深圳灣浮游植物群落結構特征及其與環境因子的關系
2021-04-12 06:35:04陳思陳海剛田斐李藝彤張林寶張喆王學鋒蔡文貴
生態科學 2021年1期
陳思, 陳海剛, 田斐, 李藝彤, 張林寶, 張喆, 王學鋒, 蔡文貴, *
深圳灣浮游植物群落結構特征及其與環境因子的關系
陳思1, 2, 3, 陳海剛2, 田斐2, 李藝彤2, 張林寶2, 張喆2, 王學鋒4, 蔡文貴2, *
1. 上海海洋大學水產與生命學院, 上海 201306 2. 中國水產科學研究院南海水產研究所, 廣東省漁業生態環境重點實驗室, 農業部南海漁業資源環境重點野外科學觀測實驗站, 廣東珠江口生態系統野外科學觀測研究站, 廣東, 廣州 510300 3. 廣州彭加木紀念中學, 廣東, 廣州 510430 4. 南方海洋科學與工程廣東省實驗室(湛江), 廣東, 湛江 524025
為研究深圳灣海域浮游植物的群落結構及其與水環境的關系, 分別于春夏秋冬(2013年8月至2014年4月)四個季度對深圳灣進行采樣分析, 并對調查區域內的浮游植物及其主要環境因子進行了相關性分析。結果表明, 深圳灣海域共鑒定出浮游植物111種(包括變種和變型):硅藻35屬86種, 甲藻4屬15種, 綠藻6屬6種, 藍藻3屬4種, 其中硅藻門是主要的優勢類群, 占總種數的77.5%, 其次是甲藻門, 占總種數的13.5%, 綠藻門和藍藻門分別占總種數的5%和4%。浮游植物種類組成以廣溫廣鹽種和暖水種為主, 中肋骨條藻(Skeletonema costatum)是該海域常年優勢種。深圳灣海域浮游植物細胞豐度年波動范圍為28.64—462.46×104cells·m–3, 平均值為156.43×104cells·m–3, 其季節變化趨勢為秋季>夏季>冬季>春季。浮游植物多樣性指數和均勻度指數變化范圍分別為0.11—4.06和0.04—0.90, 多樣性指數和均勻度指數均偏低, 群落結構趨于單一化, 穩定性差。浮游植物細胞豐度與環境因子的相關性關系在不同季節有顯著差別, 其中無機氮與浮游植物細胞豐度之間呈現正相關關系。
深圳灣;浮游植物; 群落結構; 環境因子
0 前言
深圳灣為珠江口伶仃洋東側中部的一個內寬外窄的半封閉淺水海灣, 位于深圳經濟特區和香港特別行政區之間, 海灣水域面積90.8 km2, 平均水深不超過2.9 m, 最大水深不超過5 m[1]。深圳灣生態系統是華南地區具有國際意義的最重要的濕地生態系統之一。近20年來, 深圳灣兩岸的經濟快速發展, 給深圳灣海域帶來了極大的環境壓力, 深圳灣海灣富營養化[2]、赤潮頻發[3]、細菌污染[4]等問題不斷出現, 海灣生態系統遭受到嚴重的破壞, 嚴重影響到市民的生產生活。
