麻櫟和閩楠幼苗葉功能性狀及生物量對光照和施肥的響應(yīng)
2021-04-25 08:24:10徐海東成向榮凌高潮鄭文華胡衛(wèi)明虞木奎
生態(tài)學(xué)報 2021年6期
徐海東,熊 靜,成向榮,凌高潮,鄭文華,胡衛(wèi)明,虞木奎, *
1 中國林業(yè)科學(xué)研究院亞熱帶林業(yè)研究所,華東沿海防護(hù)林生態(tài)系統(tǒng)國家定位觀測研究站,杭州 311400 2 浙江省開化縣林場,開化 324300
光是影響植物生長發(fā)育和形態(tài)建成必需的生態(tài)因子,葉片是植物接收光照的主要器官,也是植物進(jìn)行光合作用和外界氣體交換的重要場所[1]。葉形態(tài)結(jié)構(gòu)可直接反映植物對不同光環(huán)境的適應(yīng)特征,葉功能性狀與植物生長策略和植物對資源的利用能力具有緊密聯(lián)系[2]。研究表明林下充足光照環(huán)境中植物會提高光能利用率,增加光合產(chǎn)物,增大葉片生物量,傾向于資源獲取策略;在弱光環(huán)境下植物往往表現(xiàn)出與之相反的性狀特征,生存策略轉(zhuǎn)變?yōu)橘Y源保守型[3-4]。因此,了解不同光環(huán)境下植物葉功能性狀的差異,有助于闡明樹種個體生存策略機(jī)制。在森林生態(tài)系統(tǒng)中,幼苗比成年樹種對環(huán)境因子(尤其是光照)更加敏感,且不同種類幼苗對光環(huán)境適應(yīng)策略和能力的差異成為促進(jìn)森林群落物種多樣性的有效機(jī)制[5]。徐飛等[6]研究認(rèn)為適度遮蔭有利于麻櫟和刺槐幼苗生長,而頡洪濤等[7]對5種常綠木本幼苗的研究發(fā)現(xiàn),所有植物總生物量和總氮磷積累均隨光照的增強(qiáng)呈升高趨勢。因此,理解不同生活型幼苗對不同光環(huán)境響應(yīng)特性,對于森林生態(tài)系統(tǒng)的可持續(xù)管理、人促更新以及植被的生態(tài)恢復(fù)起著重要的作用[8]。
氮和磷是控制陸地生態(tài)系統(tǒng)中植物生長和初級生產(chǎn)力的關(guān)鍵限制性營養(yǎng)元素[9]。然而,人類活動加劇了化石燃料的大量燃燒和氮肥的過量使用,導(dǎo)致全球生態(tài)系統(tǒng)活性氮輸入增加[10]。同時,我國(尤以亞熱帶地區(qū)最為嚴(yán)重)已成為全球第三大氮沉降區(qū)[11]。而磷主要來源于礦藏,相對于氮源是有限的,在許多生態(tài)系統(tǒng)中氮磷比正在逐漸增大,并可能加劇植物磷限制,從而導(dǎo)致植物表型的變化[12-13]。但是人為磷添加,即適當(dāng)降低氮磷供應(yīng)比在一定程度上可提高土壤有效氮和有效磷的含量,改善植物的養(yǎng)分狀況,增強(qiáng)植物在逆境脅迫下的生存能力[14]。因此,氮磷施肥試驗不僅可以闡明模擬全球變化(如氮磷比增加)對植物功能性狀特性及生存策略的影響機(jī)制,而且可能會提高植物對環(huán)境異質(zhì)性的適應(yīng)能力。目前,氮磷施肥試驗多集中于氮磷供應(yīng)量的影響效應(yīng),對氮磷供應(yīng)比例影響效應(yīng)的研究較少,且相關(guān)研究多集中于草地生態(tài)系統(tǒng),缺乏關(guān)于森林生態(tài)系統(tǒng)的相關(guān)研究[15]。總之,光照和氮磷供應(yīng)比會影響植物的生長,但是較少報道光照和氮磷供應(yīng)比交互作用對植物生長的影響[16]。
