熒光假單胞菌的溶磷機制及其在楊樹菌根際的定殖動態*
2021-04-10 04:07:08吳小芹葉建仁
林業科學 2021年3期
劉 輝 吳小芹 葉建仁 陳 丹
1.南京林業大學林學院 南方現代林業協同創新中心 南京 210037; 2.安徽師范大學生態與環境學院 蕪湖 241002)
土壤磷素存量大,但其中約95%不能被植物直接吸收利用,溶磷微生物(phosphate-solubilizing microorganisms, PSM)是一類參與土壤磷循環的重要微生物類群,其可將土壤磷庫中的難溶性磷酸鹽轉化為可溶性磷,從而顯著提高土壤有效磷含量,促進植物生長和發育(Kuceyetal., 1989; 陳哲等, 2009)。目前,學者們已對 PSM 的種類、溶磷能力和應用效果等開展了廣泛而深入的研究(Oliveriaetal., 2009; Wangetal., 2017; Chenetal., 2019)。有研究發現,許多在離體條件下表現出明顯溶磷能力的 PSM 在實際應用中效果并不理想(Collavinoetal., 2010; 張英等, 2015)。影響 PSM 應用效果的因素很多,分析其原因主要有2方面: 一方面是由于 PSM 種類較多,溶磷過程十分復雜,其潛在機制也因 PSM 種類和菌株不同而有所區別(Antoun, 2012),當前對 PSM 溶磷機制認識的局限性制約了該類菌肥的研發和推廣應用進程; 另一方面是由于磷在土壤中的移動性較小,PSM 釋放到土壤中的磷必須在植物可以吸收的范圍內(根際)才能被利用(朱培淼等, 2007)。Gupta 等(2011)研究發現,產氣腸桿菌(Enterobacteraerogenes)10208 的溶磷能力是唐菖蒲伯克氏菌(Burkholderiagladioli)10216 的 13.8 倍,但對甜葉菊(Steviarebaudiana)促生效應和地上部磷吸收量之間的差異并不顯著,分析其原因發現可能是 10216 菌株在甜葉菊根際定殖數量比 10208 菌株高1個數量級造成的。可見,PSM 能否在植物根際成功定殖也是影響其應用效果的關鍵因素(Mamtaetal., 2010)。在自然和半自然生態系統中,植物的根部常常被菌根真菌侵染,根際的概念已擴大到包括菌根真菌部分而被稱作菌根際(Mycorrhizosphere)(Johanssonetal., 2004)。據報道,PSM 和菌根真菌在磷素資源可持續利用方面具有重要作用,但只有PSM 在植物根際或菌根際高效定殖才能達到更好為宿主植物提供有效磷的目的(Antoun, 2012)。因此,在評價 PSM 的應用潛力時,其與菌根真菌的相容性問題不容忽視。