浮游植物作為海洋生態系統中的初級生產者, 其群落結構變化與環境因子密切相關, 浮游植物可對環境條件變化迅速做出響應, 其種類組成、數量和優勢種在不同的水環境中變化很大[5–7]。深圳灣海水富營養化程度很高, 高度富營養化的水體使得深圳灣海域赤潮頻發[8]。對深圳灣浮游植物的重視源于1988年春季環節環溝藻赤潮的研究, 王朝暉等[9]分析了深圳灣環節環溝藻赤潮發生的原因。孫金水[10]等(2006)分析了深圳灣海域浮游植物的分布情況, 發現深圳灣浮游植物細胞豐度呈明顯雙周期變化, 平面分布表現為由海灣中部向灣口遞減的格局。溫度和營養鹽是限制浮游植物生長的重要環境因子[11–12]。張才學[12]研究了2008年深圳灣海域浮游植物和營養鹽間的關系, 從優勢種的種類數和多樣性指數分析, 深圳灣浮游植物的群落結構已趨于單一化, 生態系統抗干擾能力極為脆弱。本文根據2013—2014年4個季度航次的現場浮游植物資料, 分析了深圳灣海域浮游植物的種類組成、豐度季節分布特征、生物多樣性和群落結構, 探討了海洋環境因子與浮游植物群落關系, 為進一步研究該海區浮游植物群落生態變化積累資料, 并為深圳灣生態環境保護和海洋資源可持續利用提供科學依據。
1 材料與方法
1.1 采樣時間與站位
在深圳灣海域設置了15個調查站位, 于2013年8月到2014年4月對深圳灣海域浮游植物群落的季節變化進行研究, 調查分4個航次進行, 分別為2013年8月(夏季)、2013年11月(秋季)、2014年1月(冬季)和2014年4月(春季)。
1.2 樣品的采集、測定和鑒定
1.2.1 樣品的采集與分析
定性樣品用使用淺水Ⅲ型浮游生物網, 采用垂直拖網方式采樣, 樣品用中性甲醛溶液(濃度為采集樣品的 4%—5%)固定, 定性樣品沉淀24 h后, 搖勻后取中間層水樣制作臨時封片, 在顯微鏡 (Olympus CX31)下觀察浮游植物形態特征并對浮游植物進行定性鑒定。浮游植物定量樣品用有機玻璃采水器取水樣1 L裝入聚乙烯瓶中, 水樣采集后, 立即用魯哥氏液固定。水樣經過48 h兩次沉淀后被濃縮至20—30 mL, 取0.1 mL均勻樣品在光學顯微鏡下進行種類鑒定和細胞計數。樣品處理方法依照《海洋調查規范第6部分:海洋生物調查》(GB 12763.6—2007)要求進行。
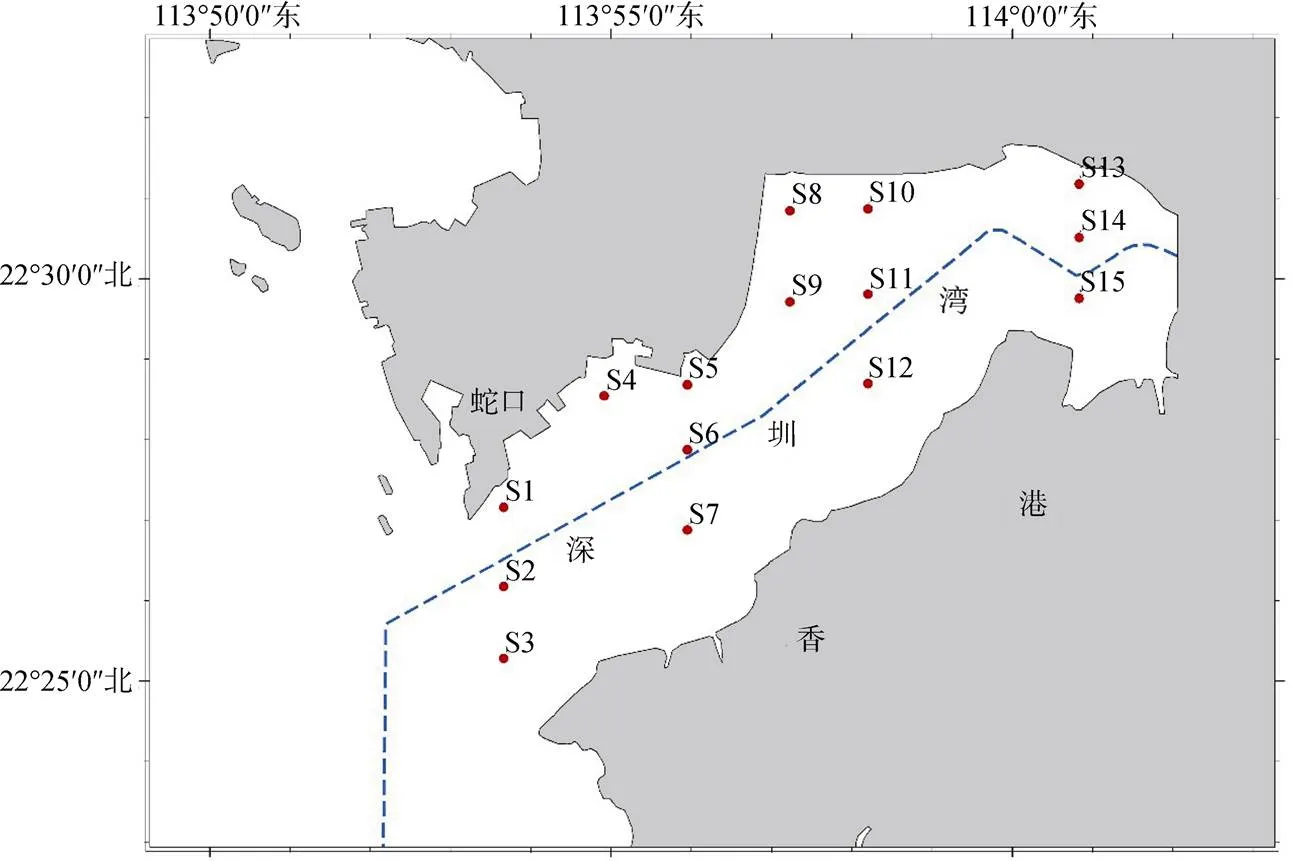
圖1 深圳灣調查站位
Figure 1 Locations of sampling sites in Shenzhen Bay
對同步環境要素進行監測, 監測內容包括溫度、鹽度、溶解氧、無機氮和活性磷酸鹽等, 監測方法按照《海洋監測規范第4部分:海水分析》(GB17378.4—2007)的要求進行。
1.2.2 生物多樣性分析
(1)浮游植物優勢度()應用以下公式計算:
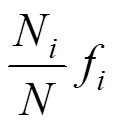
式中:n為第種的個體數;f是該種在各站中出現的頻率;為所有站每個種出現的總個體數, 定義值大于 0.02 的種類為本次調查的優勢種。
(2)采用Shannon—Weaner指數[13]測定浮游植物的多樣性指數, 其計算公式為:
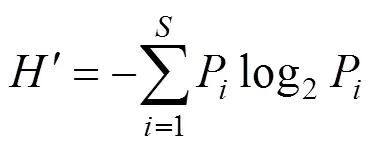
式中:′為種類多樣性指數;為樣品中的種類總數;為第種的個體數與總個體數的比值。
