19]。
目前許多研究發現,硼作為植物必需的微量元素,在花粉萌發和花粉管伸長中的主要作用是增加糖的吸收、運轉和代謝,以增強花粉管壁的可塑性[20]。在離體條件下,補充一定量的外源硼,可以大大提高其花粉萌發率。在本試驗中,提高硼酸濃度顯著提高了花粉萌發率,當硼酸濃度為0.005%時,大白杜鵑花粉的萌發率最高。
鈣離子在生物體的電化學震蕩理論中被稱為細胞中調節的“第二信使”,鈣離子水平受到環境中鈣含量和植物自身鈣受體的影響,從而引起一系列生長發育過程。在花粉的離體培養過程中,Ca2+可以通過多種途徑影響花粉萌發和花粉管的生長,其中涉及細胞骨架的組裝、分泌小泡的運輸和融合、花粉管頂端的脈沖式生長、花粉管生長方向調控等各個方面[21-24]。許多研究發現,外源鈣對花粉萌發和花粉管生長的作用取決于細胞內游離鈣的含量,即在細胞內游離鈣充足或已充分吸收外源鈣時,外源鈣對花粉萌發和花粉管生長就可能起抑制作用,反之則起促進作用。本試驗中單因素Ca2+濃度明顯影響大白杜鵑花粉萌發,表明一定程度的外源鈣能促進大白杜鵑花粉萌發和花粉管生長,當Ca2+濃度為0.002%時,花粉萌發率最高,而當Ca2+濃度高于0.005%時,花粉萌發則明顯受到抑制,這與周瑞金等的研究結果[25]一致。
通過正交試驗,得到大白杜鵑花粉萌發的最適培養基配方是10%蔗糖+0.005%硼酸+0.002% CaCl2,在此條件下花粉萌發率最高,且遠超過單因素試驗中的最高萌發率。另外,當鈣離子為高水平而硼酸為低水平,或鈣離子為低水平而硼酸為高水平時,兩者表現出負交互作用。原因可能是單因素之間的交互作用促進了花粉萌發,如硼酸可以與蔗糖形成絡合物,從而促進果膠物質的合成、糖的吸收,進而更大程度地促進花粉萌發[15]。此外,硼還可能通過影響關鍵酶活性與Ca2+濃度梯度相關聯,影響其花粉管細胞壁的構建與生長[26]。試驗結果顯示,蔗糖對花粉萌發影響最大,硼酸次之,Ca2+對花粉萌發有促進作用,但影響比硼酸小。鈣離子對花粉萌發的影響較硼酸小的結論得到了其他研究者的支持,一般認為花粉細胞內鈣離子較多而硼素較少,因而萌發時胞外硼素的影響較胞外鈣離子的影響高[27]。
通過花期噴施具有促進花粉萌發和花粉管生長的物質的方法來提高結實率和促進種子發育、膨大是生產上的一項重要措施。本研究結果表明,大白杜鵑花粉萌發的最適培養基組合10%蔗糖+0005%硼酸+0.002%CaCl2可為大白杜鵑雜交育種提供技術上的參考。在花期噴灑蔗糖、硼酸、CaCl2或者其三者組合有助于大白杜鵑結實,而在授粉過程中噴灑含有蔗糖、硼酸、CaCl2的花粉營養液的方法是輔助授粉的較好手段。但在實際生產中,由于營養液對花粉萌發的作用可能還與樹齡和溫度等其他因素有關,因此營養液的適宜濃度應根據環境條件經試驗而定。
參考文獻:
[1]張敬麗,張長芹,吳之坤,等. 探討種間傳粉在杜鵑花屬自然雜交育種形成中的應用[J]. 生物多樣性,2007,15(6):658-665.
[2]唐丹林,張光凡,陳孝泉. 大白杜鵑尖葉杜娟的營養成分的研究[J]. 四川大學學報,1992,29(2):316-318.
[3]中國科學院《中國植物志》編委會. 中國植物志:第五十七卷 第二分冊[M]. 北京:科學出版社,1994.
[4]黃承玲,陳訓高,貴 龍. 3種高山杜鵑對持續干旱的生理響應及抗旱性評價[J]. 林業科學,2011,47(6):48-55.