筆者課題組前期研究發現,熒光假單胞菌(Pseudomonasfluorescens)菌株JW-JS1和紅絨蓋牛肝菌(Xerocomuschrysenteron, Xc)共接種對NL-895楊(Populus×euramericanacv. ‘Nanlin-895’)的生長、光合效應、氮代謝、礦質元素含量、土壤酶活性和微生物多樣性等的影響均表現出顯著的正交互效應(姚如斌等, 2012; 劉輝等, 2018; 2019),但關于JW-JS1菌株的溶磷機制及其在楊樹根際和菌根際的定殖動態尚不明確。鑒于此,本研究以熒光假單胞菌菌株JW-JS1和紅絨蓋牛肝菌 Xc 為研究對象,通過分析JW-JS1菌株溶磷能力與pH、可滴定酸含量以及有機酸種類和含量的關系,初步揭示其溶磷機制, 利用抗利福平標記篩選穩定的標記菌株,采用灌根法對其在楊樹根際和菌根際的定殖動態進行深入研究,進一步揭示溶磷細菌與外生菌根真菌的互作機制,以期為楊樹專用復合菌劑的開發與應用提供理論依據。
1 材料與方法
1.1 試驗材料
1.1.1 供試菌株 溶磷細菌: 熒光假單胞菌菌株JW-JS1,從楊樹根際土壤中分離篩選獲得(Liuetal., 2011),保藏于中國典型培養物保藏中心,編號CCTCC M 209027; 外生菌根真菌: 紅絨蓋牛肝菌,保存于南京林業大學森林病理實驗室。
1.1.2 供試試劑與培養基 利福平(Rifampicin)購自美國 Sigma 公司; 磷酸鹽生長培養基(NBRIP): 葡萄糖 10.0 g,MgCl25.0 g,MgSO4·7H2O 0.25 g,KCl 0.2 g,(NH4)2SO40.1 g,Ca3(PO4)25.0 g,蒸餾水1 000 mL,pH 7.0; 細菌活化培養基(NA): 牛肉膏 3.0 g,蛋白胨 10.0 g,NaCl 5.0 g,瓊脂 15.0~20.0 g,pH7.2~7.4,蒸餾水1 000 mL; 細菌發酵培養基(NB): 牛肉膏 3.0 g,蛋白胨 10.0 g,NaCl 5.0 g,蒸餾水 1 000 mL,pH7.2~7.4。以上培養基均于 121 ℃條件下滅菌 20 min,備用。