麻櫟(QuercusacutissimaCarruth.)作為重要的生物質(zhì)能源和用材樹種,在我國分布極為廣泛,也是營造水土保持林的優(yōu)良落葉闊葉樹種。閩楠(Phoebebournei(Hemsl.)Yang)是我國珍貴用材樹種,兼具防風(fēng)、防火和固土等多種生態(tài)屬性,也是中亞熱帶地區(qū)重點(diǎn)發(fā)展的常綠闊葉樹種。我國亞熱帶地區(qū)以松杉等針葉人工林為主,尤其是大面積的人工純林結(jié)構(gòu)簡單、生態(tài)系統(tǒng)綜合效益較低,在現(xiàn)有針葉人工林內(nèi)引入闊葉樹種改善林分結(jié)構(gòu),對提高生態(tài)系統(tǒng)功能和效益具有重要意義[17]。林木幼苗期對環(huán)境異質(zhì)性變化較為敏感,直接影響林木早期成活及生長[18]。因此,通過模擬控制試驗,研究不同光照和不同氮磷供應(yīng)比施肥(以下簡稱施肥)對麻櫟和閩楠幼苗期葉功能性狀及生物量影響,明確兩種典型闊葉樹種對異質(zhì)環(huán)境的適應(yīng)策略,為今后林分結(jié)構(gòu)調(diào)控過程中樹種選擇和配置提供參考。
1 材料與方法
1.1 試驗區(qū)概況
試驗開展于中國林科院亞熱帶林業(yè)研究所內(nèi)虎山生態(tài)學(xué)試驗基地(30°05′N,119°57′E)。該地區(qū)屬亞熱帶季風(fēng)濕潤氣候,年均降水量1415.7 mm,年均氣溫16.4℃,年日照時數(shù)1816 h,土壤類型主要為紅黃壤。
1.2 試驗材料和設(shè)計
選擇生長狀況基本一致的麻櫟和閩楠1年生容器苗各96棵(由浙江省開化縣林場提供,試驗開始前麻櫟和閩楠樹高分別為(50.0±2.1)cm和(13.3±1.8)cm,試驗結(jié)束時麻櫟和閩楠的樹高范圍是(51.5±7.3)—(115.5±1.6)cm和(20.8±4.2)—(69.3±8.1)cm)進(jìn)行盆栽實驗。盆栽試驗土按照生黃土∶蛭石4∶1的比例配制,土壤基本化學(xué)性質(zhì)如下:pH 4.87,有機(jī)質(zhì)2.86 mg/kg,全氮0.24 g/kg,全磷0.11 g/kg,水解性氮20.53 mg/kg,有效磷0.51 mg/kg。每個花盆(35 cm×25 cm×15 cm)內(nèi)裝基質(zhì)3.5 kg。施肥所用氮肥和磷肥分別為硝酸銨(NH4NO3)和磷酸二氫鉀(KH2PO4);采用50%透光度的黑色尼龍遮蔭網(wǎng)進(jìn)行遮蔭處理。
2019年3月中旬,將麻櫟和閩楠容器苗移栽到裝有試驗土的花盆,每盆一株。待常規(guī)管理及緩苗30 d后,將苗木置于溫室大棚內(nèi)。于4月中旬以及5月初分別進(jìn)行光照和施肥試驗。試驗采用雙因素(光照和施肥)隨機(jī)區(qū)組設(shè)計,光照和施肥共8個處理,每個處理12盆苗木,共192盆苗木。其中,光照處理采用自然全光照和45%全光照(由遮蔭網(wǎng)下實測光強(qiáng)和自然全光照比值計算);施肥采用氮磷比5,氮磷比15和氮磷比45三個典型梯度的處理方法[19],并將不施肥處理作為對照(CK)。每個處理速效態(tài)氮肥含量均為100 mg/kg,通過控制速效態(tài)磷肥含量形成3個氮磷比處理水平:5(NP5)、15(NP15)和45(NP45)。將氮磷肥混合為水溶液(現(xiàn)配現(xiàn)用),在一個月內(nèi)完成5次均等施肥試驗,試驗期間對所有苗木進(jìn)行統(tǒng)一管理。
1.3 光合氣體交換參數(shù)測定
于2019年4月中旬選擇晴朗無云天(9:00—11:00)利用便攜式光合測定儀LI-6400進(jìn)行葉片氣體交換參數(shù)的測定。測量時設(shè)定光照強(qiáng)度為1000 μmol m-2s-1(經(jīng)測定,麻櫟和閩楠均達(dá)到光飽和狀態(tài))。2019年10月初于每個處理選擇麻櫟和閩楠幼苗(各5株)樹冠中端展開的成熟健康葉片(各3片)進(jìn)行凈光合速率(Net photosynthetic rate,Pn)、蒸騰速率(Transpiration rate,Tr)、氣孔導(dǎo)度(Stomatal conductance,Gs)、胞間CO2濃度(Intercellular CO2concentration,Ci)參數(shù)的測定,瞬時水分利用效率(Water use efficiency,WUE)=凈光合速率/蒸騰速率。
1.4 葉綠素?zé)晒鈪?shù)測定