(3)采用Pielou[14]均勻度測定浮游植物的均勻度, 其公式為:
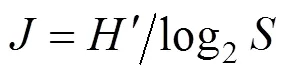
式中:為均勻度;′為種類多樣性指數;為樣品中的種類總數。
浮游植物與環境因子的Pearson相關性分析采用SPSS軟件; 細胞豐度分布圖采用ArcGIS軟件完成; 其余插圖用Origin2017完成; 此外, 采用馬建新[15]提出的浮游植物多樣性評判閾值對深圳灣海域浮游植物多樣性水平進行評價。
2 結果與分析
2.1 浮游植物的種類與組成
深圳灣海域共鑒別出浮游植物111種, 其中, 硅藻35屬86種, 甲藻4屬15種, 綠藻6屬6種, 藍藻3屬4種。深圳灣海域浮游植物的主要類群是硅藻, 占總種數的77.5% ; 其次是甲藻, 占總種數的13.5%, 其他種類占總種類數9%。硅藻門中以角毛藻屬()種類最多(18種), 占硅藻總種數的20.9%, 其次為圓篩藻屬()(13種)和菱形藻屬()(8種), 分別占硅藻總種數的15.1%和22.9%。甲藻門中以角藻屬()(9種)為主, 占甲藻總種數的60%。4個季節的調查中, 浮游植物種類季節變化明顯, 冬季種類數最高, 為59種, 其次是春季, 為57種, 夏季、秋季各為27和25種。浮游植物的群落組成以沿岸廣布種為主, 同時出現較多咸淡海種類。浮游植物種類組成及分布見圖2。
2.2 浮游植物細胞豐度
浮游植物細胞豐度季節性變化顯著, 波動范圍為28.64—462.46×104cells·m–3, 平均為156.43×104cells·m–3。秋季的平均細胞豐度最高, 為462.46×104cells·m–3, 夏季和冬季平均細胞豐度較接近, 分別為76.32×104和58.29×104cells·m–3, 春季細胞豐度最小, 僅為28.64×104cells·m–3。深圳灣海域浮游植物細胞豐度在秋季出現了明顯的峰值, 在春季細胞豐度達到最低值, 屬于單峰型。總體上, 深圳灣浮游植物細胞豐度呈現西岸(深圳一側)高于東岸(香港一側), 灣內高于灣口的趨勢。
深圳灣海域春夏秋冬四個季節, 硅藻門的平均豐度遠高于其他浮游植物, 分別占相應季節的95.99%、93.12%、91.20%和98.86%, 甲藻門和藍藻門所占比例均低于硅藻門, 深圳灣海域主要浮游植物為硅藻門。
2.3 浮游植物的優勢種和季節演替
調查結果顯示, 深圳灣海域共出現浮游植物優勢種14種, 其中春季7種; 夏季4種; 秋季2種; 冬季3種(表3)。4個季度均為優勢種的種類沒有, 中肋骨條藻在三個季度都是優勢種。優勢度最高的是秋季的中肋骨條藻, 占細胞總量的83.99%; 其次是夏季的小型月牙藻(), 占細胞總量的42.64%; 冬季第一優勢種為中肋骨條藻, 其優勢度為0.565, 占細胞總量的56.46%; 春季以冕孢角毛藻()優勢度最高(0.122)。四個季度中, 硅藻門占所出現優勢種比例最大。這一結果與袁超[11]2010—2011年深圳灣浮游植物調查結果一致。