1.1.3 供試植物材料 采用0.5% 高錳酸鉀水溶液浸泡美洲黑楊(Populusdeltoides)種子(采自南京林業大學實驗基地)10 min,殺菌處理后播種于分別盛有滅菌盆栽基質和含有0.5% 紅絨蓋牛肝菌 Xc 固體菌劑(曾麗瓊, 2010)的滅菌盆栽基質的花盆中,置于日光溫室培養,出苗2周后按照每盆中央1株、周圍5株的方式保留6株實生苗,定期澆水保濕,培養 3 個月后備用。花盆規格為12 cm×9 cm×12 cm(上口徑×下口徑×高),裝干盆栽基質每盆質量0.6 kg; 盆栽基質為土壤(pH7.56,有機質2.65 g·kg-1、采自南京林業大學校園后山)、砂子、蛭石按2∶1∶1比例混合并攪拌均勻在1.01×106Pa 壓力下滅菌120 min 后備用。 經檢測,接種Xc的美洲黑楊實生苗菌根侵染率約為 70%。
1.2 試驗方法
1.2.1 JW-JS1菌株溶磷能力測定 將供試菌株活化后接種于 NB 培養基中,28 ℃、180 r·min-1振蕩培養 18~24 h 制成種子液,菌數約 109cfu·mL-1。按 1% 接種量接入 NBRIP 培養基,以不接菌處理為對照,30 ℃、180 r·min-1振蕩培養。每隔 24 h 取 3 個重復,培養液離心 20 min(4 ℃、10 000 r·min-1),上清液采用鉬銻抗比色法測定可溶性磷含量(扣除對照后的值)(劉輝等, 2013),滴定法測定可滴定酸含量(劉輝等, 2010), pH 計(上海雷磁 pHS-3C 型號)測定上清液pH 變化。
1.2.2 JW-JS1菌株分泌有機酸的種類及含量測定 按1.2.1方法制備JW-JS1菌株的種子液,按1% 接種量接入NBRIP 培養基,以不接菌處理為對照,30 ℃、180 r·min-1振蕩培養 72 h 后,培養液離心 20 min(4 ℃、10 000 r·min-1)。取 5 mL 培養液上清液過 0.22 μm 孔徑濾膜,采用高效液相色譜儀(HPLC)(Agilent 1100 series,美國產)測定有機酸種類和含量。色譜條件: 色譜柱為菲羅門 C18柱,緩沖液為 0.05 mol·L-1磷酸二氫鉀(pH 2~3),柱溫 30 ℃,流速 1.0 mL·min-1,檢測波長 214 nm。有機酸標準樣品: 乙酸、草酸、酒石酸、檸檬酸、順丁烯二酸、反丁烯二酸、丁二酸和乳酸。