光合氣體交換參數(shù)測定后,利用PAM-2500便攜式脈沖調(diào)制熒光儀(WALZ,德國)進(jìn)行葉片葉綠素?zé)晒馓匦缘臏y定。測定前先對植株葉片進(jìn)行30 min充分暗適應(yīng),測定的葉綠素?zé)晒鈪?shù)包括:最大光化學(xué)效率(Maximum photochemical efficiency of PSII,Fv/Fm)、實際光合效率(Actual photochemical efficiency of PSⅡ,Y(II))、調(diào)節(jié)性能量耗散的量子產(chǎn)額(Quantum yield of regulated energy losses,Y(NPQ))、非調(diào)節(jié)性能量耗散的量子產(chǎn)額(Quantum yield of non-regulated energy losses,Y(NO))、非光化學(xué)猝滅系數(shù)(Non-photochemical quenching coefficient,NPQ)、光化學(xué)猝滅系數(shù)(Photochemical quenching coefficient,qP)和電子傳遞速率(Electron transport rate,ETR)。
1.5 生物量和葉形態(tài)性狀指標(biāo)測定
2019年10月中旬試驗結(jié)束后,于每個處理中隨機(jī)選取6棵平均木進(jìn)行生物量和葉形態(tài)性狀的測定。首先,在每株苗木上隨機(jī)選取10片健康完整的葉子用手持葉面積儀準(zhǔn)確掃描和測量葉面積。并將掃描后的葉片、未掃描葉片、枝干以及地下所有根系于65℃烘箱烘干至恒重并稱重。比葉面積(Specific leaf area,SLA)=掃描葉面積/掃描葉干重,葉干物質(zhì)含量(Leaf dry matter content,LDMC)=掃描葉干重/掃描葉飽和鮮重。同時計算地上生物量(Aboveground biomass,AGB)、地下生物量(Belowground biomass,BGB)、總生物量(Total biomass,TB)=地上部生物量+地下部生物量、根冠比(Root shoot ratio,RSR)=地下部生物量/地上部生物量,最后根據(jù)單株葉生物量計算單株總?cè)~面積(Leaf area,LA)。
1.6 葉氮含量測定
將稱取干重的葉片研碎,單位質(zhì)量葉氮含量(Unit mass leaf nitrogen content,Nmass)用濃硫酸-過氧化氫消煮-半微量凱氏定氮法測定。單位面積葉氮含量(Unit area leaf nitrogen content,Narea)=單位質(zhì)量葉氮含量×10/比葉面積,光合氮利用效率(Photosynthetic nitrogen use efficiency,PNUE)=凈光合速率/單位面積葉氮含量。
1.7 數(shù)據(jù)處理方法
采用SPSS 18.0進(jìn)行數(shù)據(jù)進(jìn)行統(tǒng)計分析。首先,利用Duncan進(jìn)行同一樹種不同處理下相同指標(biāo)的差異顯著性檢驗(α=0.05)。其次,通過一般線性模型量化樹種、光照、施肥及其交互作用對葉功能性狀和生物量的影響。最后,利用PCA和Pearson相關(guān)性法分析生物量和葉功能性狀間關(guān)系。上述可視化分析均用R(Version 3.6.3)完成,表中數(shù)據(jù)為平均值±標(biāo)準(zhǔn)差(Mean±SD)。
2 結(jié)果與分析
2.1 光照和施肥對葉功能性狀的影響
2.1.1光合氣體交換參數(shù)
與全光照處理相比,遮蔭顯著降低了麻櫟Tr和閩楠Pn,顯著增加了麻櫟WUE(表1,P<0.05)。全光照處理下麻櫟的Pn、Gs和Ci隨氮磷供應(yīng)比的增大而顯著升高(P<0.05),而遮蔭處理僅Pn隨氮磷比增大而增加。麻櫟Gs、Tr、PNUE在遮蔭氮磷比15時最低,WUE在全光照和遮蔭下均在CK中最低;在全光照下,閩楠光合氣體交換參數(shù)(除Tr外)均隨氮磷比的增大而升高,而遮蔭下,僅Pn和PNUE隨氮磷比增大而增加。閩楠的Gs、Ci和Tr在遮蔭氮磷比15時最低,而WUE和PNUE在氮磷比5時最低。此外,在全光照下,閩楠僅平均水分利用效率高于麻櫟(增加了1.9倍)。而遮蔭下,麻櫟平均蒸騰速率和光合氮利用率分別是閩楠1.8倍和2.4倍。