表1 浮游植物多樣性程度評判閾值
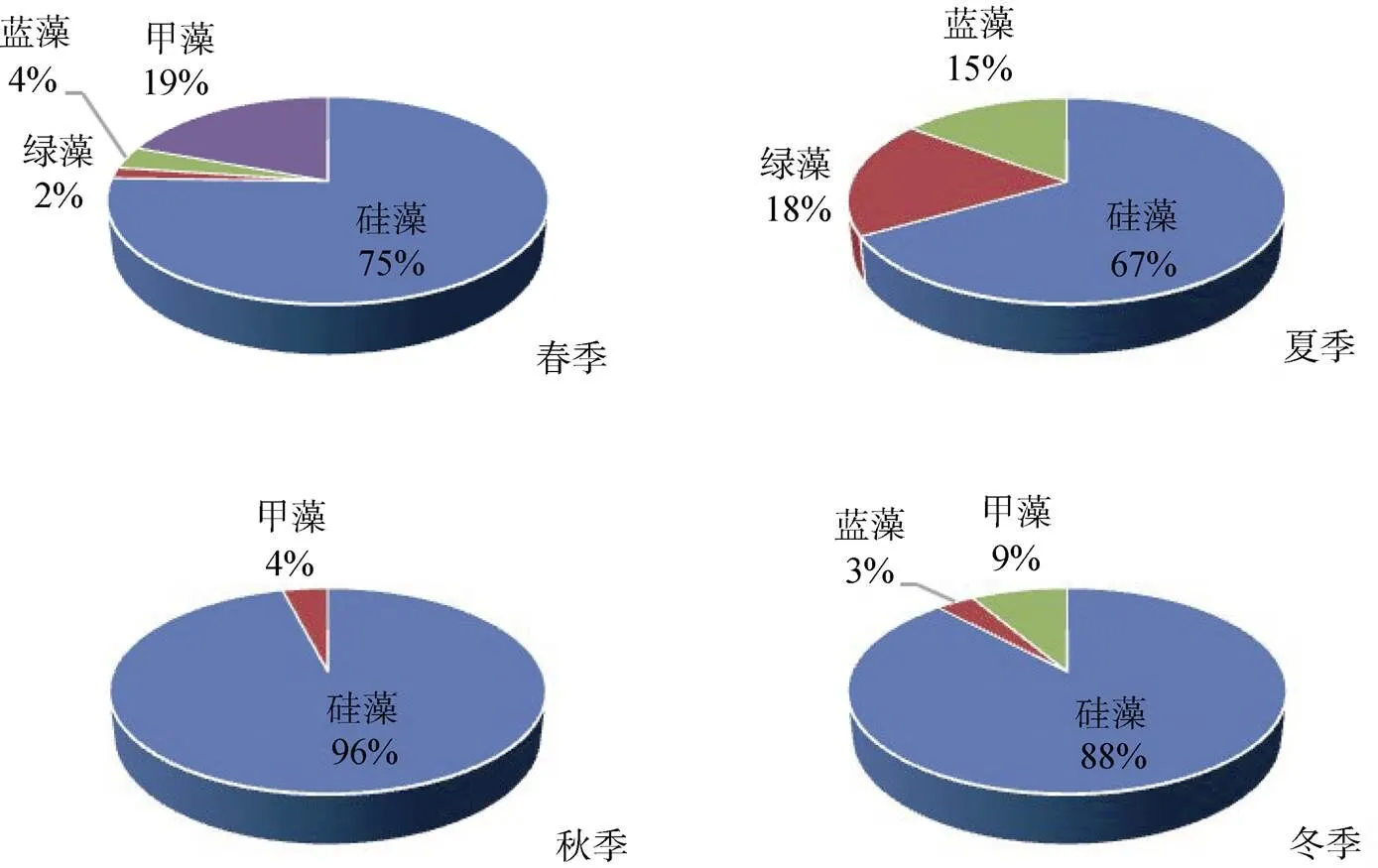
圖2 物種類組成和數量的季節變化
Figure 2 Seasonal changes of species composition and phytoplankton density
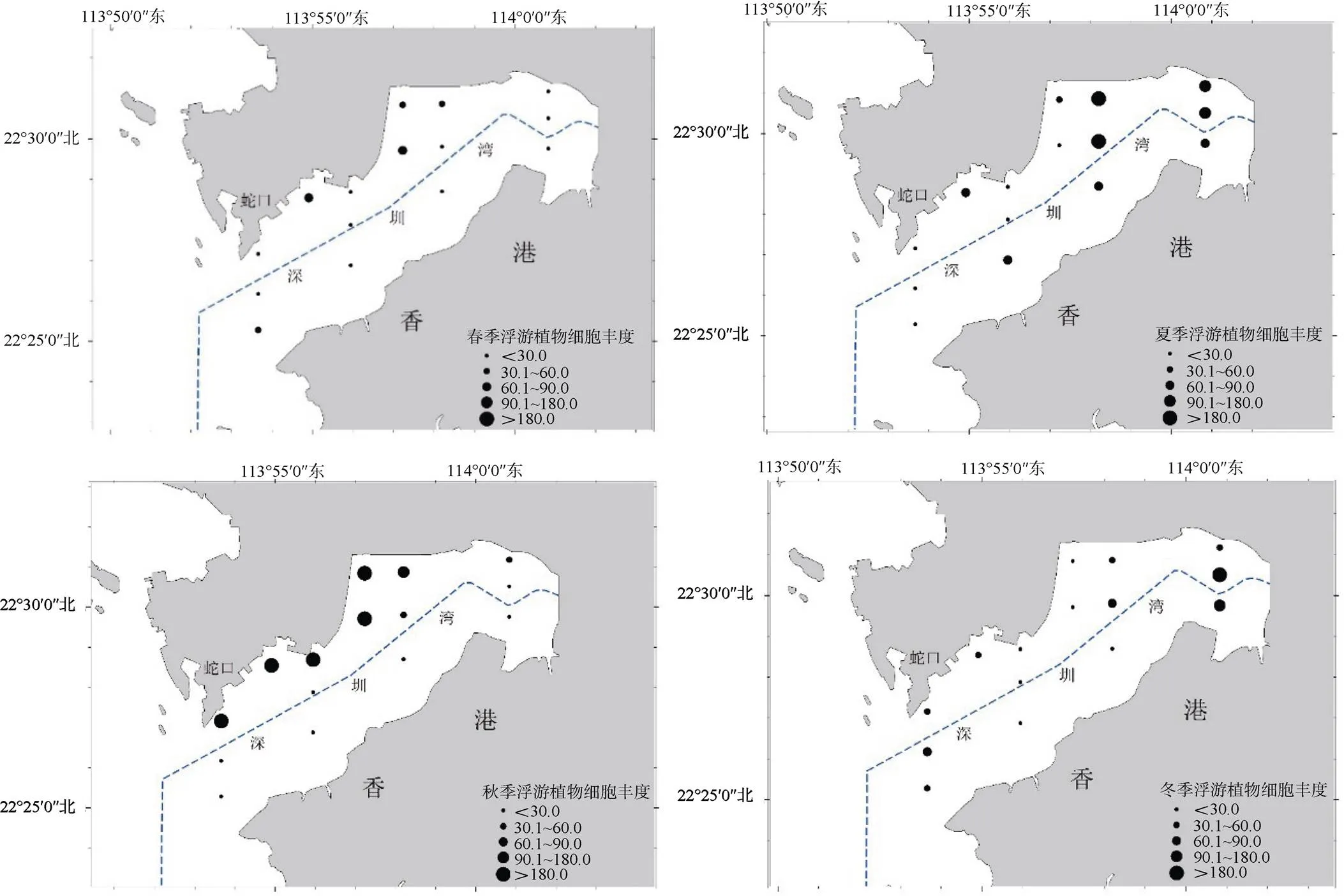
圖3 深圳灣浮游植物平面分布和季節變化
Figure 3 Horizontal distributions and seasonal changes of densities of phytoplankton in Shenzhen Bay
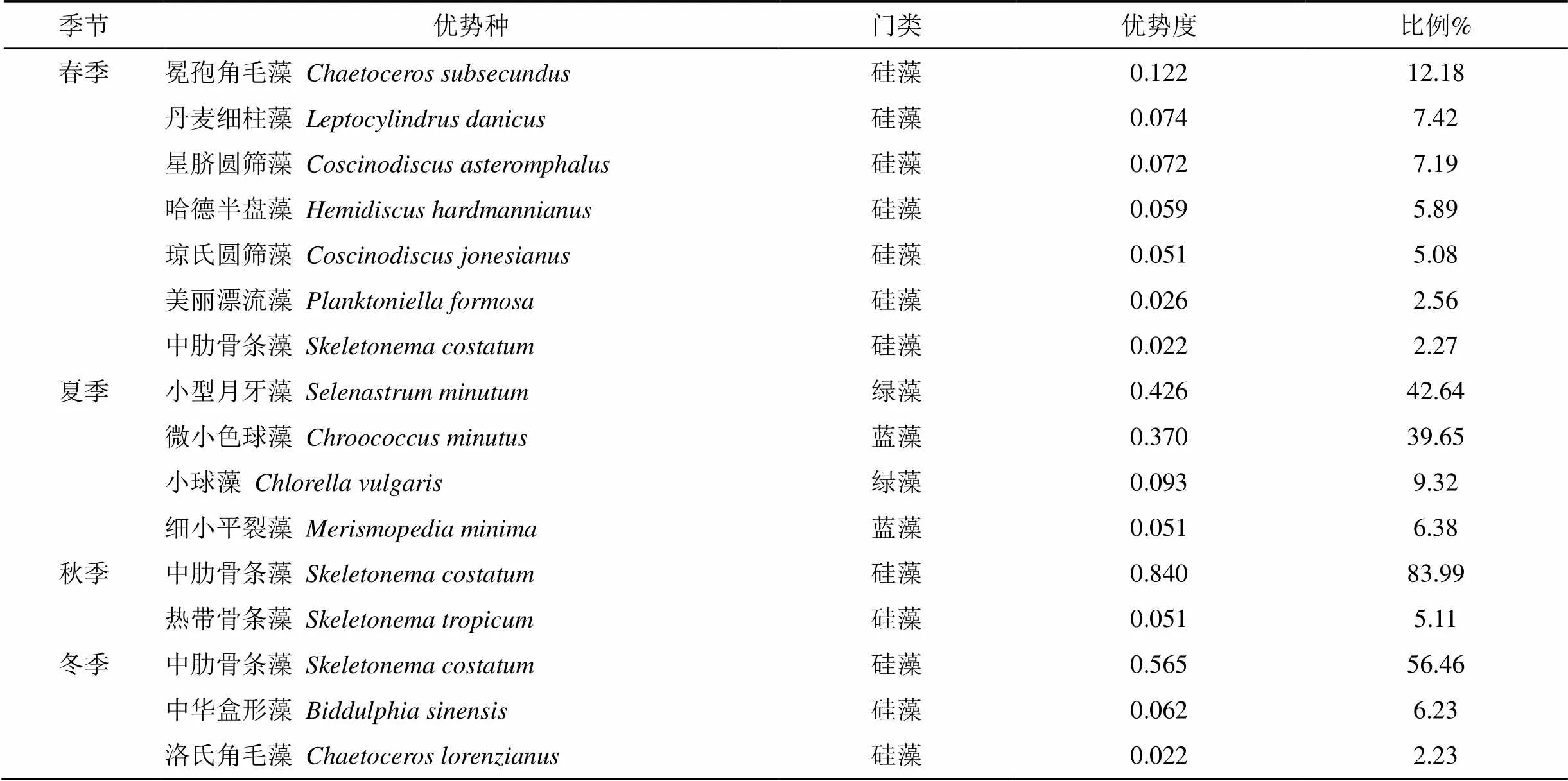
表 2 浮游植物的優勢種及在各站的細胞豐度占比(優勢度Y>0.02)
注:細胞豐度單位:×104cells·m–3。
2.4 浮游植物多樣性指數和均勻度的變化
深圳灣各站位浮游植物多樣性指數變化范圍為0.11—4.06, 在平面分布上, 深圳灣呈現灣口處浮游植物物種多樣性較高, 而灣中部和灣內處多樣性水平較低。在季節變化上, 春季最高, 平均為2.80, 夏季次之, 平均為2.00, 秋季最小, 平均僅為1.50。在春季, 多樣性指數由灣口向灣內逐漸減小; 在夏季, 灣口處多樣性指數較高, 而灣中部較小; 在秋季, 呈現西岸(深圳一側)高于東側(香港一側); 在冬季, 灣中部較高, 灣口和灣內多樣性指數較低均勻度變化范圍為0.04—0.90, 平均是春季較高(0.80), 其余季節次之, 均為0.50。
2.5 浮游植物豐度與環境因子關系
深圳灣海域水溫季節變化表現為夏季最高(28.65 ℃), 春季次之(28.59 ℃), 秋季(26.9℃)冬季最低(17.6 ℃)。鹽度季節變化表現為秋季(23.46)>冬季(23.28)>夏季(14.44)>春季(12.38), 春季海水鹽度平均值顯著低于其他季節, 僅為12.87。pH值和鹽度各站位差異較小, 其變化趨勢表現為從灣內到灣口呈逐漸升高趨勢。總氮、總磷、無機氮和磷酸鹽等表現為灣內到灣口逐漸降低趨勢。將浮游植物細胞豐度與pH、鹽度、溶解氧、和水溫等主要環境因子進行Pearson相關回歸系數分析(表4), 從表中可以看出, 夏季、秋季浮游植物細胞豐度和無機氮顯著正相關, 冬季浮游植物細胞豐度和鹽度顯著負相關, 和總氮、總磷和無機氮顯著正相關(p<0.05), 而其他環境因子與浮游植物細胞豐度相關性不大。
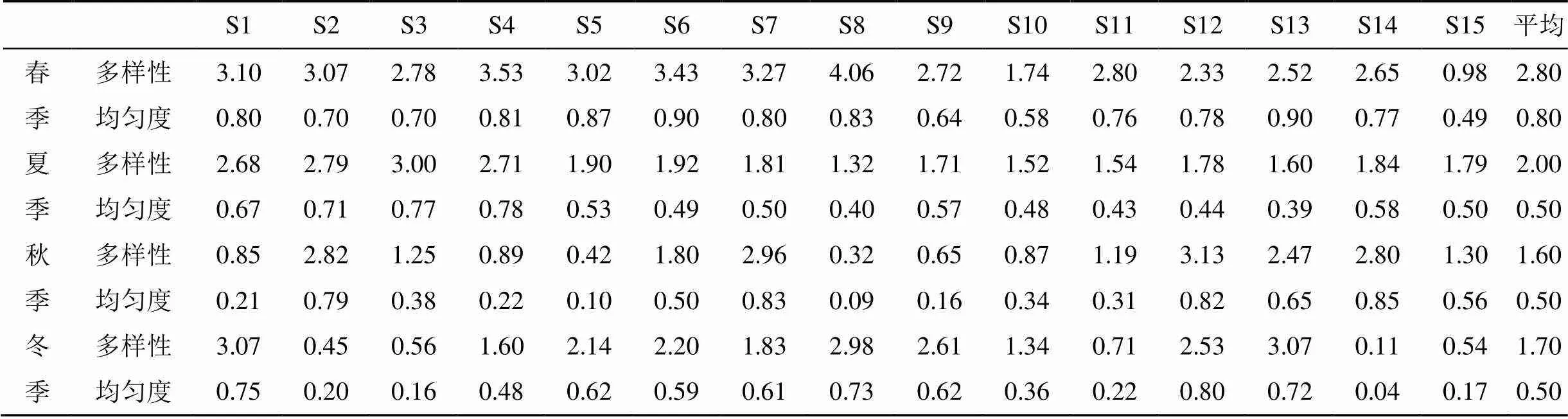
表 3 各季節浮游植物多樣性指數及均勻度

表4 浮游植物細胞豐度與環境因子的相關性
注:*.<0.05; **.<0.01。
3 討論
3.1 浮游植物群落結構
