2019年廣州市甲型H1N1流感病毒全基因組序列的遺傳特征分析
2021-05-06 02:02:32陳藝韻陸劍云張周斌李魁彪
中國人獸共患病學(xué)報(bào) 2021年4期
曹 藍(lán),吳 迪,陳藝韻,曾 慶,夏 丹,陸劍云,狄 飚,張周斌,李魁彪
長期以來,季節(jié)性流感威脅著人類健康,每年造成嚴(yán)重的人群發(fā)病和死亡。甲型H1N1流感病毒自2009年首次出現(xiàn)以來,由于不斷快速的發(fā)生基因變異,已代替了原季節(jié)性H1N1流感而持續(xù)流行。Mi L等[1]在對1977年以來全球H1N1流感病毒抗原進(jìn)化模型的研究中發(fā)現(xiàn),在8個(gè)主要進(jìn)化模式中有6個(gè)進(jìn)化模式起源于亞洲,進(jìn)一步發(fā)現(xiàn),我國南方地區(qū)H1N1流感病毒的進(jìn)化要早于北方地區(qū),提示南方地區(qū)H1N1流感病毒在我國H1N1流感傳播進(jìn)化過程起到重要作用。而廣州市地處華南地區(qū),是我國重要的經(jīng)濟(jì)發(fā)展中心,也是包括H1N1流感在內(nèi)多種傳染病流行的重要地區(qū)。早期研究發(fā)現(xiàn),廣州市流感流行呈現(xiàn)雙峰的流行特點(diǎn),甲型H1N1流感和季節(jié)性H3N2流感交替出現(xiàn)[2],同時(shí)甲型H1N1流感病毒也是近期廣州市流感疫情的主要病原[3]。既往的研究中重點(diǎn)對HA和NA基因進(jìn)行了變異分析,而對甲型H1N1基因組全序列的分子進(jìn)化研究相對較少,因此本研究選取7株2019年廣州市甲型H1N1流感流行株進(jìn)行了全基因組測序,為全面掌握近期廣州市甲型H1N1流感病毒的分子流行病學(xué)特點(diǎn)提供研究數(shù)據(jù)。
1 材料與方法
