滇中亞高山不同森林土壤酶活性對(duì)模擬N 沉降的響應(yīng)
2021-04-10 05:22:32鄭興蕊王克勤宋婭麗張轉(zhuǎn)敏張雨鑒
林業(yè)科學(xué)研究 2021年2期
鄭興蕊,王克勤,宋婭麗,張轉(zhuǎn)敏,張雨鑒,潘 禹
(西南林業(yè)大學(xué)生態(tài)與環(huán)境學(xué)院,云南 昆明 650224)
化石燃料與化學(xué)N 肥大量施用引起的N 沉降現(xiàn)象已成為不容忽視的全球變化問(wèn)題之一[1]。N 沉降的急劇增加,直接威脅著水域生態(tài)系統(tǒng)和陸地生態(tài)系統(tǒng)的平衡[2],而森林生態(tài)系統(tǒng)作為陸地生態(tài)系統(tǒng)的主體,以其復(fù)雜的組成與結(jié)構(gòu),不可避免地受全球氣候變化的影響。森林土壤作為森林生態(tài)系統(tǒng)的主要C 庫(kù)、N 庫(kù)[3],約將N 沉降中2/3 固定在土壤有機(jī)質(zhì)中[4]。其中,土壤酶作為森林土壤生態(tài)系統(tǒng)的組成部分,其活性高低在很大程度上反映了物質(zhì)循環(huán)的快慢[5],并在一定程度上反映了森林土壤狀況,包括有機(jī)質(zhì)的分解能力和簡(jiǎn)單物質(zhì)的再合成能力[6]。為研究N 添加對(duì)土壤酶活性的影響,可將其分為兩大類:(1) 負(fù)責(zé)獲得C、N、P 以維持初級(jí)代謝的水解酶,包括蔗糖酶、脲酶、酸性磷酸酶、β-葡萄糖苷酶等;(2) 負(fù)責(zé)降解復(fù)雜化合物以提供營(yíng)養(yǎng)物質(zhì)的氧化還原酶,包括過(guò)氧化氫酶、過(guò)氧化物酶和酚氧化酶等[7]。這些土壤酶可能對(duì)N 添加有不同的反應(yīng),而研究更多種類的土壤酶有助于我們更全面的了解N 添加后土壤有機(jī)質(zhì)的周轉(zhuǎn)過(guò)程。
N 沉降對(duì)不同森林生態(tài)系統(tǒng)下土壤酶活性存在促進(jìn)、抑制和無(wú)影響3 種影響,主要原因在于森林中的土壤理化性質(zhì)受氣候、林地類型、林齡及模擬N 沉降時(shí)間長(zhǎng)短等多個(gè)因素共同作用[8-10]。目前關(guān)于N 沉降下土壤酶活性的研究,土壤酶的種類較少,且多集中在同一林型[11],如張藝等[12]對(duì)北京市遼東櫟(Quercus mongolicaFisch.ex Ledeb.)