2.1.2葉綠素?zé)晒鈪?shù)
由表2可知,在全光照下,與CK相比,施肥顯著增加了麻櫟的Y(II)、qP和ETR,同時也顯著降低了Y(NPQ)和NPQ(P<0.05)。在遮蔭下,與CK相比,施肥顯著降低了麻櫟qP和ETR(P<0.05);與全光照CK相比,施肥僅顯著降低了閩楠Y(NO),但在氮磷比45時顯著增加了NPQ和Fv/Fm(P<0.05)。與遮蔭CK相比,氮磷供應(yīng)比15和45顯著增加了閩楠Y(NPQ)和Y(NO),但顯著降低了Y(II)、qP和ETR(P<0.05)。在所有處理中,麻櫟Y(II)、qP和ETR均值都低于閩楠,其余指標(biāo)差異較小。
2.1.3葉形態(tài)性狀及氮含量
與全光照處理相比,遮蔭顯著降低了麻櫟LA,但顯著增加了SLA和Nmass(P<0.05)。在全光照處理中,與CK相比,施肥顯著增加了麻櫟Nmass(P<0.05),并在氮磷比45時達(dá)到最大值(表3)。在遮蔭處理中,施肥對麻櫟葉形態(tài)性狀及Narea的影響不顯著(P>0.05);與全光照處理相比,遮蔭顯著降低了閩楠LA(P<0.05)。在全光照處理中,與CK相比,施肥顯著增加了閩楠LA(P<0.05),對其余指標(biāo)整體影響較小。在遮蔭處理中,閩楠僅LDMC在氮磷比為5時顯著低于CK,其余指標(biāo)在不同施肥處理間差異不顯著(P>0.05)。此外,相同處理下,閩楠僅全光照下Nmass和遮蔭下Narea整體高于麻櫟。
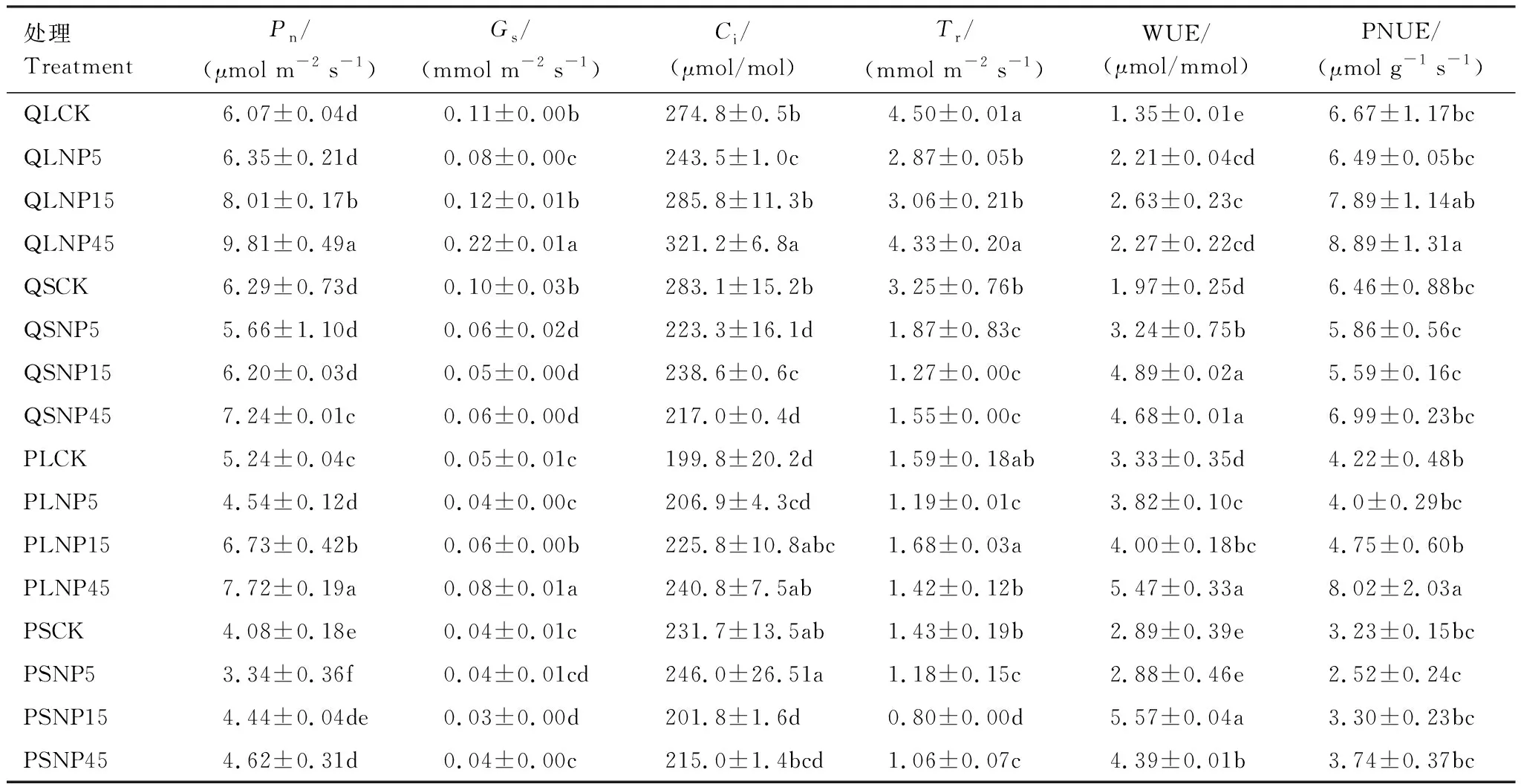
表1 光照和施肥處理下麻櫟和閩楠的光合氣體交換參數(shù)
Pn: 凈光合速率Net photosynthetic rate;Gs: 氣孔導(dǎo)度 Stomatal conductance;Ci: 胞間CO2濃度 Intercellular CO2concentration;Tr: 蒸騰速率 Transpiration rate; WUE: 水分利用效率 Water use efficiency; PNUE: 光合氮利用率 Photosynthetic nitrogen use efficiency; QL: 麻櫟全光照,Quercusacutissima+Light; QS: 麻櫟遮蔭,Q.acutissima+Shading; PL: 閩楠全光照,Phoebebournei+Light; PS: 閩楠遮蔭P.bournei+Shading; CK: 對照 Control; NP5: 氮磷供應(yīng)比為5 Nitrogen and phosphorus supply ratio of 5; NP15: 氮磷供應(yīng)比為15 Nitrogen and phosphorus supply ratio of 15; NP45: 氮磷供應(yīng)比為45 Nitrogen and phosphorus supply ratio of 45; 表中不同小寫字母表示相同樹種同一指標(biāo)在不同處理間差異顯著(P<0.05)
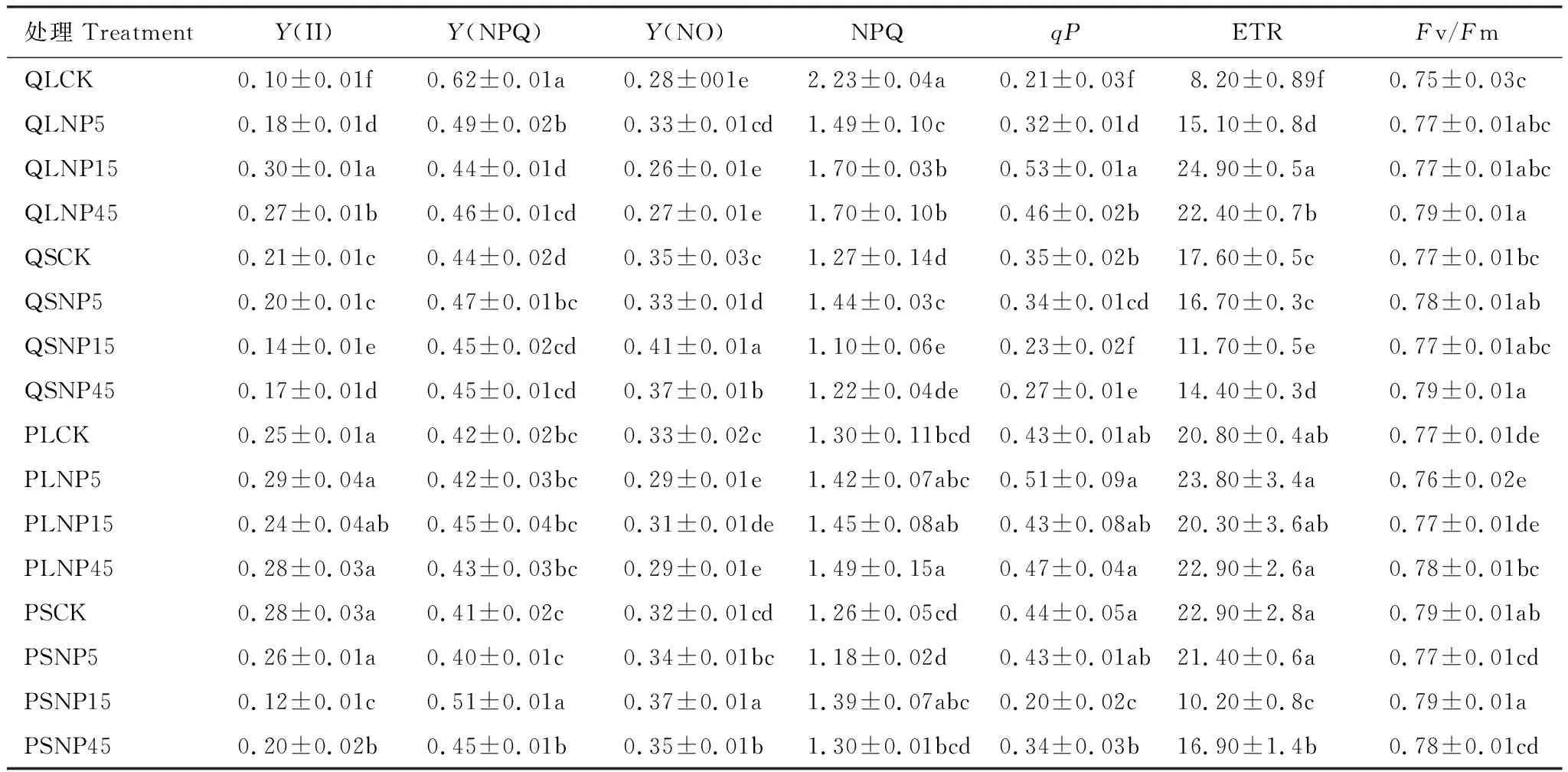
表2 光照和施肥處理下麻櫟和閩楠的葉綠素?zé)晒鈪?shù)
2.2 光照和施肥對生物量的影響
如表3所示,與全光照處理相比,遮蔭顯著降低了麻櫟A(chǔ)GB、BGB和TB及RSR(P<0.05)。在全光照處理中,與CK相比,施肥顯著增加了麻櫟A(chǔ)GB,并在氮磷比45時值最高,同時顯著降低了BGB和RSR(P<0.05)。在遮蔭處理中,施肥處理對麻櫟生物量的影響不顯著(P>0.05);與全光照處理相比,遮蔭顯著降低了閩楠AGB、BGB和TB(P<0.05)。與全光照CK相比,施肥顯著增加了全光照閩楠BGB和TB(P<0.05),其余指標(biāo)差異較小。在遮蔭處理中,閩楠生物量指標(biāo)在不同施肥處理中無顯著差異(P>0.05)。此外,相同處理下,麻櫟生物量指標(biāo)均高于閩楠。
2.3 樹種、光照和施肥及其交互作用對生物量和葉功能性狀的影響