1.2.3 抗利福平標記菌株的篩選及穩定性分析 采用抗利福平標記法(王長方等, 2006)篩選JW-JS1菌株的標記菌株,直至所篩選菌株在含有 300 μg·mL-1利福平的 NA 平板上能穩定生長,菌落形態與原始菌株保持一致,菌株編號為JW-JS1Rif。按1.2.1方法,測定 JW-JS1和JW-JS1Rif菌株的溶磷能力。
1.2.4 JW-JS1Rif在楊樹根際和菌根際的定殖動態測定 將標記菌株JW-JS1Rif活化后接種于 NB 培養基中,28 ℃、180 r·min-1振蕩培養 36 h制成接種劑,菌數約109cfu·mL-1。采用灌根法將標記菌株接種劑均勻澆灌于各植株周圍土壤,接種量為每盆50 mL,以清水為對照(CK),每處理3次重復(3盆)。參照居正英(2008)菌株分離回收方法,分別于接種后第 1、2、3、4、5、7、9、11、16、20、25、30、40 和 50 天取楊樹處理組和對照組進行目標菌株分離回收。使用抖土法收集楊樹根際和菌根際的土壤樣品,采用平板稀釋法計數菌落,具體方法如下: 稱取楊樹根際和菌根際待測土樣各1.0 g,加入9.0 mL無菌水進行梯度稀釋,選取10-2、10-3、10-4土壤梯度稀釋液各100 μL均勻涂布于 NA 培養基平板上(含300 μg·mL-1利福平),28 ℃培養48 h后,分別計數每皿菌落數,并換算成每克干土中的菌落數(cfu·g-1)。每處理 3次重復。
1.3 數據分析
采用Excel 2016和SPSS 22.0軟件進行數據統計分析并制圖,利用單因素(One-way ANOVA)和Duncan法進行方差分析和多重比較(α= 0.05)。圖表中數據為平均值 ± 標準差。
2 結果與分析
2.1 JW-JS1菌株的溶磷能力
JW-JS1 菌株在 NBRIP 培養基中培養后培養液可溶性磷含量隨接種時間延長逐漸增加。由圖 1 可知,可溶性磷含量在培養 1~3 天時增速較大,3 天后基本達到平衡狀態,增速明顯變緩(4~7 天),7 天后含量達 616.39 mg·L-1。培養液 pH 在培養1天時迅速下降,從初始的 7.0 降至 3.73,隨后(2~7 天)基本維持在這一水平(圖 1)。相關分析表明,JW-JS1 菌株培養液 pH(X)與可溶性磷含量(Y)呈極顯著負相關(r=-0.889**)。