1.1毒株來源 收集2019年廣州市流感監(jiān)測哨點(diǎn)醫(yī)院流感樣病例和社區(qū)流感暴發(fā)疫情病例的呼吸道標(biāo)本,通過熒光定量RT-PCR方法進(jìn)行甲型H1N1、甲型H3N2和乙型流感病毒核酸檢測(試劑盒購于江蘇碩世生物科技公司),并將流感陽性標(biāo)本接種MDCK細(xì)胞系進(jìn)行病毒分離,培養(yǎng)產(chǎn)物經(jīng)過血凝實(shí)驗(yàn)-血凝抑制試驗(yàn)(HA-HI)對流感病毒培養(yǎng)物進(jìn)行鑒定(血球?yàn)樽灾齐嗍笱逵芍袊膊☆A(yù)防控制中心提供)。根據(jù)分離時(shí)間的不同,本研究中隨機(jī)選取7株甲型H1N1流感病毒進(jìn)行全基因組測序。
1.2全基因組序列測定 參考Deng YM等[4]關(guān)于流感病毒全基因組測序的方法,并參考近期甲型H1N1流感病毒各基因全長序列,應(yīng)用Oligo 6軟件設(shè)計(jì)甲型H1N1流感病毒全序列各基因片段全長測序引物(引物由廣州天一輝遠(yuǎn)生物公司合成),見表1。通過RT-PCR方法擴(kuò)增HA、NA、PB2、PB1、PA、NP、M和NS全長序列,陽性鑒定產(chǎn)物送至廣州天一輝遠(yuǎn)生物公司,通過ABI 3730進(jìn)行病毒一代測序。

表1 甲型H1N1流感病毒各基因片段擴(kuò)增引物Tab.1 Amplification primers for gene fragments of the avian influenza A(H1N1)pdm09 virus
1.3分子特征分析 通過DNA Star7.1軟件拼接全序列各基因片段,同時(shí)對各基因進(jìn)行同源性分析。采用MEGA 6.0軟件,以基因ORF為基本單元,對不同進(jìn)化分支變異氨基酸進(jìn)行分析。
1.4遺傳進(jìn)化分析 以GISAID數(shù)據(jù)庫中歷年WHO推薦的甲型H1N1流感病毒疫苗株各基因片段作為參比序列(A/Guangdong-Maonan/1536/2019 (H1N1)、A/Hawaii/70/2019(H1N1)、A/ Brisbane/02/2018 (H1N1)、A/Michigan/45/2015 (H1N1)、A/California/7/2009 (H1N1)、A /Christchurch/16/2010 (H1N1)、A/Brisbane/59/2007 (H1N1)、A/Solomon/Islands/3/2006 (H1N1)、A/New/Caledonia/20/1999 (H1N1)等)。使用MEGA 軟件繪制各基因遺傳進(jìn)化樹,參數(shù)設(shè)置為:Neighbor-joining法(參數(shù)設(shè)置為1 000 replications)及Maximum composite likelihood model核苷酸替代模型。
2 結(jié) 果
2.1總體情況 2019年累計(jì)從10 541份監(jiān)測標(biāo)本中檢測到甲型H1N1流感陽性標(biāo)本703份,其中分離甲型H1N1流感毒株70株(均感染MDCK細(xì)胞經(jīng)一代培養(yǎng))。本研究中7株病毒中,5株分離自流感監(jiān)測病例,2株分離自流感暴發(fā)疫情病例。2株分離于2019年1月,2株分離于2019年3月,2株分離于2019年6月,1株分離于2019年12月。
2.2全基因組序列測定 全序列各基因擴(kuò)增產(chǎn)物經(jīng)毛細(xì)管電泳鑒定,經(jīng)過序列拼接,獲得全基因各片段開放閱讀框序列(HA基因:1 701 bp、NA基因:1 410 bp、PB2基因:2 280 bp、PB1基因:2 274 bp、PA基因:2 151 bp、NP基因:1 497 bp、M1基因:759 bp、NS1基因:693 bp)。7株病毒全基因序列中,核苷酸同源性最高為PB2基因(98.8%~99.9%),最低為NA基因(97.3%~99.2%),氨基酸同源性最高為PB2蛋白(99.3%~100%),最低為NA蛋白(96.4%~99.4%),全基因組同源性見表2。
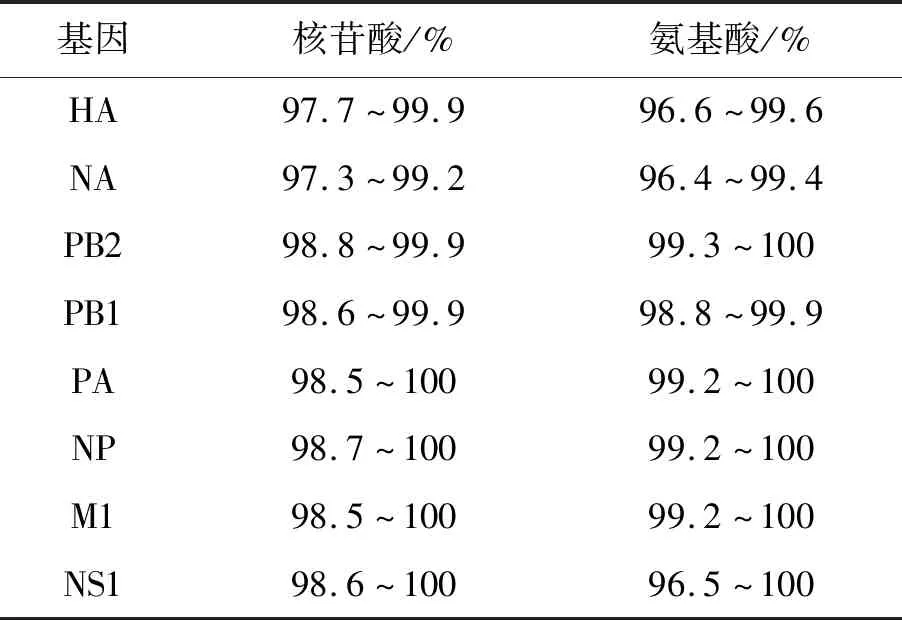