從圖1可知,樹種和施肥對光合氣體交換參數(shù)均具有顯著影響,兩因子累計解釋量占總變異量的23.4%—62.4%,此外,光照和施肥×光照處理顯著影響Pn、Gs、Ci、Tr和PNUE (P<0.05);施肥、光照及施肥×光照對葉綠素?zé)晒鈪?shù)影響顯著(P<0.05),對Y(NO)和NPQ方差解釋量分別為54.8%和36.6%,施肥是影響Fv/Fm的主導(dǎo)因子(方差解釋量為15.2%);LA、LDMC和Nmass均受光照和施肥(光照為主)的顯著影響(P<0.05),而SLA和Narea主要受樹種影響;光照是影響生物量指標(biāo)(除RSR外)的主要因子,光照和樹種對AGB、BGB和TB的方差累計解釋量分別為82.5%、72.0%和81.1%,而RSR主要由樹種影響,其次是光照。
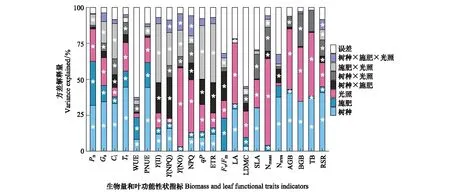
圖1 樹種、施肥和光照及其交互作用對麻櫟和閩楠生物量及葉功能性狀的變異解釋量Fig.1 The explained variance of tree species, fertilization, light and their interaction on biomass and leaf functional traits in Q. acutissima and P. bourneiPn: 凈光合速率Net photosynthetic rate; Gs: 氣孔導(dǎo)度 Stomatal conductance; Ci: 胞間CO2濃度 Intercellular CO2 concentration; Tr: 蒸騰速率 Transpiration rate; WUE: 水分利用效率 Water use efficiency; PNUE: 光合氮利用率 Photosynthetic nitrogen use efficiency; Y(II): 實際光合效率 Actual photochemical efficiency of PSⅡ; Y(NPQ): 調(diào)節(jié)性能量耗散的量子產(chǎn)額 Quantum yield of regulated energy losses ; Y(NO): 非調(diào)節(jié)性能量耗散的量子產(chǎn)額Quantum yield of non-regulated energy losses; NPQ: 非光化學(xué)猝滅系數(shù) Non-photochemical quenching coefficient; qP: 光化學(xué)猝滅系數(shù) Photochemical quenching coefficient; ETR: 電子傳遞速率 Electron transport rate; Fv/Fm: 最大光化學(xué)效率 Maximum photochemical efficiency of PSII; LA: 葉面積 Leaf area; LDMC: 葉干物質(zhì)含量 Leaf dry matter content; SLA: 比葉面積 Specific leaf area; Nmass: 單位質(zhì)量葉氮含量 Unit mass leaf nitrogen content; Narea: 單位面積葉氮含量 Unit area leaf nitrogen content; AGB: 地上生物量 Aboveground biomass; BGB: 地下生物量 Belowground biomass; TB: 總生物量 Total biomass; RSR: 根冠比 Root shoot ratio; * P < 0.05
2.4 生物量和葉功能性狀間關(guān)系