培養液可滴定酸含量的變化趨勢與可溶性磷基本一致。由圖2可知,可滴定酸含量在培養1天時迅速增加,隨后增速變緩(2~7 天),7 天后含量為 56.30 mmol·L-1。相關分析表明,JW-JS1 菌株培養液可滴定酸含量(X)與可溶性磷含量(Y)呈極顯著正相關(r= 0.958**)。進一步分析JW-JS1 菌株培養液 pH 與可滴定酸含量的關系發現,二者呈極顯著負相關(r=- 0.980**)。
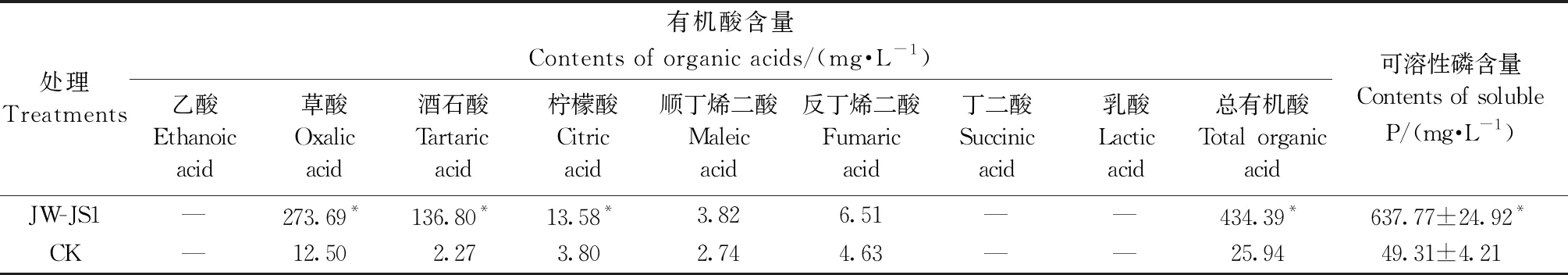
表1 熒光假單胞菌菌株JW-JS1分泌有機酸種類和含量及可溶性磷含量①
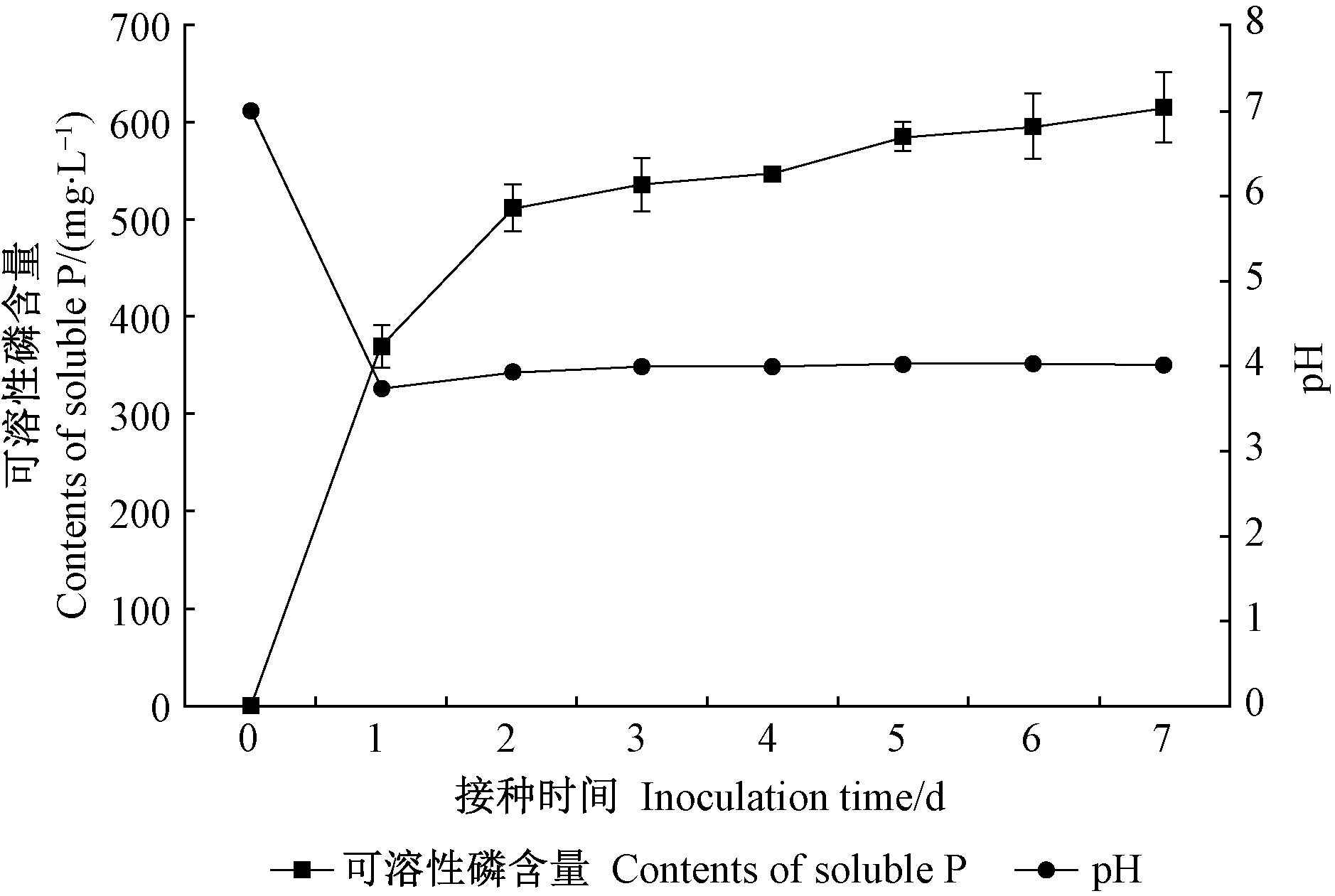
圖1 熒光假單胞菌菌株JW-JS1的溶磷能力與pH的關系
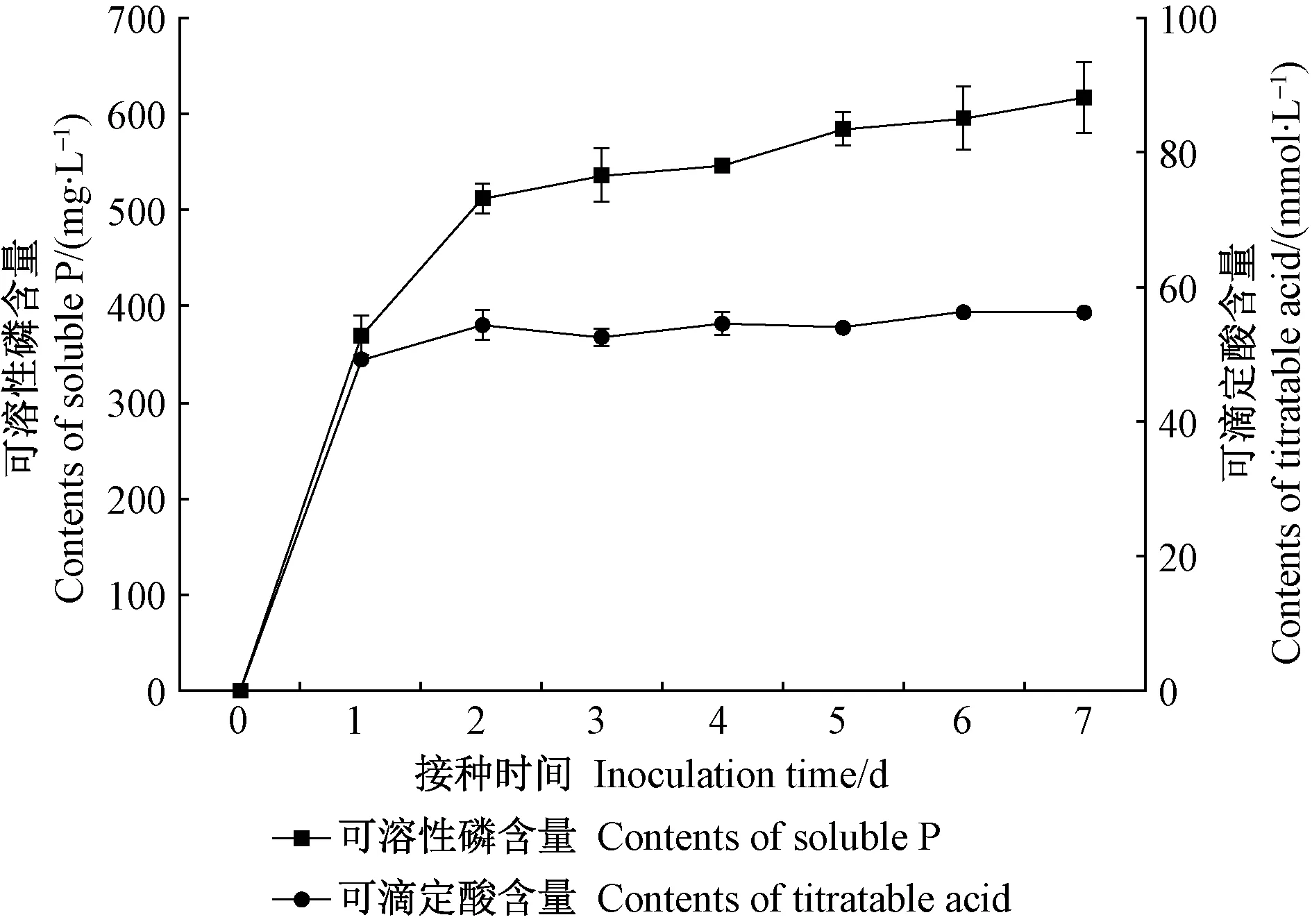
圖2 熒光假單胞菌菌株JW-JS1的溶磷能力與可滴定酸含量的關系
2.2 JW-JS1菌株分泌有機酸的種類和含量
HPLC分析表明,JW-JS1 菌株在 NBRIP 培養基中培養 72 h 后能夠分泌多種有機酸。由表1可知,JW-JS1 菌株分泌的總有機酸量(434.39 mg·L-1)明顯高于 CK(25.94 mg·L-1),是 CK 的 16.74 倍。在培養液中共檢測出為草酸、酒石酸、檸檬酸、順丁烯二酸和反丁烯二酸 5 種有機酸,其中,草酸含量(273.69 mg·L-1)明顯高于其他種類有機酸,約占總有機酸量的 63.01%,這說明 JW-JS1 菌株分泌的草酸可能在溶磷過程中發揮重要作用。
2.3 JW-JS1菌株在楊樹菌根際的定殖動態
2.3.1 抗利福平標記菌株的篩選及穩定性 通過對JW-JS1菌株進行抗利福平標記,篩選得到抗利福平(300 μg·mL-1)突變菌株JW-JS1Rif。標記菌株在不含利福平的NA平板上連續傳代10次,轉接于含300 μg·mL-1利福平的NA平板上生長良好,菌落形態和顏色等與原始菌株基本相同,即 JW-JS1Rif菌株與JW-JS1菌株的菌落均為乳黃色,圓形菌落小,濕潤,邊緣整齊,說明突變菌株抗性標記穩定。由表2可知,原始JW-JS1菌株培養液可溶性磷含量為584.17 mg·L-1,在NBRIP培養基中培養 72 h 后,標記菌株JW-JS1Rif培養液可溶性磷含量為579.26 mg·L-1,二者差異不顯著(P<0.05),這說明標記菌株JW-JS1Rif的溶磷能力基本未退化,可作為溶磷細菌的靶標細菌,適合開展定殖試驗研究。