表2 2019年廣州市H1N1流感分離株全序列基因的同源性分析Tab.2 Sequence similarity analysis of whole genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019
2.3遺傳進(jìn)化分析 總體上,不同月份的H1N1流感分離株按時(shí)間分布,分別于A/Michigan/45/2015 (H1N1)、A/Brisbane/02/2018 (H1N1)、A/Guangdong-Maonan/1536/2019 (H1N1)、A/Hawaii/70/2019(H1N1)疫苗株聚為一簇,特別是2019年6月份以后的H1N1分離株與世界衛(wèi)生組織2020-2021年推薦使用的疫苗推薦株A/Guangdong-Maonan/1536/2019 (H1N1)親緣關(guān)系最近。分離于哨點(diǎn)醫(yī)院流感樣癥狀監(jiān)測病例的毒株與社區(qū)流感暴發(fā)疫情的毒株歸屬于同一進(jìn)化分支。全基因組遺傳進(jìn)化樹顯示,未發(fā)現(xiàn)不同基因來源的基因重配現(xiàn)象。
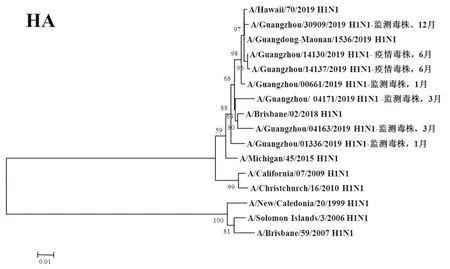
圖1 2019年廣州市甲型H1N1流感分離株HA基因遺傳進(jìn)化樹Fig.1 Genetic evolution analysis of HA genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019
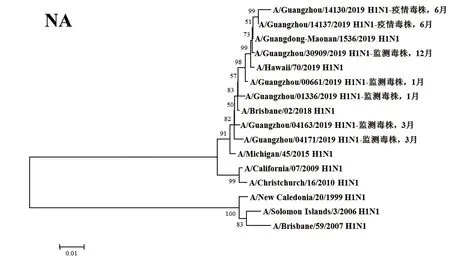
圖2 2019年廣州市甲型H1N1流感分離株NA基因遺傳進(jìn)化樹Fig.2 Genetic evolution analysis of NA genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019
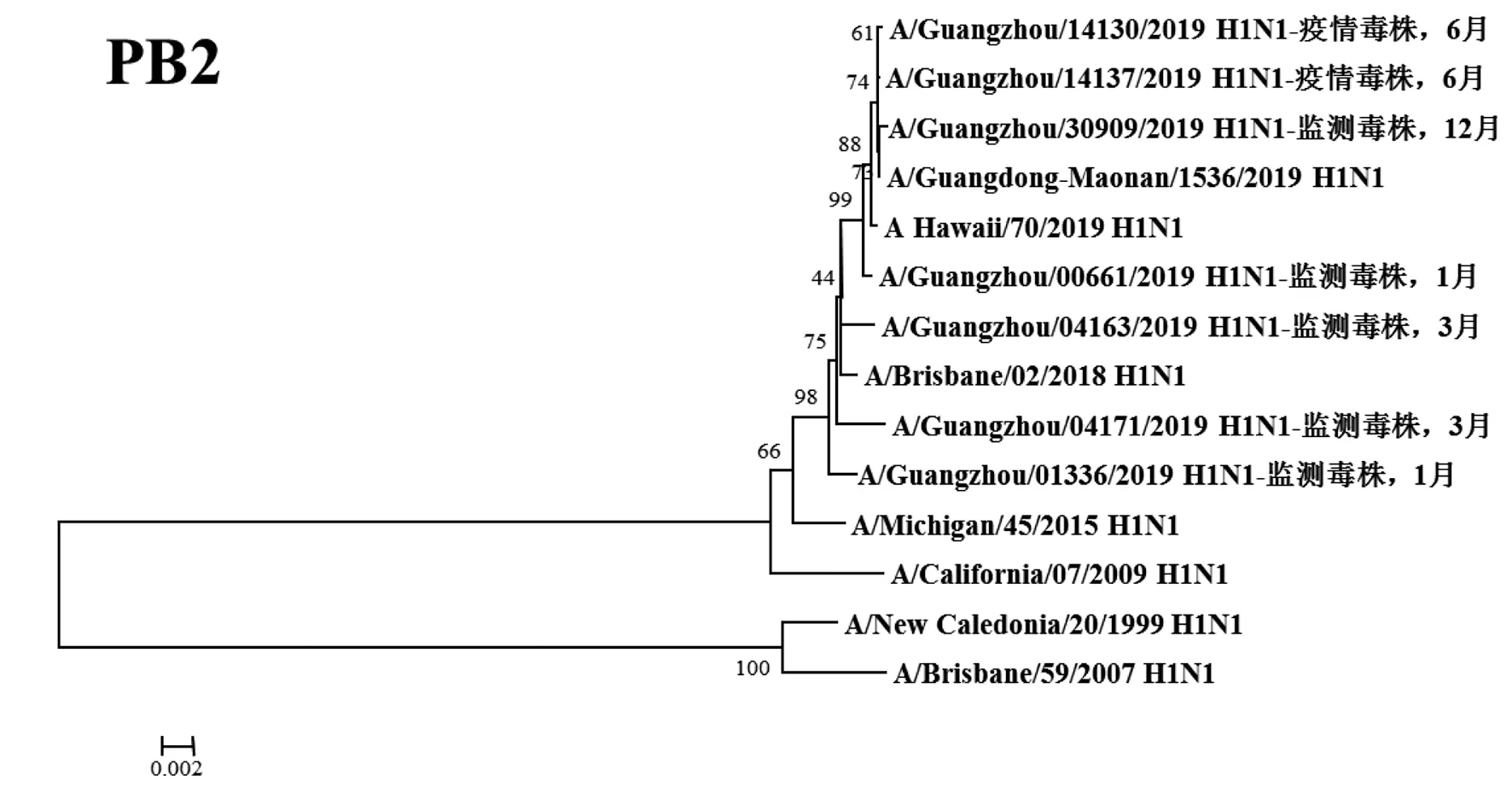