深圳灣地處亞熱帶, 水溫較高, 灣口水域寬闊, 是咸淡水的交匯處, 鹽度較低, 營養鹽較高, 適合浮游植物的生長繁殖[1]。該海域浮游植物一般以廣溫廣鹽種和沿岸性種類為主。本次調查中, 深圳灣海域共鑒定出浮游植物111種, 以硅藻門為主。這與膠州灣[16]大亞灣[17]萊州灣[18]等其他海灣情況一致。與2006年孫金水[10]的調查結果相比, 深圳灣海域硅藻門總種類數所占比例下降, 甲藻門所占比例呈現明顯上升趨勢。
優勢種在群落中起著主導作用, 決定著群落結構中的能量傳遞和物質循環的途徑和方式, 支配著群落結構的演替方向。深圳灣海域主要優勢種是中肋骨條藻、冕孢角毛藻、小型月牙藻和微小色球藻()。其中冕孢角毛藻為春季主要優勢種, 優勢度達0.12, 所占比例12.5%。大量研究表明, 春季是角毛藻的多發季節[19-21]。在春季鹽度較低且溫度適宜的環境下大量繁殖成為優勢種群。夏季優勢種有小型月牙藻、微小色球藻、小球藻和細小平裂藻。其中小型月牙藻、微小色球藻優勢度較高分別為0.426和0.370, 所占比例為42.64 %和39.65%。冬季和秋季主要優勢種為中肋骨條藻, 優勢度分別為0.840和0.565, 所占比例為83.99%和56.46%。中肋骨條藻在春秋冬三個季節均為優勢種, 這可能與深圳灣無機氮和活性磷酸鹽含量較高相關[22]。研究表明, 中肋骨條藻是中國近海常見赤潮藻類, 是富營養化海域常見的優勢種[23](表5)。
Shannon—Weaner多樣性指數通常反映群落結構的復雜程度, 多樣性指數越高, 其群落結構越復雜, 水質越好; 多樣性指數減小, 群落結構趨于簡單, 穩定性變差, 水質下降[12]。浮游植物物種多樣性和均勻度指數季節變化明顯, 分別為0.11—4.06和0.04—0.90, 深圳灣海域大部分處于中度污染的狀態, 均勻度指數差異較大, 群落結構趨于單一化, 群落穩定性較差。
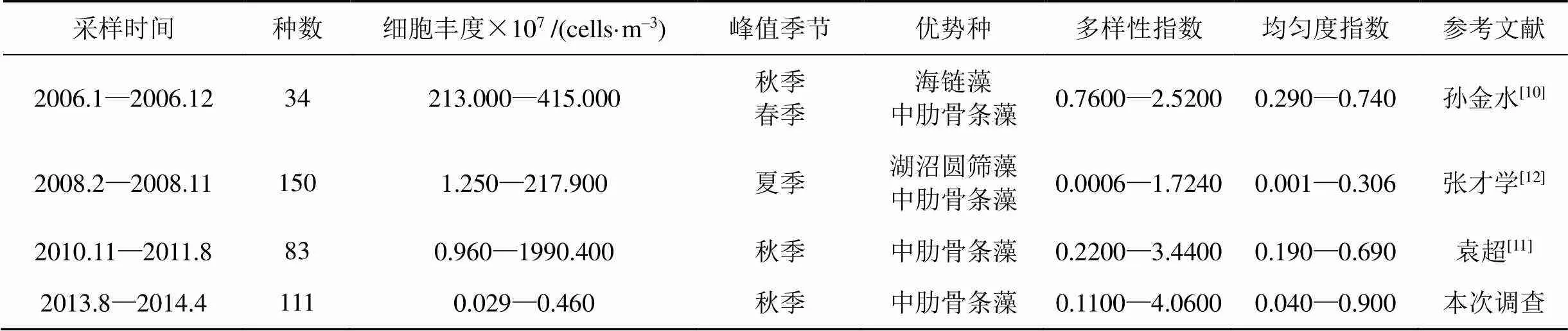
表5 深圳灣浮游植物群落結構歷史資料對比
3.2 浮游植物細胞豐度
浮游植物細胞豐度最高值和最低值分別出現在秋季和春季, 這與袁超等[11]2010—2011年對深圳灣浮游植物的研究相一致。與歷史資料相比, 本次調查中浮游植物細胞豐度明顯偏低, 這可能與深圳灣水生態系統近年來受到人類活動的破壞相關。深圳灣浮游植物細胞豐度呈現西岸(深圳一側)高于東岸(香港一側), 灣內高于灣口的趨勢。這可能與深圳灣陸源污染影響相關。人類活動所造成的大量陸源污染物進入深圳灣, 引起富營養化, 促使藻類大量生長繁殖[12]。陸源輸入是影響深圳灣海域浮游植物細胞豐度的重要因素。在一般亞熱帶海域, 浮游植物細胞豐度周年變化曲線呈現雙周期變動形式, 春季為最高峰秋季為次高峰[24]。而在本次調查中, 深圳灣浮游植物細胞豐度的季節變化呈單峰型, 秋季浮游植物的平均細胞豐度遠大于其余3個季節, 春季細胞豐度最低, 其季節變化趨勢為秋季>夏季>冬季>春季。夏秋季降水豐富, 陸源徑流較大, 給該海域帶來了大量營養物質, 在適宜的水溫和光照的作用下, 夏秋季浮游植物大量繁殖。相反, 春季是枯水期, 所需的營養物質較少, 浮游植物生長受限, 細胞豐度降低。