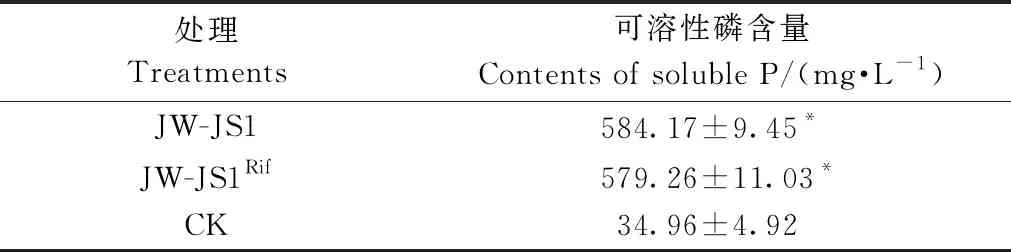
表2 JW-JS1Rif 菌株與 JW-JS1 菌株溶磷能力比較
2.3.2 JW-JS1Rif菌株在楊樹根際和菌根際的定殖動態 采用灌根法接種后,在楊樹根際和菌根際中均回收到標記菌株JW-JS1Rif,而無菌水對照組在含300 μg·mL-1利福平的NA平板上未見細菌菌落生長。灌根接種處理后,標記菌株在楊樹根際和菌根際均能長期穩定存活并保持一定的定殖數量,隨接種時間延長定殖數量呈下降趨勢,定殖動態基本一致(圖3)。接種第 1 天時JW-JS1Rif菌株在楊樹根際和菌根際的定殖數量最大,分別為2.34 × 106和1.34 × 106cfu·g-1,第 2 天時急劇下降,之后趨于平緩,進入穩定期。接種后第1~9 天JW-JS1Rif菌株在楊樹根際的定殖數量高于菌根際,隨著接種時間延長,第9~50 天定殖數量基本保持一致,接種50 天后定殖數量分別為 5.2×104和4.5×104cfu·g-1。
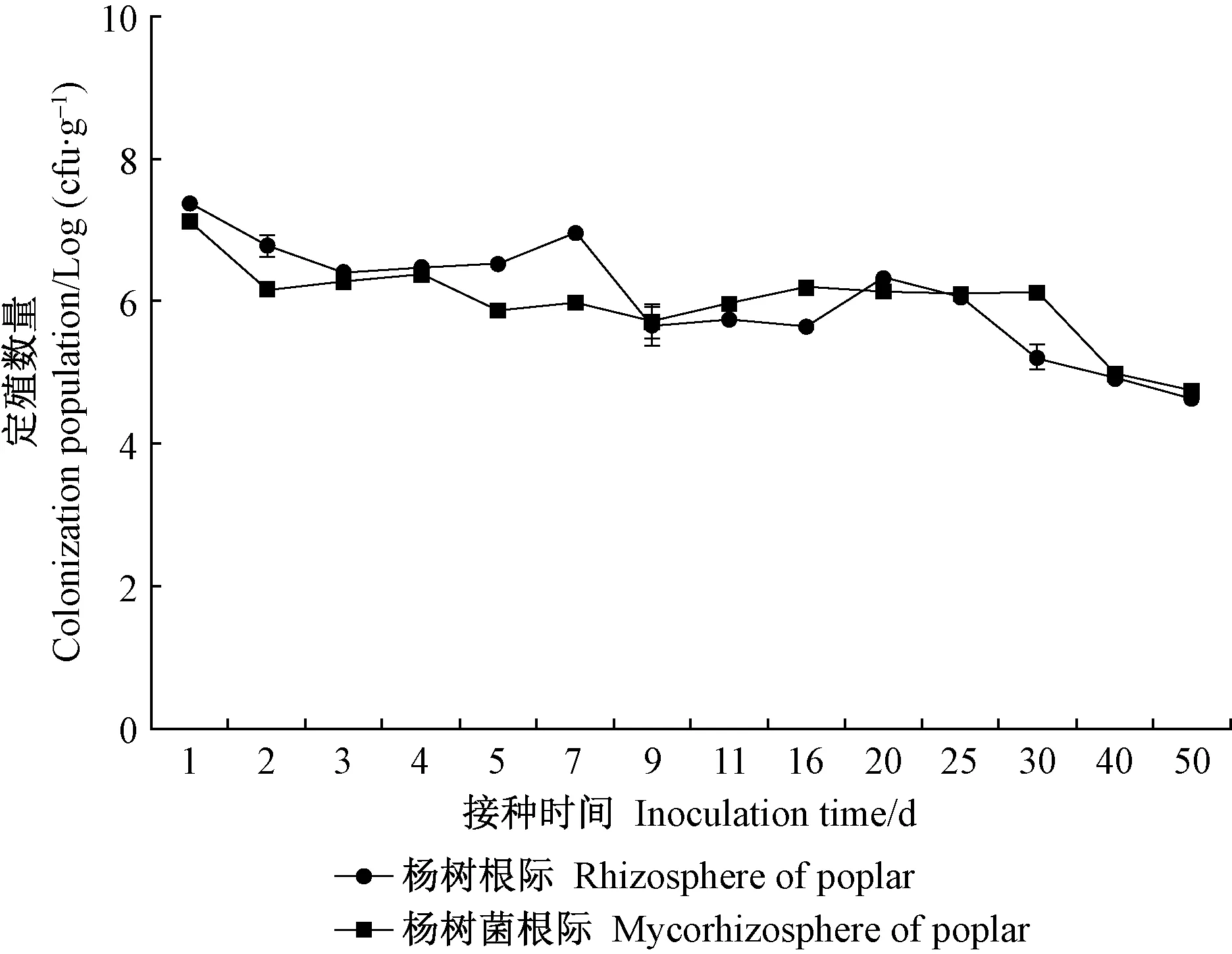
圖3 JW-JS1Rif菌株在楊樹根際和菌根際的定殖動態
3 討論
不同種類溶磷微生物的溶磷過程存在較大差異,溶磷機制也不盡相同。有研究發現,培養介質 pH 降低是溶磷的重要條件而并不是必要條件(Kucey, 1988),但近年來的研究認為培養介質 pH 與溶磷量之間存在一定的負相關(Seshadrietal., 2004)。本研究 JW-JS1 菌株在溶磷過程中,隨著 pH 快速下降,培養液可溶性磷含量迅速上升,當 pH 降到一定值時可溶性磷基本達到平衡狀態,其含量雖有少量增加,但增速明顯變緩,與熒光假單胞菌RAF15 菌株的溶磷規律基本一致(Parketal., 2009)。分析其其原因可能是: 隨著接種時間延長,培養液中碳源、氮源等營養物質逐漸被消耗,一定時間后微生物生長進入穩定期,可溶性磷含量不再增加。相關分析發現,JW-JS1 菌株的溶磷能力與培養液 pH 呈極顯著負相關(r=-0.889**),與趙小蓉(2002)、Antoun(2012)和張英等(2015)研究結論基本一致。