圖3 2019年廣州市甲型H1N1流感分離株P(guān)B2基因遺傳進(jìn)化樹Fig.3 Genetic evolution analysis of PB2 genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019

圖4 2019年廣州市甲型H1N1流感分離株P(guān)B1基因遺傳進(jìn)化樹Fig.4 Genetic evolution analysis of PB1 genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019
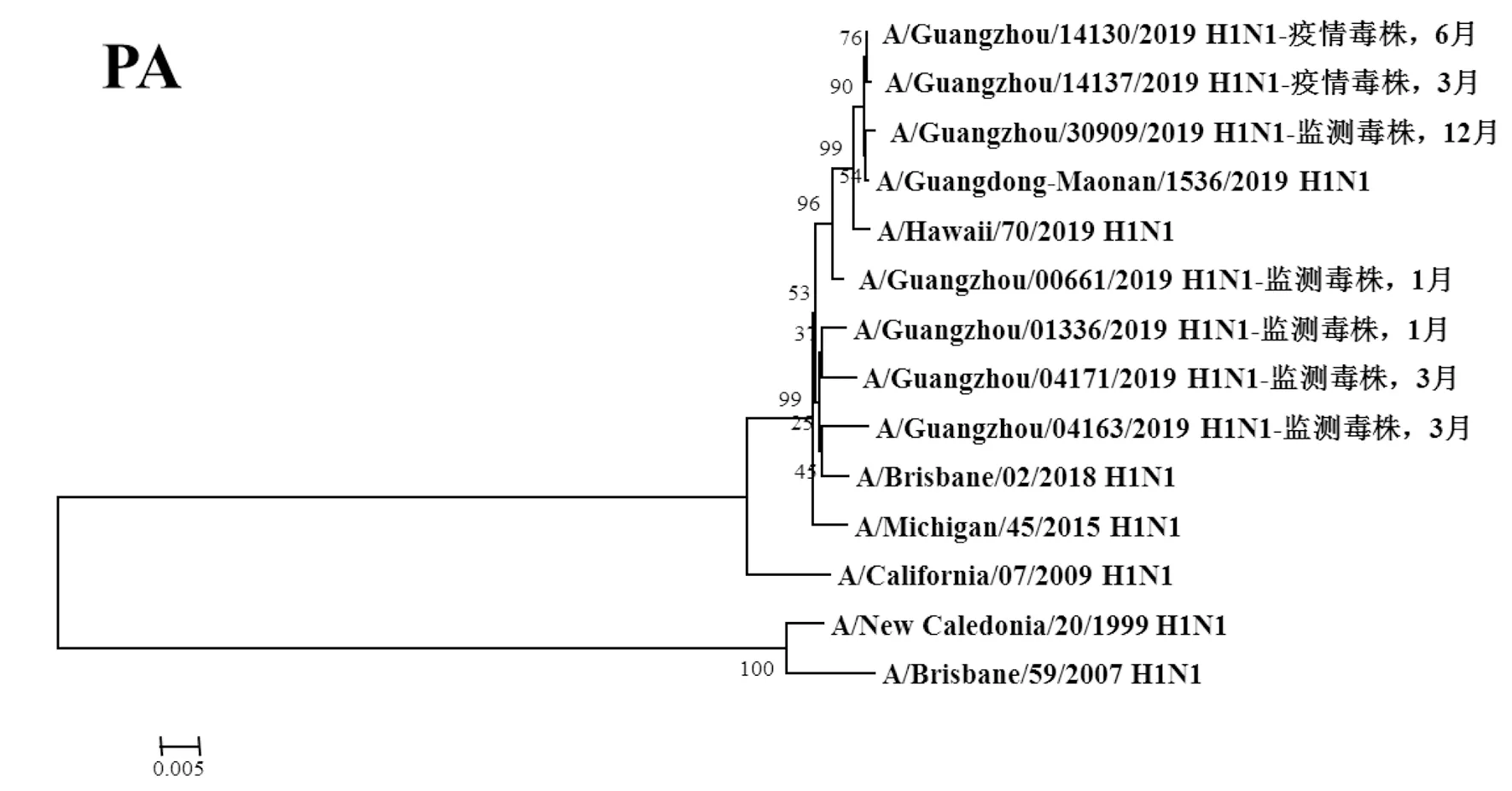
圖5 2019年廣州市甲型H1N1流感分離株P(guān)A基因遺傳進(jìn)化樹Fig.5 Genetic evolution analysis of PA genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019
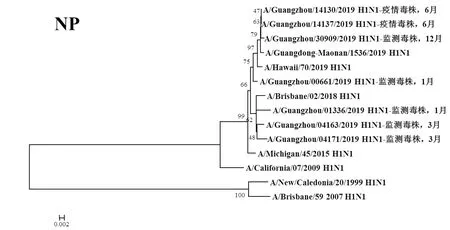
圖6 2019年廣州市甲型H1N1流感分離株NP基因遺傳進(jìn)化樹Fig.6 Genetic evolution analysis of NP genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019
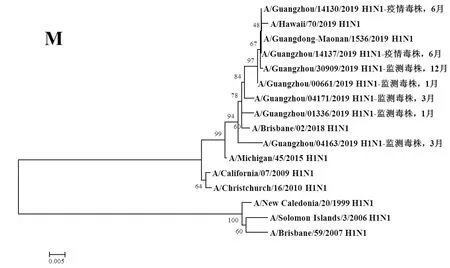
圖7 2019年廣州市甲型H1N1流感分離株M基因遺傳進(jìn)化樹Fig.7 Genetic evolution analysis of M genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019
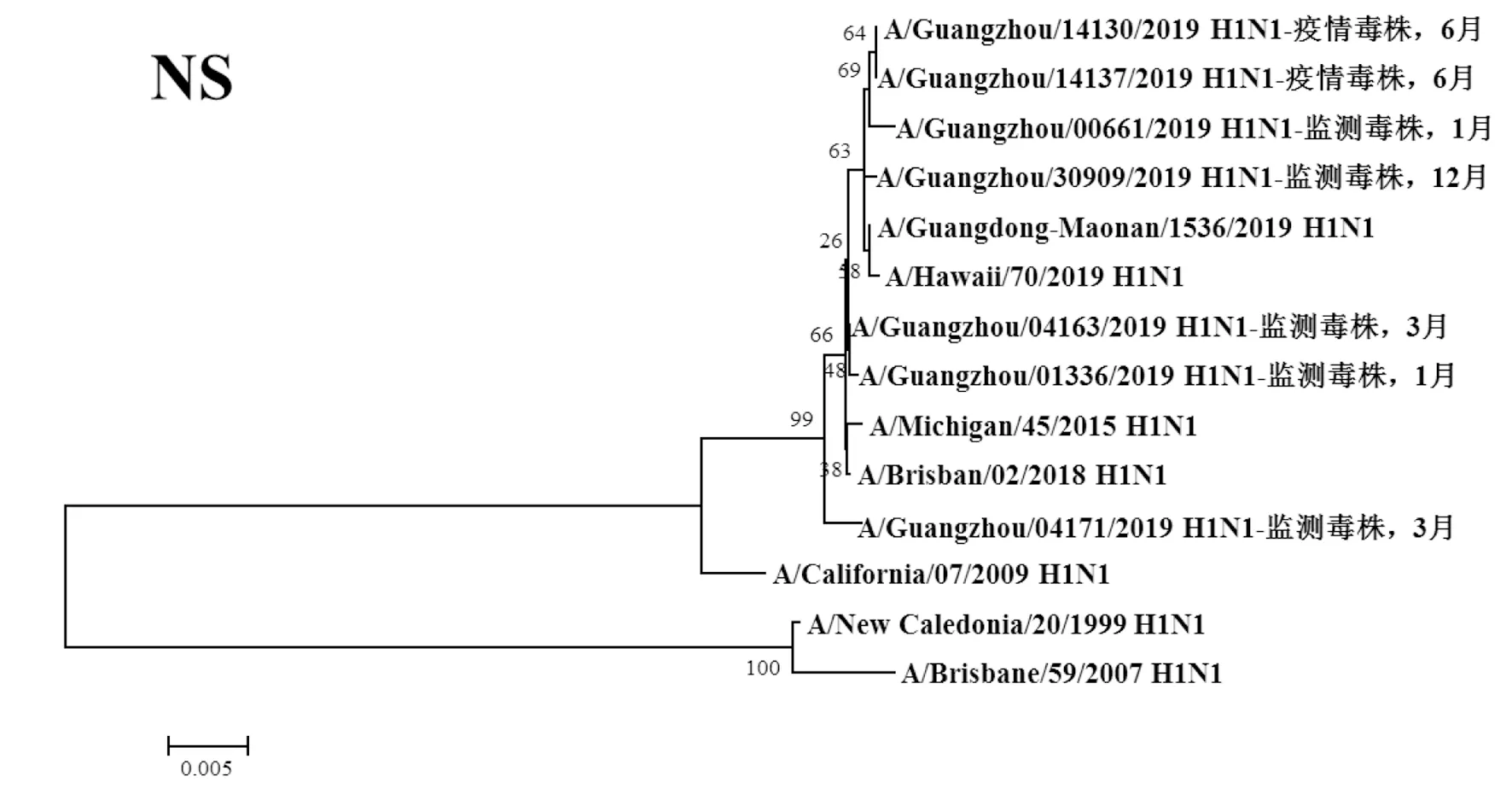