3.3 影響深圳灣浮游植物分布的環境因子
浮游植物的生長繁殖與海水透明度、溫度、鹽度、pH以及營養鹽含量等環境因子密不可分[25–27]。海水中無機氮和磷酸鹽等營養鹽是浮游植物生長繁殖必不可少的營養元素, TEELING等[28]研究表明, 浮游植物細胞密度與水體內氮磷含量成正相關關系, 在一定的范圍內且其他影響因子一致的情況下, 氮磷含量越高, 浮游植物細胞密度越高。對深圳灣各季節浮游植物細胞豐度與其對應的環境因子進行相關性分析發現, 在夏季深圳灣浮游植物細胞豐度與總氮呈極顯著正相關關系(<0.01), 在冬季, 深圳灣浮游植物細胞豐度與總氮和無機氮呈極顯著相關關系(<0.01), 與總磷呈顯著正相關關系(<0.05)。與珠江口的開放性水域不同, 深圳灣屬于一個半封閉的水體, 水動力條件差, 水體交換較弱[29], 且受人類活動干擾明顯。深圳一側由于經濟高速發展, 營養鹽入海負荷量一直處于較高水平, 香港一側入海通量主要來自元朗, 元朗地區人口稠密, 工商業和養殖業較發達[30]。大量的陸源輸入且與外海水域的水體交換能力較弱, 導致深圳灣中平均的氮磷含量居高不下, 無機氮和無機磷含量全年高于四類海水標準。
4 結論
(1)本次調查深圳灣浮游植物共鑒定出111種, 大部分隸屬于硅藻門, 占種類總數的77.5%, 甲藻門占13.5%, 綠藻門和藍藻門分別占5%和4%。中肋骨條藻為深圳灣海域的常年優勢種, 且優勢度較高。
(2)浮游植物細胞豐度變化范圍為28.64—462.46× 104cells·m–3范圍內, 平均為156.43×104cells·m–3。秋季的平均細胞豐度遠大于其余3個季節, 其季節變化趨勢為秋季>夏季>冬季>春季; 其空間變化趨勢為灣內高于灣口, 西側高于東側。
(3)深圳灣浮游植物物種多樣性指數和均勻度指數變化范圍分別為0.11—4.06和0.04—0.90, 多樣性指數和均勻度指數均偏低, 群落結構趨于單一化, 穩定性差。
(4)浮游植物細胞豐度與環境因子的相關性關系在不同季節有顯著差別, 其中營養鹽和鹽度與浮游植物細胞豐度之間呈現顯著的相關性關系。
[1] 王琳, 陳上群. 深圳灣自然條件特征及治理應注意的問題[J]. 人民珠江, 2001, (06): 4–7.
[2] 張瑜斌, 章潔香, 張才學, 等. 赤潮多發區深圳灣葉綠素a的時空分布及其影響因素[J]. 生態環境學報, 2009, 018(5): 1638–1643.
[3] 冷科明, 江天久. 深圳海域近20年赤潮發生的特征分析[J]. 生態科學, 2004, 23(2): 166–170.
[4] 陳淑文, 宋俊廷, 類成龍, 等. 深圳灣浮游細菌生物量的時空動態及影響因素[J]. 海洋學研究, 2016, 34(4): 84– 91.
[5] PADISáK J, CROSSETTI L O, Naselli-Flores L. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates[J]. Hydrobio-logia, 2009, 621(1): 1–19.