施N 后,N 沉降顯著促進(jìn)了土壤脲酶、酸性磷酸酶活性,對(duì)土壤過(guò)氧化氫酶和多酚氧化酶活性無(wú)顯著差異;李吉玫等[13]對(duì)天山云杉林(Picea schrenkianaFisch.et Mey.)進(jìn)行模擬施N,結(jié)果表明N 沉降顯著抑制了過(guò)氧化物酶和多酚氧化酶活性,而對(duì)土壤脲酶活性有較小促進(jìn)作用;Cusack 等[7]對(duì)實(shí)驗(yàn)森林的研究表明,N 沉降可提高熱帶山地森林土壤多酚氧化酶和過(guò)氧化物酶活性。但不同土壤酶活性對(duì)外源N 添加如何響應(yīng),哪種酶活性對(duì)不同施N 濃度的響應(yīng)最為敏感仍具有不確定性。
滇中亞高山區(qū)域位于我國(guó)云貴高原、青藏高原和橫斷山地三大自然地理區(qū)域相結(jié)合的部位,植被以半濕性常綠闊葉林為主[14]。本研究選取國(guó)內(nèi)外常用于研究模擬N 沉降下土壤活動(dòng)的8 種代表性土壤酶[11-13,15-18],以該地區(qū)不同森林生態(tài)系統(tǒng)云南松(Pinus yunnanensisFranch.) 林、高山櫟(Quercus semecarpifoliaSmith) 林、華山松(Pinus armandiiFranch.)林和常綠闊葉(Evergreen broad-leaf)林為研究對(duì)象,分析林下不同土壤酶活性對(duì)不同N 沉降的響應(yīng)差異,旨在為研究N 沉降對(duì)區(qū)域森林生態(tài)系統(tǒng)的土壤生態(tài)學(xué)過(guò)程及維護(hù)土壤生態(tài)系統(tǒng)的穩(wěn)定性提供理論依據(jù)。
1 研究地區(qū)與研究方法
1.1 研究區(qū)概況
在云南省新平縣磨盤山森林生態(tài)系統(tǒng)國(guó)家定位觀測(cè)研究站(23°46′18″~23°54′34″ N,101°16′06″~101°16′12″ E)設(shè)置試驗(yàn)樣地。該地區(qū)屬于中亞熱帶氣候,年降水量1 050 mm,年平均氣溫15 ℃,極端最高氣溫為33.0 ℃,極端最低氣溫為-2.2 ℃。磨盤山海拔1 260.0~2 614.4 m,以第三紀(jì)古紅土發(fā)育的山地紅壤、玄武巖紅壤和黃棕壤為主[19]。該區(qū)植被類型隨海拔的升高呈明顯的垂直分布特征,主要分布的森林類型為針葉林(華山松林、云南松林)、亞熱帶常綠闊葉林和高山矮林(高山櫟林)等。