圖8 2019年廣州市甲型H1N1流感分離株NS基因遺傳進(jìn)化樹Fig.8 Genetic evolution analysis of NS genes from novel influenza A (H1N1)pdm09 isolates in Guangzhou in 2019
2.4全基因分子特征分析 HA蛋白抗原位點(diǎn)分析結(jié)果顯示,與2020-2021年疫苗推薦株A/Guangdong-Maonan/1536/2019 (H1N1)相比,01336、04163、04171分離株發(fā)生I202T變異,14130、14137、30390分離株發(fā)生D204A變異。大部分毒株HA蛋白有7個(gè)糖基化位點(diǎn),04171分離株發(fā)生104-NGT糖基化位點(diǎn)缺失。在NA蛋白神經(jīng)氨酸酶抑制劑耐藥位點(diǎn)上,所有分離株未發(fā)生耐藥突變。
根據(jù)全基因進(jìn)化樹結(jié)果顯示,A/Guangzhou/00661/2019 (H1N1)、A/Guangzhou/14130/2019 (H1N1)、A/Guangzhou/14137/2019 (H1N1)和A/Guangzhou/30309/2019 (H1N1)毒株全基因片段均與我國2020-2021年疫苗推薦株A/Guangdong-Maonan/1536/2019 (H1N1)歸屬同一進(jìn)化分支,對上述4株分離株毒株全基因氨基酸位點(diǎn)與另3株分離株進(jìn)行變異分析,結(jié)果顯示,除PA、M1、M2和NS2蛋白外,其余蛋白均有不同數(shù)量的相同特異性氨基酸突變,見表3。同時(shí)對兩株流感疫情分離株A/Guangzhou/14130/2019 (H1N1)和 A/Guangzhou/14137/2019 (H1N1)進(jìn)行全基因氨基酸變異位點(diǎn)特異性分析顯示,兩毒株具有HA-K188E、NA-I40T、PB1-M646L、NS1-I145V等共同的氨基酸變異位點(diǎn)。
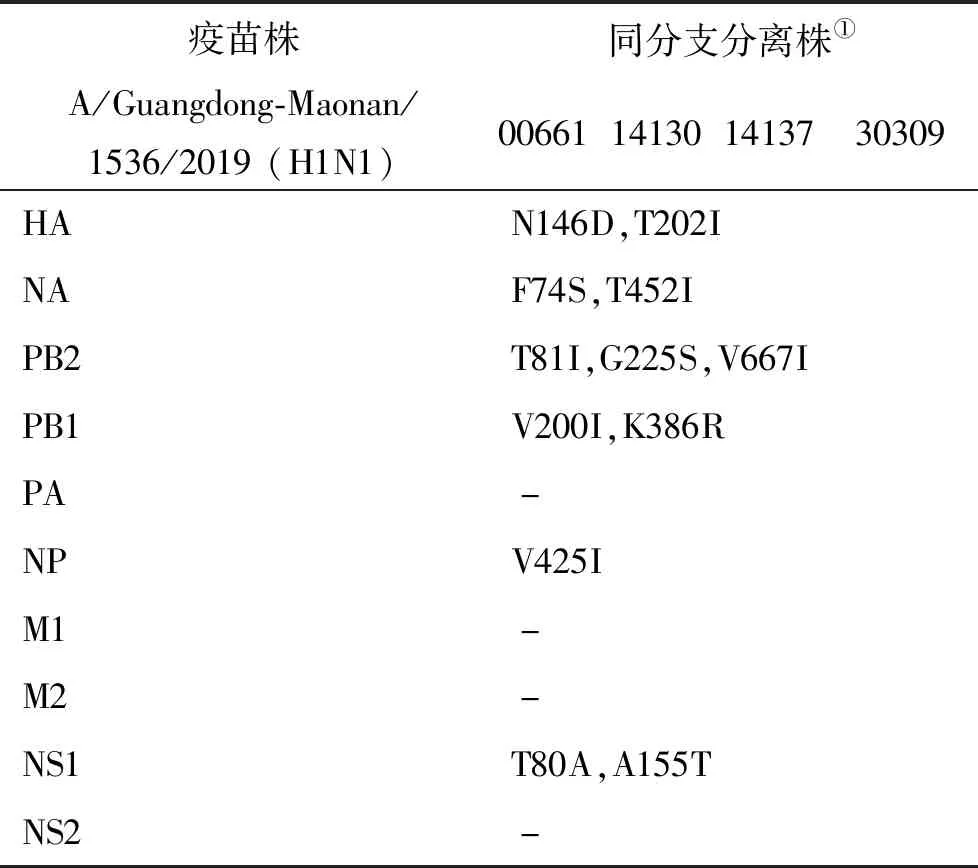