[6] RODRIGUES L, SIM?ES N, BOVO-SCOMPARIN V, et al. Phytoplankton alpha diversity as an indicator of environmental changes in a neotropical floodplain[J]. Ecological Indicators, 2015, 48: 334–341.
[7] CUNLIFFE M, SALTER M, MANN P, et al. Dissolved organic carbon and bacterial populations in the gelatinous surface microlayer of a Norwegian fjord mesocosm[J]. FEMS Microbiology Letters, 2009, 299(2): 248–254.
[8] 桓清柳, 龐仁松, 周秋伶, 等. 深圳近岸海域氮、磷營養鹽變化趨勢及其與赤潮發生的關系[J]. 海洋環境科學, 2016, 035(6): 908–914.
[9] 王朝暉, 齊雨藻, 尹伊偉, 等. 1998年春深圳灣環節環溝藻赤潮及其發生原因的探討[J]. 海洋科學, 2001, 25(05): 48–51.
[10] 孫金水, WING-HONG WO, 戴紀翠, 等. 深圳灣海域浮游植物的生態特征[J]. 環境科學研究, 2010, 31(1): 63– 68.
[11] 袁超, 徐宗軍, 張學雷. 2010-2011年深圳灣浮游植物季節變化及其與環境因子關系[J]. 海洋湖沼通報, 2015, (1): 114–122.
[12] 張才學, 周凱, 孫省利, 等. 深圳灣浮游植物的季節變化[J]. 生態環境學報, 2010, 19(10): 2445–2451.
[13] 中華人民共和國國家標準. 海洋監測規范, 第7部分:近海污染生態調查和生物監測[S]. 北京:中國標準出版社, 2007: 84-85.
[14] PIELOU E C. Species-diversity and pattern-diversity in the study of ecological succession[J]. Journal of theoretical biology, 1966, 10: 370–83.
[15] 馬建新, 鄭振虎. 萊州灣浮游植物分布特征[J]. 海洋湖沼通報, 2002(4): 63–67.
[16] 楊世民, 王麗莎, 石曉勇. 2009年春季膠州灣浮游植物群落結構特征[J]. 海洋與湖沼, 2014, 45(6): 1234–1240.
[17] 魏雷. 大亞灣海域浮游植物群落結構調查—以2011—2012年為例[J]. 農業與技術, 2017, 37(16): 243–244.