由PCA分析可知,麻櫟和閩楠兩主成分軸累計解釋率分別為77.1%和71.9%,其中,全光照和遮蔭處理在PC1軸上顯著分離。麻櫟SLA、Nmass、Y(NO)和WUE集中分布在遮蔭處理中,而其余指標(biāo)均分布在PC2軸左側(cè),且生物量指標(biāo)和Tr、LA以及NPQ關(guān)系緊密(圖2a);閩楠PNUE、Pn、LA與生物量指標(biāo)關(guān)系更為緊密,且集中分布在光照處理中(圖2b)。由Pearson相關(guān)性分析可知,在全光照處理中(圖2c),麻櫟A(chǔ)GB與LA、Y(II)和ETR顯著正相關(guān),與Y(NPQ)和NPQ顯著負(fù)相關(guān)(P<0.05)。麻櫟RSR與NPQ顯著正相關(guān),卻與LA和Pn顯著負(fù)相關(guān)(P<0.05)。在遮蔭處理中(圖2d),Ci與生物量指標(biāo)均顯著負(fù)相關(guān),BGB、TB與Fv/Fm顯著正相關(guān),AGB與LA顯著正相關(guān)(P<0.05)。在全光照處理中(圖2e),閩楠AGB與大部分光合氣體交換參數(shù)和LA正相關(guān),BGB與部分葉綠素?zé)晒鈪?shù)和LA正相關(guān),TB與Ci和WUE顯著正相關(guān)(P<0.05)。在遮蔭處理中(圖2f),閩楠AGB、BGB和TB與LA顯著正相關(guān),而BGB、TB和RSR與SLA和Nmass均顯著負(fù)相關(guān)(P<0.05)。
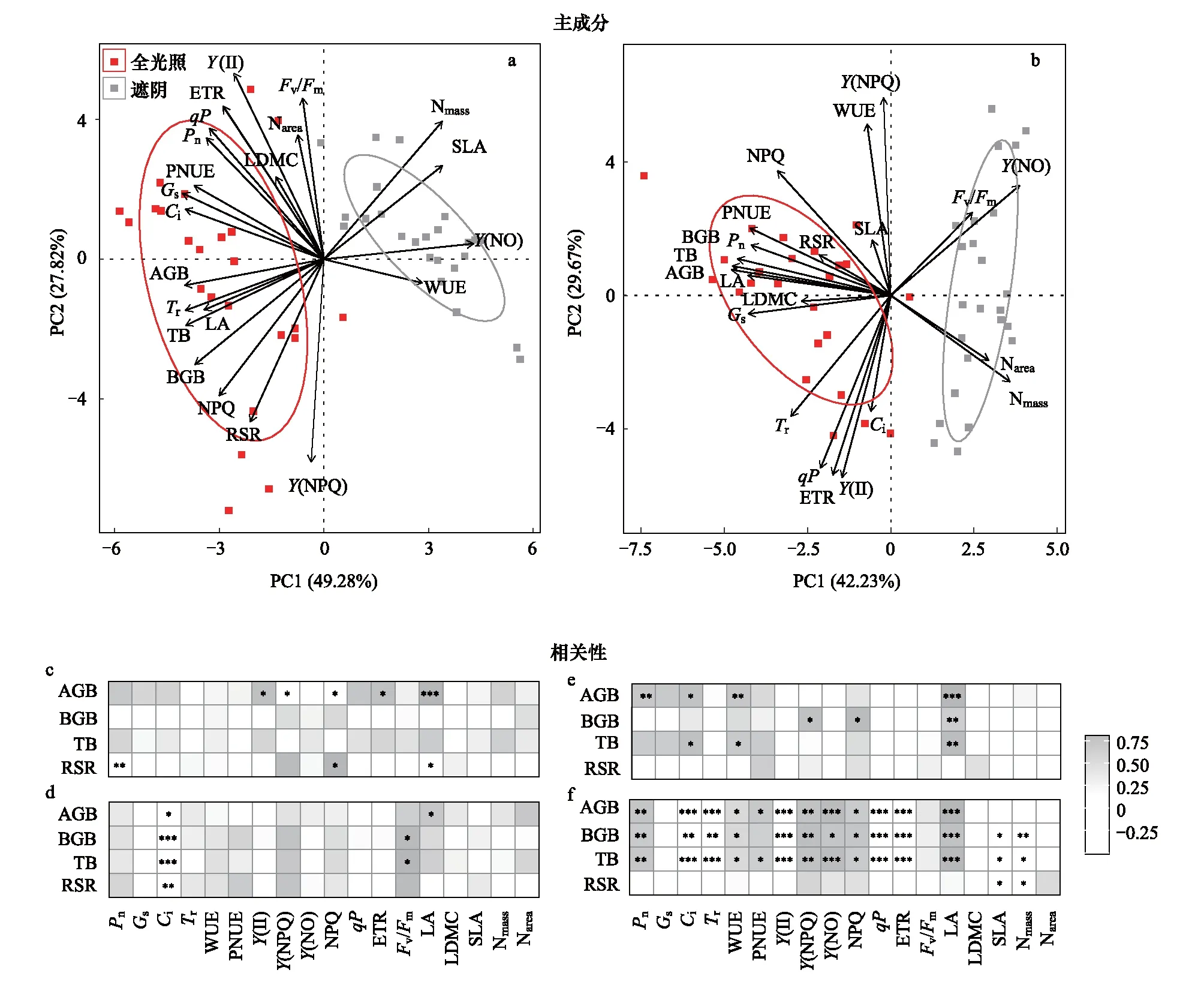
圖2 麻櫟、閩楠生物量和葉功能性狀間的主成分及相關(guān)性分析Fig.2 The correlation and principal component analysis among plant biomass and leaf functional traits in Q. acutissima and P. bournei* P < 0.05,** P < 0.01,*** P < 0.001
3 討論
光合作用是植物生長發(fā)育最重要的生理過程,受自然環(huán)境條件如光照和土壤養(yǎng)分異質(zhì)性的影響[5]。本研究發(fā)現(xiàn),遮蔭僅顯著影響了麻櫟WUE、Tr和閩楠Pn,尤其對閩楠光合氣體交換參數(shù)的整體影響較小。說明麻櫟和閩楠在遮蔭環(huán)境下具有較強(qiáng)的光合生理適應(yīng)能力,其中,閩楠在弱光環(huán)境下可能具有更穩(wěn)定的光合能力或更強(qiáng)的生理調(diào)節(jié)能力[20]。葉片Pn可反應(yīng)植物的光合能力,進(jìn)而決定植物的生產(chǎn)力[21]。通過線性回歸分析發(fā)現(xiàn)樹種和施肥對Pn方差累計解釋量超過60%。在相同光照處理下,麻櫟Pn、Gs、Ci和Tr普遍高于閩楠,而Gs的增大會增加植物對Ci的吸收與儲存,從而增加植物光合作用底物。類似的,Tr增大會使得光合產(chǎn)物輸出加快,通過反饋促進(jìn),來提高麻櫟光合效能[22]。此外,麻櫟和閩楠Pn隨氮磷供應(yīng)比增大而升高,并在氮(100 mg/kg)磷(2.2 mg/kg)比45時達(dá)到最大值,而袁義福[23]研究發(fā)現(xiàn)麻櫟Pn在氮(1350 mg/kg)磷(90 mg/kg)比15時值最高,研究結(jié)果的差異可能是由氮磷供應(yīng)量的不同導(dǎo)致的,但具體原因有待于進(jìn)一步試驗驗證。PNUE是描述植物葉片養(yǎng)分利用、生理特性和生存策略的重要指標(biāo)。本研究發(fā)現(xiàn)樹種是影響PNUE最重要的因子,且常綠樹種閩楠表現(xiàn)出較低的PNUE,但低PNUE樹種在土壤貧瘠或者低溫環(huán)境中往往更占優(yōu)勢[24]。WUE可反映植物對環(huán)境的適應(yīng)能力,高WUE意味著植物具有低耗水、高生產(chǎn)率和抗性強(qiáng)的特征[25]。本文發(fā)現(xiàn)施肥、樹種以及樹種×光照對WUE均有顯著影響。與不施肥處理相比,施肥顯著增加了麻櫟和閩楠WUE,普遍在氮磷比15時達(dá)到最大值,這與施肥下Tr的變化趨勢基本一致(表1)。同理,本文發(fā)現(xiàn)遮蔭處理中只有麻櫟WUE顯著高于其全光照處理,主要?dú)w因于遮蔭下麻櫟Pn的不顯著變化及Tr的顯著降低導(dǎo)致了WUE增大,且Gs的降低在一定程度上也減少了水分散失,從而間接提高了WUE[26]。這些結(jié)果說明Tr可能是調(diào)控施肥和光照處理下麻櫟和閩楠WUE變化的關(guān)鍵因子。此外,無論在光照還是遮蔭條件下,閩楠WUE均值都高于麻櫟,說明與麻櫟相比,閩楠在林下復(fù)雜光環(huán)境中可能表現(xiàn)出較高的水分利用效率,從而提高自身的競爭及生存力,這與廖德寶等[27]研究結(jié)果一致,即常綠闊葉樹比落葉闊葉樹表現(xiàn)出更高的水分利用效率。