表3 同分支分離株基因組特異性氨基酸變異分析Tab.3 Analysis of genome specific amino acid variations in homocladistic isolates
3 討 論
自2017年開始,WHO推薦北半球使用的3株H1N1疫苗株(A/Michigan/45/2015 (H1N1)(2017-2019)、A/ Brisbane/02/2018 (H1N1)(2019-2020)、A/Guangdong-Maonan/1536/2019 (H1N1)(2020-2021))親緣關(guān)系較近,遺傳進(jìn)化上不斷進(jìn)化,2020-2021年疫苗株與2019年6月之后的流行株在全基因組上位于同一遺傳進(jìn)化簇上,提示2020年下半年啟動(dòng)使用的疫苗株與流行株匹配性較好。但該分支中00661毒株分離于2019年1月,說明廣州地區(qū)甲型H1N1流感病毒流行的復(fù)雜性,不排除較早期流行株與近期流行株共同流行的情況,進(jìn)而存在病毒重配風(fēng)險(xiǎn),因此及時(shí)監(jiān)測流行的變異情況、評估疫苗株與流行株的匹配性尤為重要。
廣州流感流行表現(xiàn)為一年兩次的流行高峰,本研究發(fā)現(xiàn)6月份以后的流行株全基因組序列在遺傳進(jìn)化上與疫苗株(2020-2021)共同形成獨(dú)立進(jìn)化分支,進(jìn)一步氨基酸位點(diǎn)分析也顯示,該分支分離株與疫苗株具有多個(gè)位點(diǎn)相同的特異性氨基酸突變,揭示2019年下半年流行株與疫苗株匹配性較好的分子基礎(chǔ)。本研究中2株流感疫情流行株與門診監(jiān)測流行株高度同源,進(jìn)化起源相同。但對2株疫情分離株全基因組氨基酸變異分析顯示,在HA、NA、PB1和NS1蛋白存在不同于監(jiān)測毒株的特異性氨基酸變異,這些位點(diǎn)是否與病毒傳播能力相關(guān),進(jìn)而引發(fā)流感疫情暴發(fā),需要進(jìn)一步研究。