1.2 試驗(yàn)設(shè)計(jì)
2017 年10 月,在研究站內(nèi)4 種典型森林類型(云南松林、華山松林、常綠闊葉林和高山櫟林)中(表1),設(shè)置3 個(gè)20 m × 20 m 的樣地,在每個(gè)樣地中隨機(jī)設(shè)置4 個(gè)3 m × 3 m 的小樣方,對(duì)應(yīng)4 種不同施N 濃度,根據(jù)林地實(shí)際情況,將高濃度樣地設(shè)置在下坡面,以盡量減小坡面徑流泥沙引起的氮素遷移對(duì)土壤酶活性的影響。樣方之間距離大于1 m,防止相互干擾。
本實(shí)驗(yàn)參考美國(guó)Harvard Forest 模擬氮沉降和華西雨屏區(qū)模擬氮沉降等試驗(yàn)中的實(shí)驗(yàn)設(shè)置[14-17],結(jié)合我國(guó)西南地區(qū),如四川盆地西緣[20]、昆明東郊[21]及西雙版納[22]等地的年N 沉降量水平,以推測(cè)該地區(qū)未來(lái)可能的N 沉降趨勢(shì)。本實(shí)驗(yàn)設(shè)定N 沉降處理分4 個(gè)水平:對(duì)照(CK,0 g N·m-2·a-1)、低N(LN,5 g N·m-2·a-1)、中N(MN,15 g N·m-2·a-1)、高N(HN,30 g N·m-2·a-1),每個(gè)水平3 個(gè)重復(fù)。將年施用量均分成12 份,從2018 年1 月初開始,每月用不同濃度N 對(duì)各樣方進(jìn)行定量模擬N 沉降處理。方法為在750 mL 去離子水中溶解各N 濃度水平所需NH4NO3(相當(dāng)于全年新增降水1.0 mm),用噴霧器在3 個(gè)水平樣方中來(lái)回均勻噴灑,為減少水分增加對(duì)土壤環(huán)境與土壤酶活性的影響,對(duì)照組同時(shí)段噴灑750 mL 去離子水。
1.3 樣品采集、處理與測(cè)定
于施N 一年后(2019 年1 月)采集0~5、5~10 和10~20 cm 土壤樣品,每個(gè)樣方用隨機(jī)數(shù)字定位法隨機(jī)取3 鉆后混合,帶回實(shí)驗(yàn)室,過(guò)2 mm篩,置于無(wú)菌袋中。用四分法將樣品均分,分別進(jìn)行保存:一份室溫自然風(fēng)干保存,用于天門冬酰胺酶、脲酶、蔗糖酶的測(cè)定;一份冷藏于4 ℃冰箱,用于磷酸酶、過(guò)氧化氫酶、多酚氧化酶的測(cè)定;另一份冷凍于-40 ℃冰箱,用于蛋白酶、淀粉酶的測(cè)定。