可滴定酸含量可間接反映有機酸的變化情況(Tyletal., 2017),JW-JS1菌株的溶磷能力與可滴定酸含量呈極顯著正相關(r= 0.958**),與劉輝等(2010)和 Tyl 等(2017)研究結論基本一致。進一步分析發現,JW-JS1菌株培養液pH與可滴定酸含量呈極顯著負相關(r=- 0.980**),這說明培養液 pH 降低和可滴定酸含量增加可能是由于JW-JS1 菌株分泌有機酸引起的。
分泌有機酸是微生物溶解無機磷的主要機制之一,分泌的酸既可降低培養介質 pH,又可通過羥基或羧基與 Ca2+、Fe3+、AL3+等金屬陽離子螯合,使難溶性磷轉化為可溶性磷(Khanetal., 2007)。目前,已報道的 PSM 產生的有機酸多種多樣,主要包括草酸、乳酸、蘋果酸、琥珀酸、酒石酸、檸檬酸、丁二酸、乙酸和葡萄糖酸等(Chenetal., 2006; Songetal., 2008)。本研究通過 HPLC 檢測發現, JW-JS1 菌株在溶磷過程中能夠分泌草酸、酒石酸、檸檬酸、順丁烯二酸和反丁烯二酸。不同微生物在溶解不溶性磷酸鹽時,產生的有機酸種類和數量有很大不同,特定的溶磷微生物在溶磷過程中分泌的有機酸種類和數量也在變化(Vyasetal., 2009),與本研究結果相一致。草酸屬共軛酸,草酸根遇到 Ca2+會形成草酸鈣沉淀,從而使與 Ca2+結合的磷酸根得以釋放,對提高難溶性磷酸鈣鹽的溶解性起到重要作用(王樹起等, 2009)。本研究發現,在檢測出的5種有機酸中,草酸分泌量最大,其含量明顯高于其他種類有機酸,這說明 JW-JS1 菌株分泌的草酸可能在溶磷過程中發揮重要作用,與 Li 等(2016)、馮哲葉等(2017)研究論相似。近年來,隨著分子生物學的發展,有關溶磷基因的報道很多,如pqq基因(焦子偉等, 2016)、gab-Y基因(Babu-khanetal., 1995)、GDH基因(楊美英等, 2016)等;gcd基因(Sulemanetal., 2018)的成功克隆不僅說明了溶磷基因的豐富性,而且也佐證了溶磷機制的復雜性。因此,今后應采用分子生物學技術深入揭示溶磷相關基因的種類和功能,從而更深入地闡明熒光假單胞菌菌株JW-JS1的溶磷機制。