由于流感病毒基因組分節(jié)段的特點(diǎn),當(dāng)不同亞型流感病毒共同感染宿主細(xì)胞時(shí)可發(fā)生基因重配[5],這也是流感病毒不斷進(jìn)化的重要方式之一,新重配的病毒常引起人間流感的大流行,在歷史上有3次流感大流行就是由重配流感病毒所引起[6-7]。2009年引起全球廣泛流行的pdm2009 H1N1(本研究中也稱“甲型H1N1”)即是由南美H1N1禽流感病毒的PB2、PA基因,人H3N2流感病毒的PB1基因、南美H1N1經(jīng)典豬流感病毒的HA、NP、NS和歐亞類禽H1N1豬流感病毒的NA、M基因重配進(jìn)化而來[8]。大量研究發(fā)現(xiàn),pdm2009 H1N1流感病毒宿主范圍多樣,可以感染火雞[9]、犬[10]、貓[11-13]、野鳥[14]等,提示pdm2009 H1N1流感病毒有重配產(chǎn)生新型流感病毒的可能[15]。因此開展甲型H1N1流感病毒全基因組進(jìn)化分析對新型重配病毒的發(fā)現(xiàn)有著重要意義。在本研究中所選取的9株2019年廣州市甲型H1N1流感病毒未發(fā)現(xiàn)不同基因來源的基因重配現(xiàn)象。2020年新型冠狀病毒在全球廣泛流行,我國及時(shí)啟動(dòng)了全面且有效的防疫對策,在采取高風(fēng)險(xiǎn)地區(qū)封閉管理、學(xué)校和托幼機(jī)構(gòu)休學(xué)、加強(qiáng)佩戴口罩和衛(wèi)生消毒等一系列防疫措施后,我國的新冠病毒傳播得以有效控制,同時(shí)也控制了包括甲型H1N1流感病毒在內(nèi)的呼吸道病原體的流行和傳播。2020年廣州市流感監(jiān)測數(shù)據(jù)顯示,截止目前尚未監(jiān)測到甲型H1N1流感病毒,但隨著復(fù)工復(fù)產(chǎn)、人流活動(dòng)的增加,下一階段是否會發(fā)生甲型H1N1流感病毒流行,以及繼續(xù)2019年下半流行株持續(xù)流行,還是在此基礎(chǔ)上出現(xiàn)新變異株,需要持續(xù)監(jiān)測。
利益沖突:無