土壤酶活性的測(cè)定:實(shí)驗(yàn)通過(guò)自主添加基質(zhì)進(jìn)行土壤培養(yǎng),測(cè)定反應(yīng)產(chǎn)物和消耗基質(zhì)量來(lái)表示酶活性,為了排除土壤本身含有的基質(zhì)成分影響,每個(gè)實(shí)驗(yàn)都設(shè)置無(wú)基質(zhì)-純水和無(wú)土壤兩種對(duì)照,蔗糖酶活性采用3,5-二硝基水楊酸比色法測(cè)定;淀粉酶活性采用3,5-二硝基水楊酸比色法測(cè)定;脲酶活性采用靛酚比色法測(cè)定;酸性磷酸酶活性采用磷酸苯二鈉比色法測(cè)定;過(guò)氧化氫酶活性采用高錳酸鉀滴定法測(cè)定;蛋白酶活性采用茚三酮比色法測(cè)定;天門冬酰胺酶活性采用擴(kuò)散法測(cè)定;多酚氧化酶采用碘量滴定法測(cè)定[6]。
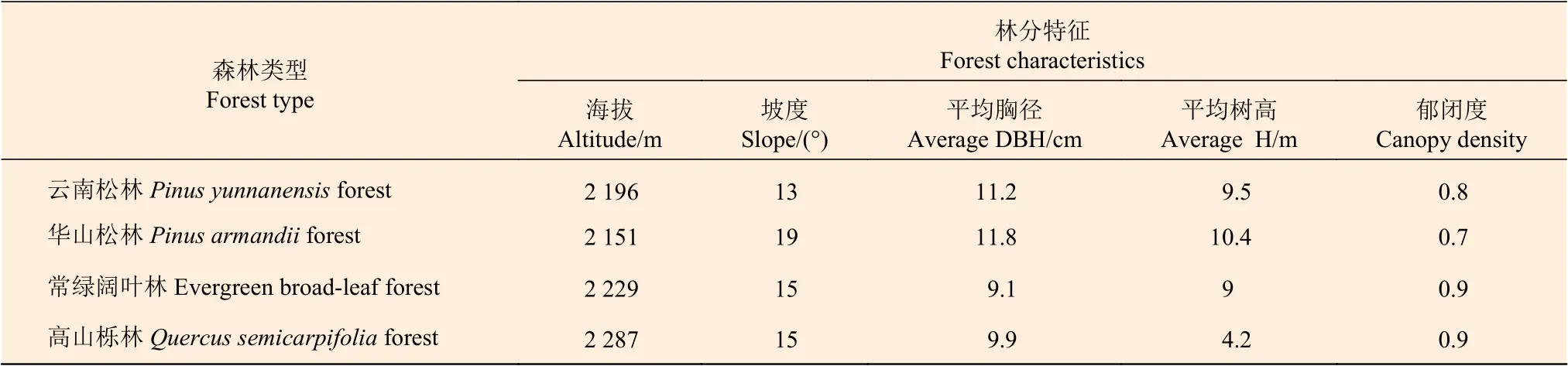
表1 4 種森林類型立地特征Table 1 Site characteristics of four forest types
1.4 數(shù)據(jù)處理
統(tǒng)計(jì)數(shù)據(jù)、繪制圖表采用Microsoft Office Excel 2010。數(shù)據(jù)分析采用SPSS 23.0 中,其中,單因素方差分析法(One-way ANOVA)用以分析不同林分間、不同土層間及不同N 沉降處理間土壤酶活性差異特征;多因素方差分析法(Multivariate ANOVA)用以分析土壤深度、施N 水平和森林類型對(duì)土壤酶活性的影響。顯著性水平設(shè)定為ɑ= 0.05。
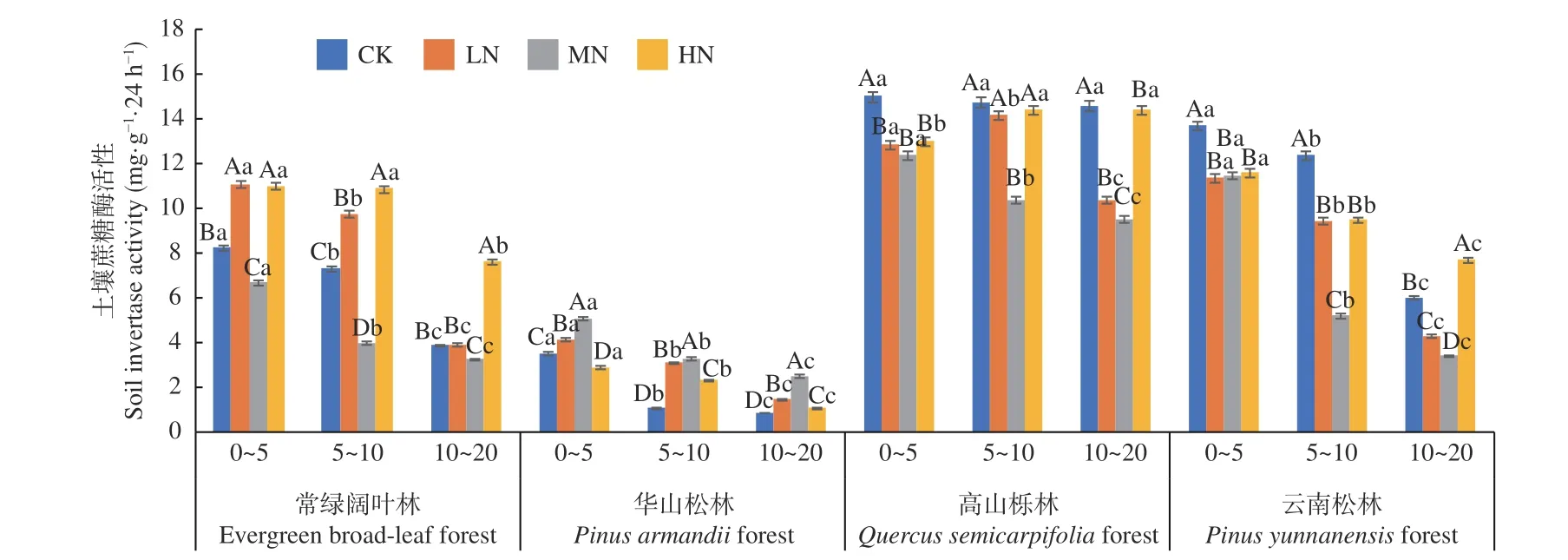
圖1 不同施N 水平4 種林地各土層土壤蔗糖酶活性變化Fig. 1 Changes of soil invertase activities in different soil layers of four forest types under different N deposition
2 結(jié)果與分析
2.1 不同施N 水平4 種林地各土層土壤酶活性的變化特征