植物根際有益微生物一直面臨著應用效果不穩定的問題,而造成不穩定的因素主要是有益微生物在植物根際的定殖能力存在差別(孫真等, 2017)。李阜棣(1996)研究認為,外源微生物介入土壤必須經過與土著微生物生態位的競爭(包括營養、空間等),才能在根際定殖并發揮作用。王恒煦等(2019)利用利福平標記3株芽孢桿菌(Bacillussp.),明確了其在水稻(Oryzasativa)根際的定殖能力。本研究利用利福平標記篩選出含 300 μg·mL-1利福平抗性的菌株JW-JS1Rif,灌根法定殖檢測發現,JW-JS1Rif菌株在楊樹根際和菌根際均能穩定存活并保持一定的定殖數量,隨接種時間延長定殖數量呈下降趨勢,定殖動態基本一致,分析其原因可能是標記菌株在楊樹根際和菌根際定殖需要一個適應過程,定殖數量在早期迅速下降,但經過一段時間適應后逐漸趨于穩定,與Theoduloz等(2003)和王恒煦等(2019)研究結論一致。接種后第1~9天JW-JS1Rif菌株在楊樹根際的定殖數量略高于菌根際,這可能是因為楊樹根際環境不存在外生菌根真菌的競爭,更適于其繁殖,而楊樹菌根際由于紅絨蓋牛肝菌Xc 的存在,可能對JW-JS1Rif菌株在營養、生態位等方面產生一定的競爭性,從而導致其定殖數量低于根際,但隨著接種時間延長,二者定殖數量基本維持在同一水平,這也說明該菌株與Xc 生物兼容性較好,能夠在同一生態位上形成穩定的功能菌群,在活化土壤磷營養和促進楊樹生長上發揮協同互作作用,與姚如斌等(2012)、劉輝等(2018; 2019)研究結論一致。單一菌種向多種功能菌種的組合應用是提高微生物肥料效果的有效途徑(楊順等, 2018),本研究結果表明熒光假單胞菌菌株JW-JS1 與紅絨蓋牛肝菌 Xc 是構建楊樹專用功能復合微生物肥料的理想材料。
4 結論
熒光假單胞菌菌株 JW-JS1 的溶磷能力與培養液 pH 呈極顯著負相關、與可滴定酸含量呈極顯著正相關,培養液 pH降低和可滴定酸含量增加可能是由于JW-JS1菌株分泌有機酸引起的。JW-JS1菌株在溶磷過程中能夠分泌草酸、酒石酸、檸檬酸、順丁烯二酸和反丁烯二酸,其中草酸可能在溶磷過程中發揮重要作用。JW-JS1 菌株在楊樹根際和菌根際均能長期穩定存活并保持一定的定殖數量,定殖動態基本一致,熒光假單胞菌菌株JW-JS1與紅絨蓋牛肝菌 Xc 生物兼容性較好,能夠在同一生態位形成穩定的功能菌群,具有開發成楊樹專用功能復合微生物肥料的潛力。