葉綠素?zé)晒馀c光合作用密切相關(guān),常用來檢測光合機(jī)構(gòu)對環(huán)境變化的響應(yīng)[28]。本研究發(fā)現(xiàn),遮蔭對麻櫟和閩楠葉綠素?zé)晒馓匦缘挠绊戄^小,表明不同光環(huán)境下麻櫟和閩楠光合機(jī)構(gòu)穩(wěn)定性較高。與全光照不施肥處理相比,施肥顯著增加麻櫟Y(II)、qP和ETR,同時顯著降低Y(NPQ)和NPQ,說明施肥提高了麻櫟光能轉(zhuǎn)化效率和電子傳遞活性,進(jìn)而提高了PSII活性[29]。此外,全光照下麻櫟Y(II)、qP和ETR在氮磷比15和45時均顯著高于氮磷比5,即較高氮磷比施肥有利于全光照下麻櫟光能利用率的提高,此時葉片光化學(xué)效率可能主要由氮的有效性影響[30]。然而,Y(II)、qP和ETR在遮蔭且氮磷比5時均顯著高于相同處理下氮磷比15和45,NPQ則表現(xiàn)出相反的趨勢。說明在低氮磷比施肥處理下,麻櫟葉片傾向于將吸收的光能主要用于光化學(xué)傳遞,并在光照不足情況下通過調(diào)整光合機(jī)構(gòu)來提高光能捕獲量和電子傳遞速率,而磷供應(yīng)的相對增加可能是驅(qū)動遮蔭下麻櫟光合結(jié)構(gòu)活性的主要動力[31]。對閩楠而言,全光照氮磷供應(yīng)比45處理顯著降低閩楠Y(NO),但顯著提高NPQ;遮蔭氮磷供應(yīng)比45處理顯著降低閩楠Y(II)、qP和ETR,但顯著提高Y(NPQ)和Y(NO),這可能是閩楠對不同光環(huán)境及氮磷比失衡的一種適應(yīng)策略,即通過合理分配葉片吸收的光能,從而維持光合作用的正常進(jìn)行和實現(xiàn)葉片的自我保護(hù)[32]。Fv/Fm是PSII反應(yīng)中心利用光能的最大能力,在非脅迫條件下葉片F(xiàn)v/Fm一般介于0.75—0.85[33]。麻櫟和閩楠在光照和施肥處理中Fv/Fm的變化范圍為0.75—0.79,然而,施福軍[34]等人研究發(fā)現(xiàn)全光照及單一氮肥添加下閩楠幼苗Fv/Fm均低于0.75,結(jié)果的差異可能是由于施肥方式的不同造成的,同時說明氮磷比配施可能更有利于維持苗木光合生理機(jī)構(gòu)的穩(wěn)定性。此外,測定季節(jié)及苗木年齡也會影響試驗結(jié)果,其具體原因有待進(jìn)一步研究。
葉形態(tài)和化學(xué)性狀可反應(yīng)植物的資源利用策略及其對生存環(huán)境的適應(yīng)能力。本研究發(fā)現(xiàn),光照處理和樹種類型主要影響葉形態(tài)性狀和葉氮含量,其中,光照對葉形態(tài)指標(biāo)的影響效應(yīng)遠(yuǎn)高于施肥效應(yīng)(圖1)。此外,與不施肥處理相比,施肥僅顯著增加了麻櫟Nmass和閩楠LA,而光照處理顯著影響了麻櫟LA、SLA和Nmass以及閩楠LA,說明麻櫟和閩楠幼苗對光照變化敏感度高于施肥處理,這與武高林等[35]的研究結(jié)果一致,即光照異質(zhì)性對植物幼苗葉形態(tài)性狀的影響高于養(yǎng)分異質(zhì)性。盡管如此,施肥處理下麻櫟和閩楠LA均高于不施肥處理,說明施肥在一定程度可增大LA,進(jìn)而積累更多光合產(chǎn)物[36],這也符合本文Pn的變化趨勢。與全光照處理相比,遮蔭顯著提高了麻櫟SLA,高SLA可以使麻櫟在弱光環(huán)境下獲得更多的光能,增強(qiáng)麻櫟的存活能力[32]。閩楠SLA在光照處理中(尤其在遮蔭下)遠(yuǎn)低于麻櫟,說明閩楠的耐蔭性要高于麻櫟[37]。此外,遮蔭下麻櫟和閩楠Nmass的升高(相比于全光照處理),說明營養(yǎng)元素氮在葉片光合結(jié)構(gòu)投資較多,這有利于樹種加快生長以競爭林下有限光資源[38]。