在土壤深度、施N 水平和森林類型三者交互作用下,高山櫟林CK 處理下0~5 cm 土層蔗糖酶活性最高(15.035 mg·g-1·24 h-1),華山松林CK 處理下10~20 cm 土層活性最低(0.878 mg·g-1·24 h-1)。常綠闊葉林、華山松林和云南松林下土壤蔗糖酶活性均表現(xiàn)為隨土層深度增加逐漸降低(圖1),3 種林分不同土層平均值為0~5 cm (12.04~3.91 mg·g-1·24 h-1) > 5~10 cm (9.14~2.43 mg·g-1·24 h-1) >10~20 cm (5.37~1.48 mg·g-1·24 h-1),降幅分別在64.7%~30.5%、75.2%~50.7% 和70.4%~33.5%,其中華山松林0~5 cm 土層降幅最高。高山櫟林中除LN 處理下隨著土層深度增加呈現(xiàn)先增加后降低的趨勢(shì),HN 下隨著土層深度增加逐漸增加(增幅為10.8%),其他土層蔗糖酶活性均表現(xiàn)為隨土層深度增加逐漸降低。華山松林土壤蔗糖酶活性低于高山櫟林80.0%,變化范圍為0.878~13.704 mg·g-1·24 h-1,且存在顯著差異(P< 0.05)。在不同施N濃度下,常綠闊葉林土壤蔗糖酶活性在LN 與HN 下顯著高于CK 處理(P< 0.05);華山松林表現(xiàn)為先促進(jìn),后在HN 抑制,林下土壤蔗糖酶活性在LN 與MN 顯著高于CK 處理(P< 0.05);高山櫟林與云南松林表現(xiàn)為先抑制,后在HN 促進(jìn),林下土壤蔗糖酶活性在LN、MN 處理下顯著低于CK 處理(P< 0.05)。其中,高山櫟林土壤蔗糖酶活性表現(xiàn)最高,總體平均值為:高山櫟林(13.01 mg·g-1·24 h-1) > 云南松林(8.85 mg·g-1·24 h-1) > 常綠 闊 葉 林(7.31 mg·g-1·24 h-1) > 華 山 松 林(2.61 mg·g-1·24 h-1)。
從圖2 可以看出,在土壤深度、施N 水平和森林類型三者交互作用下,云南松林LN 處理下0~5 cm 土 層 淀 粉 酶 活 性 最 高(1.437 mg·g-1·24 h-1),云南松林MN 處理下10~20 cm 土層活性最低(0.103 mg·g-1·24 h-1)。常綠闊葉林、華山松林、云南松林、高山櫟林土壤淀粉酶活性均表現(xiàn)為隨土層深度增加逐漸降低,降幅分別在66.1%~22.1%、76.8%~21.7%、67.7%~21.5%和70.9%~21.9%,其中華山松林MN 處理降幅最高,4 種林分不同土層平均值為0~5 cm (0.91~0.40 mg·g-1·24 h-1) > 5~10 cm(0.71~0.33 mg·g-1·24 h-1) > 10~20 cm(0.52~0.22 mg·g-1·24 h-1),且存在顯著差異(P< 0.05)。