[18] 張曉舉, 趙升. 萊州灣西部浮游植物群落與環境因子關系的研究[J]. 大連海洋大學學報, 2018, 33(04): 122–128.
[19] 張冬鵬, 黎曉濤, 黃遠峰, 等. 深圳沿海浮游植物組成及赤潮發生趨勢分析[J]. 暨南大學學報:自然科學與醫學版, 2001, 022(5): 122–126.
[20] 張雪, 徐曉甫, 戴媛媛, 等. 天津近岸人工魚礁海域浮游植物群落及其變化特征[J]. 漁業科學進展, 2018, 039(6): 1–10.
[21] BODE A, áLVAREZ-OSSORIO M T, GONZáLEZ N, et al. Seasonal variability of plankton blooms in the Ria de Ferrol (NW Spain): II. Plankton abundance, composition and biomass[J]. Estuarine Coastal and Shelf Science, 2005, 63(1–2): 285–300.
[22] 張靜, 張瑜斌, 周凱, 等. 深圳灣海域營養鹽的時空分布及潛在性富營養化程度評價[J]. 生態環境學報, 2020, 19(2): 7–15.
[23]霍文毅, 俞志明, 鄒景忠, 等. 膠州灣中肋骨條藻赤潮與環境因子的關系[J]. 海洋與湖沼, 2001, 32(3): 311–318.
[24] 羅璇, 孫曉霞, 鄭珊, 等. 2011年膠州灣網采浮游植物群落結構及其環境影響因子[J]. 海洋與湖沼, 2016, 47(5): 915–923.
[25] HAN Boping, XIAO Lijuan, HU Ren, et al. Succession of phytoplankton functional groups regulated by monsoonal hydrology in a large canyon-shaped reservoir[J]. Water research: A journal of the international water association, 2011, 45(16): 5099-5109.
[26] BERGE T, DAUGBJERG N, Andersen B, et al. Effect of lowered pH on marine phytoplankton growth rates[J]. Marine Ecology Progress Series, 2010, 416(12): 79–91.
[27] MUHID P, DAVIS T W, BUNN S E, et al. Effects of inorganic nutrients in recycled water on freshwater phytoplankton biomass and composition[J]. Water Research, 2013, 47(1): 384–394.
[28] TEELING H, FUCHS B, BECHER D, et al. Substrate- Controlled Succession of Marine Bacterioplankton Populations Induced by a Phytoplankton Bloom[J]. Science (New York, N. Y. ), 2012, 336(6081): 608–611.
[29] HE B, DAI M, ZHAI W, et al. Distribution, degradation and dynamics of dissolved organic carbon and its major compound classes in the Pearl River estuary, China[J]. Marine Chemistry, 2010, 119(1–4): 52–64.
[30] 趙晨辰, 張世彥, 毛獻忠. 深圳灣流域TN和TP入海年通量變化規律研究[J]. 環境科學研究, 2014, 35(11): 4111–4117.
Community structure of phytoplankton and its relationship to environmental factors in Shenzhen Bay
CHEN Si1, 2, 3, CHEN Haigang2, TIAN Fei2, LI Yitong2, ZHANG Linbao2, ZHANG Zhe2, WANG Xuefeng4, CAI Wengui2, *
1. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China 2. South China Fisheries Research Institute, Guangdong Provincial Key Laboratory of Fishery Ecology and Environment, Scientific Observation and Research Field Station of Zhujiang Estuary Ecosystem, Guangdong Province, Key Field Scientific Experimental Station of South China Fishery Resource and Environment, Ministryof Agriculture, Guangzhou 510300, China 3. Guangzhou Peng Jia Mu Memorial Secondary School, Guangzhou 510430, China 4. Southern Marine Science and Engineering Guangdong Laboratory(Zhangjiang), Zhanjiang 524025, China
In order to investigate the phytoplankton community structure and its relationship with the aquatic environmental factors in the Shenzhen bay, samples were collected and analyzed in four quarters from August 2013 to April 2014, and the correlation between phytoplankton and its main environmental factors was analyzed. Results showed that a total of 111 species (including varieties and forma) were identified, of which the most abundant group was, with 35 genera and 86 species that accounted for 77.5% of total species, followed by(4 genera and 15 species, 13.5%),(6 genera and 6 species, 5%) and(3 genera 4 species, 4%).was the dominant species. The abundance of phytoplankton cells varied from (28.64-462.46)×104cells·m–3, with an average of 156.43×104cells·m–3. The maximum cell abundance appeared in the autumn, followed in summer, spring and winter The diversity index of the phytoplankton ranged from 0.11 to 4.06; the evenness of phytoplankton ranged from 0.04 to 0.90.The diversity and evenness of phytoplankton community were rather low, indicating that phytoplankton community was not steady. Better relationship was found between phytoplankton abundance and inorganic nitrogen.
Shenzhen Bay; phytoplankton; community structure; environmental factors
陳思, 陳海剛, 田斐, 等. 深圳灣浮游植物群落結構特征及其與環境因子的關系[J]. 生態科學, 2021, 40(1): 9–16.
CHEN Si, CHEN Haigang, TIAN Fei, et al. Community structure of phytoplankton and its relationship to environmental factors in Shenzhen Bay[J]. Ecological Science, 2021, 40(1): 9–16.
10.14108/j.cnki.1008-8873.2021.01.002
Q178
A
1008-8873(2021)01-009-08
2020-03-03;
2021-03-08
南方海洋科學與工程廣東省實驗室(湛江)資助項目( ZJW-2019-06) ; 中國水產科學研究院南海水產研究所中央級公益性科研院所基本科研業務費專項資金項目(2021SD17); 廣東省科技計劃項目(2019B121201001)
陳思(1994—), 女, 黑龍江雙鴨山人, 碩士研究生,主要從事海洋生態學研究, E-mail: xiaochensi2@163.com
蔡文貴(1965—), 男, 研究員, 主要從事漁業生態環境方面研究, E-mail: cai-wengui@163.com
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
趣味(語文)(2020年3期)2020-07-27 01:42:46
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