生物量是反映植物生長和生產(chǎn)力的一個重要內(nèi)容,植物通過改變生物量及根冠比來適應(yīng)光照或養(yǎng)分環(huán)境的變化[39]。本研究發(fā)現(xiàn)麻櫟通過降低根冠比的策略來適應(yīng)光照和養(yǎng)分異質(zhì)性環(huán)境,如此以獲取更多的光照資源,提高自身競爭力。但是麻櫟在光照和施肥處理中采取了不同的生長策略,與全光照相比,遮蔭雖然顯著降低了麻櫟地上、地下及總生物量,但為適應(yīng)弱光環(huán)境及權(quán)衡總生物量的適合度麻櫟傾向于降低地下生物量分配來維持地上器官生長優(yōu)勢;與全光照不施肥處理相比,施肥處理中麻櫟采取了降低地下生物量以及提高地上生物量來減小根冠比的生態(tài)策略。相反,閩楠在所有處理中根冠比均無顯著變化,這與閩楠自身生物量分配可塑性低有關(guān)[39]。除了植物自身特異性差異影響之外,光照和施肥及其交互作用也表現(xiàn)出對生物量指標(biāo)的顯著影響(圖1)。麻櫟和閩楠對不同光環(huán)境采取了相似的生存策略,即全光照條件下,兩樹種通過快速資源獲取策略,積累了更多生物量,而在遮蔭條件下采取了緩慢資源獲取的保守型策略,積累了較少的生物量,該結(jié)果符合Toledo-Aceves等[40]關(guān)于不同光環(huán)境下植物生物量分配規(guī)律。此外,通過PCA以及Pearson相關(guān)性分析發(fā)現(xiàn),LA可作為不同光環(huán)境下植物生物量變化的預(yù)測因子,同時LA也是氮磷比施肥處理中驅(qū)動麻櫟A(chǔ)GB及閩楠AGB、BGB和TB變化的關(guān)鍵形態(tài)指標(biāo)。全光照下植物單株總?cè)~面積的增加,會增加其受光面積及光合產(chǎn)物的積累,從而提高植株的地上、地下及整體生產(chǎn)力[36]。但是隨著施肥梯度的變化,與閩楠相比,麻櫟總?cè)~面積和生物量(除AGB外)間關(guān)系可能逐漸解耦(圖2c和2d),說明麻櫟葉性狀和生物量關(guān)聯(lián)性受施肥處理影響較大。此外,本文發(fā)現(xiàn)施肥處理組中不同光環(huán)境下麻櫟生物量指標(biāo)和葉功能性狀間顯著相關(guān)關(guān)系要遠(yuǎn)少于閩楠,尤其是遮蔭處理中閩楠(圖2e和2f),說明氮磷比施肥會促進(jìn)閩楠在弱光環(huán)境下表現(xiàn)出更加穩(wěn)定且多元化的適應(yīng)策略來調(diào)控生產(chǎn)力[41]。
4 結(jié)論
本文重點(diǎn)分析了光照和施肥及其交互處理下麻櫟和閩楠幼苗生物量和葉功能性狀的響應(yīng)特性,以及生物量與葉功能性狀間的關(guān)聯(lián)性,主要結(jié)論如下:(1)麻櫟和閩楠的生長對光照異質(zhì)性更加敏感,并采取了相似的生長適應(yīng)策略。但在弱光環(huán)境下,閩楠表現(xiàn)出更為多元化且穩(wěn)定的生長適應(yīng)策略。(2)光照或遮蔭下施肥對麻櫟和閩楠總生物量均具有一定的促進(jìn)作用。(3)單株總?cè)~面積是麻櫟和閩楠地上生物量變化的重要指示指標(biāo),即可間接借助易測的葉面積代替硬性狀(如光合生理、元素含量等)來篩選具有較高生產(chǎn)力的樹種。總之,麻櫟和閩楠幼苗均可在較復(fù)雜的光照和養(yǎng)分條件下生存,通過適當(dāng)增加光照(如林分結(jié)構(gòu)優(yōu)化和調(diào)控)和施肥(如施加含有較高氮磷比例的林產(chǎn)有機(jī)物料)可以提高麻櫟和閩楠幼苗的生存力、競爭力和生產(chǎn)力。這為林下復(fù)雜光環(huán)境和養(yǎng)分條件下引入樹種的篩選、配置及幼苗期管理提供了理論依據(jù)。但鑒于本研究局限于大棚控制實驗,相關(guān)結(jié)論及發(fā)現(xiàn)需要更多研究來確定其可靠性和實用性。
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