在不同施N 濃度中,常綠闊葉林表現(xiàn)為先抑制,后在HN 促進(jìn),林下土壤淀粉酶活性在LN、MN 處理下顯著低于CK 處理(P< 0.05);華山松林也表現(xiàn)為先抑制,后促進(jìn),林下土壤淀粉酶活性在MN 與HN 處理下顯著高于CK 處理(P<0.05);高山櫟林表現(xiàn)為施氮?jiǎng)t抑制,但隨著土層深度的增加,HN 處理抑制程度降低,林下土壤淀粉酶活性在LN、MN 和HN 處理下均低于CK 處理;云南松林表現(xiàn)為先促進(jìn),后抑制,林下土壤淀粉酶活性在LN 處理下顯著高于CK 處理(P<0.05)。其中,云南松林表現(xiàn)最高,總體平均值為:云南松林(0.71 mg·g-1·24 h-1) > 高山櫟林(0.52mg·g-1·24 h-1) > 華山松林(0.39 mg·g-1·24 h-1) > 常綠闊葉林(0.32 mg·g-1·24 h-1)。
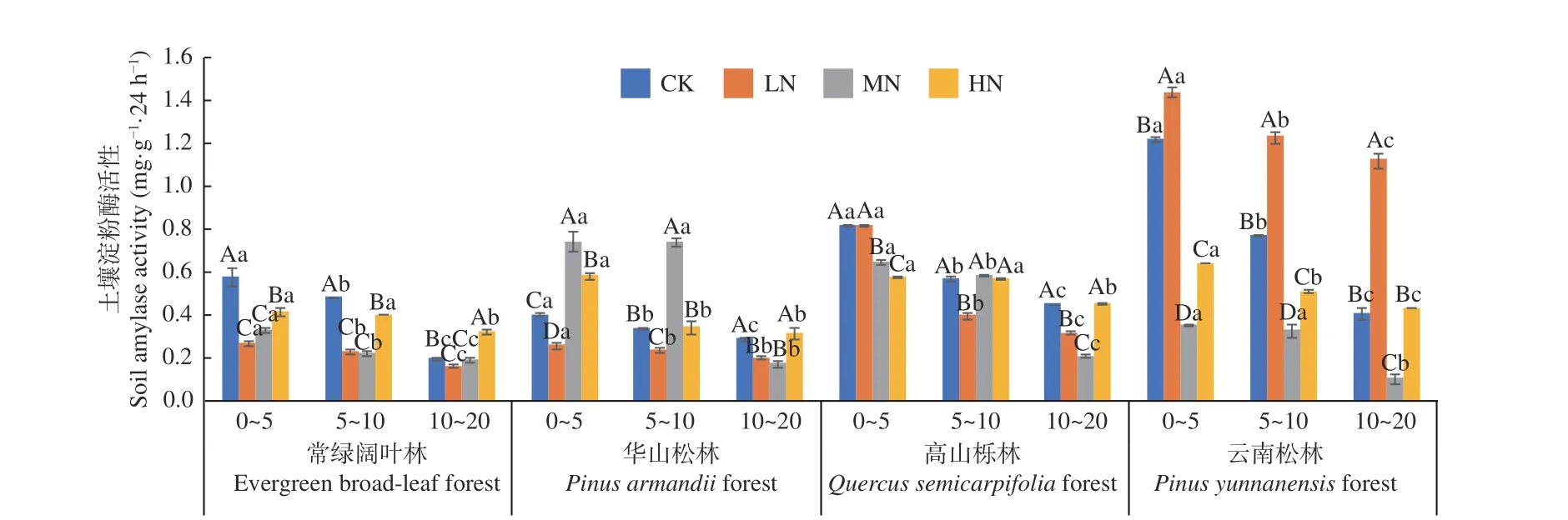
圖2 不同施N 水平4 種林地各土層土壤淀粉酶活性變化Fig. 2 Changes of soil amylase activities in different soil layers of four forest types under different N deposition
圖3 表明,在土壤深度、施N 水平和森林類型三者交互作用下,華山松林LN 處理下0~5 cm土層脲酶活性最高(0.777 mg·g-1·24 h-1),高山櫟林CK 處理下10 ~20 cm 土層活性最低(0.251 mg·g-1·24 h-1)。4 種林分不同土層間脲酶活性差異不顯著(P> 0.05),但各土層平均值表現(xiàn)為隨土層深度增加逐漸降低,不同土層平均值為0~5 cm (0.62~0.50 mg·g-1·24 h-1) > 5~10 cm (0.61~0.45 mg·g-1·24 h-1) > 10~20 cm (0.58~0.37 mg·g-1·24 h-1)。在不同施N 濃度中,除高山櫟林外,3 種林分土壤脲酶活性均表現(xiàn)為先增加后降低,但常綠闊葉林土壤脲酶活性在各處理下差異均不顯著(P> 0.05),華山松林則在LN 處理下顯著高于CK 處理(P< 0.05),高山櫟林與云南松林在LN、MN 和HN 處理下均顯著高于CK 處理(P< 0.05),增幅分別在36.9%~86.7% 和40.7%~62.2%。