[5]張洪淵,劉克武,楊守忠,等. 大白杜鵑染色質組成成分的研究[J]. 廣西植物,1990,10(1):49-53.
[6]羅 彭,莊 平,白 潔. 大白杜鵑、美容杜鵑和喇叭杜鵑的組織培養[J]. 植物生理學通訊,2007,43(2):326.
[7]徐芬芬,崔 燕,謝洪云,等. 毛杜鵑花粉離體培養萌發特性研究[J]. 黑龍江農業科學,2012(1):26-27.
[8]李玉萍,王燕青,羅鳳霞,等. 測定杜鵑花花粉萌發力的液體培養基和儲藏方法研究[J]. 金陵科技學院學報,2012,28(3):56-60.
[9]張超儀,耿興敏. 六種杜鵑花屬植物花粉生活力測定方法的比較研究[J]. 植物科學學報,2010,30(1):92-99.
[10]韓金多,謝洪云,程新平,等. 鹽脅迫對毛杜鵑花粉萌發的影響[J]. 湖南農業科技,2011(21):106-107.
[11]周蘭英,王永清,張 麗. 26種杜鵑屬植物花粉形態及分類學研究[J]. 林業科學,2008,44(2):55-64.
[12]高連明,張長芹,韋仲新,等. 杜鵑花花族花粉形態及其系統學意義[J]. 云南植物研究,2002,24(4):471-482.
[13]劉雪蓮,李慶玲,秦佳梅,等. 不同培養基對紫丁香花粉離體萌發的影響[J]. 東北林業大學學報,2010,38(7):61-63.
[14]龔 明,曹宗龔. 鈣和鈣調素對花粉萌發和花粉管生長的調控[J]. 植物生理學通訊,1999,31(5):321-328.
[15]杜玉虎,綦 影,蔣錦標,等. 蔗糖、鈣和硼對榆葉梅花粉離體萌發及花粉管生長的影響[J]. 北方園藝,2008(8):106-109.
[16]杜玉虎,張琳琳,蔣錦標,等. 蔗糖和礦質營養對榛子花粉離體萌發和花粉管生長的影響[J]. 遼寧農業職業技術學院學報,2008,10(2):1-3.
[17]滕長才,張永成. 硼酸和蔗糖對馬鈴薯花粉離體萌發的影響[J]. 種子,2009,28(2):15-16,20.
[18]年玉欣,羅鳳霞,張 穎,等. 測定百合花粉生命力的液體培養基研究[J]. 園藝學報,2005,32(5):922-925.
[19]朱展望,張改生,牛 娜. 小麥花粉的離體萌發研究[J]. 麥類作物學報,2007,27(1):12-15.
[20]楊小冬,孫素琴,李一勤. 硼缺乏導致花粉管細胞壁多糖分布的改變[J]. 植物學報,1999,41(11):1169-1176.
[21]Geitmann A,Cresti M. Ca2+ channels control the rapid expansions in pulsating growth of Petunia hybrida pollen tubes[J]. Journal of Plant Physiology,1998,152(4/5):439-447.
[22]Malho R,Read N D,Pais M S,et al. Role of cytosolic free calcium in the reorientat ion of pollen tube growth[J]. Plant Journal,1994,5(3):331-341.
[23]盛仙永,胡正海. Ca2+、pH在花粉及萌發花粉管生長中的作用研究進展[J]. 西北植物學報,2005,25(1):194-199.
[24]周建濤,姜雪婷,李 慧,等. 鈣調素及其抗血清對梨花粉萌發和花粉管生長的影響[J]. 江蘇農業學報,2016,32(2):437-441.
[25]周瑞金,李桂榮,張傳來. 培養基成分對梨花粉萌發和花粉管生長的影響[J]. 河北林果研究,2009,24(4):420-422.
[26]田翠婷,呂洪飛,王 鋒,等. 培養基組分對青杄離體花粉萌發和花粉管生長的影響[J]. 北京林業大學學報,2007,29(1):47-52.
[27]張紹鈴,陳迪新,康 瑯,等. 培養基組分及pH值對梨花粉萌發和花粉管生長的影響[J]. 西北植物學報,2005(2):225-230.李大命,唐晟凱,劉燕山,等. 滆湖鰱、鳙種群生長特征及起捕規格[J]. 江蘇農業科學,2021,49(6):134-139.