其中,華山松林表現(xiàn)最高,總體平均值為:華山松林(0.60 mg·g-1·24 h-1) >常綠闊 葉 林(0.51 mg·g-1·24 h-1) > 高山櫟林(0.49 mg·g-1·24 h-1) > 云南松林(0.44 mg·g-1·24 h-1),華山松林土壤脲酶活性高于云南松林36.4%。
圖4 表明,在土壤深度、施N 水平和森林類型三者交互作用下,華山松林LN 處理下0~5 cm 土層天門冬酰胺酶活性最高(0.434 mg·g-1·24 h-1),常綠闊葉林CK 處理下10~20 cm 土層活性最低(0.007 mg·g-1·24 h-1)。4 種林分下土壤天門冬酰胺酶活性均表現(xiàn)為隨土層深度增加逐漸降低,降幅分別在98.1%~48.3%、88.6%~30.7%、83.4%~37.3%和83.6%~55.8%,其中常綠闊葉林CK 處理降幅最高,4 種林分不同土層平均值為0~5 cm(0.36~0.19 mg·g-1·24 h-1) > 5~10 cm (0.17~0.06mg·g-1·24 h-1) > 10~20 cm (0.09~0.05 mg·g-1·24 h-1),且存在顯著差異(P< 0.05),其變化范圍為0.007~0.434 mg·g-1·24 h-1。在不同施N 濃度中,常綠闊葉林和華山松林土壤天門冬酰胺酶活性隨著施N 水平的增加均表現(xiàn)為先增加,后減少,高山櫟林和云南松林則表現(xiàn)為先增加,后減少,再增加。4 種林分在LN 處理促進(jìn),其中華山松林在0~5 cm 土層對(duì)比CK 處理增幅最大,為75.8%。常綠闊葉林表現(xiàn)最高,總體平均值為:常綠闊葉林(0.20 mg·g-1·24 h-1) > 高山櫟林(0.16 mg·g-1·24 h-1) > 華 山 松 林(0.14 mg·g-1·24 h-1) > 云 南 松林(0.10 mg·g-1·24 h-1),常綠闊葉林土壤天門冬酰胺酶活性高于云南松林50.0%。
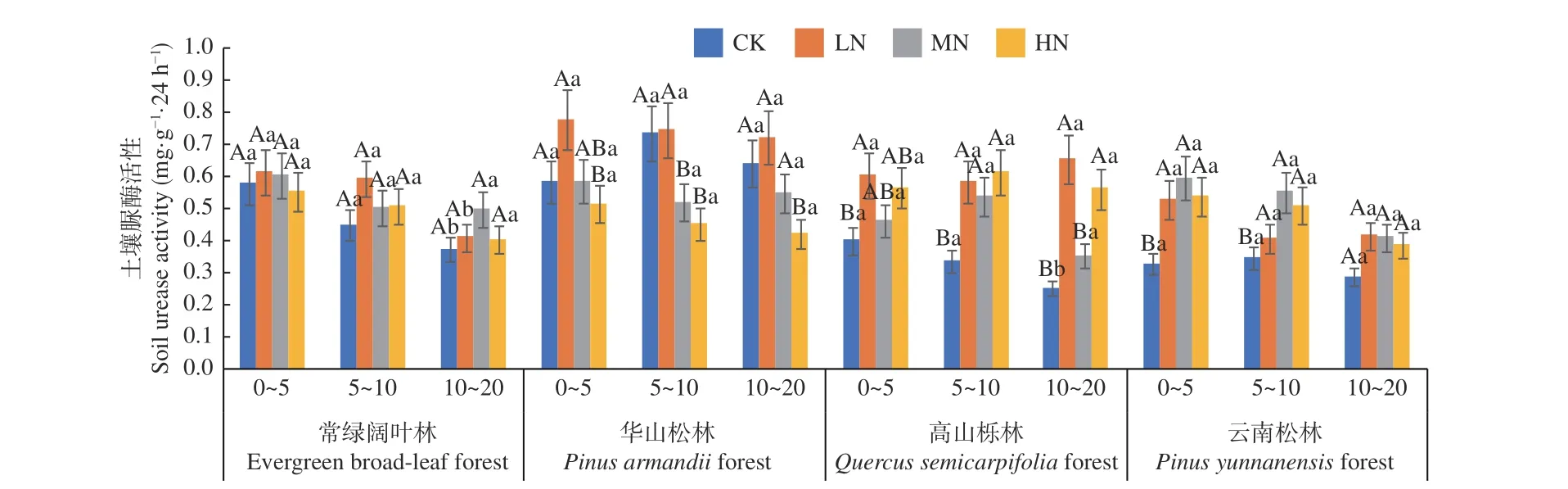
圖3 不同施N 水平4 種林地各土層土壤脲酶活性變化Fig. 3 Changes of soil urease activities in different soil layers of four forest types under different N deposition
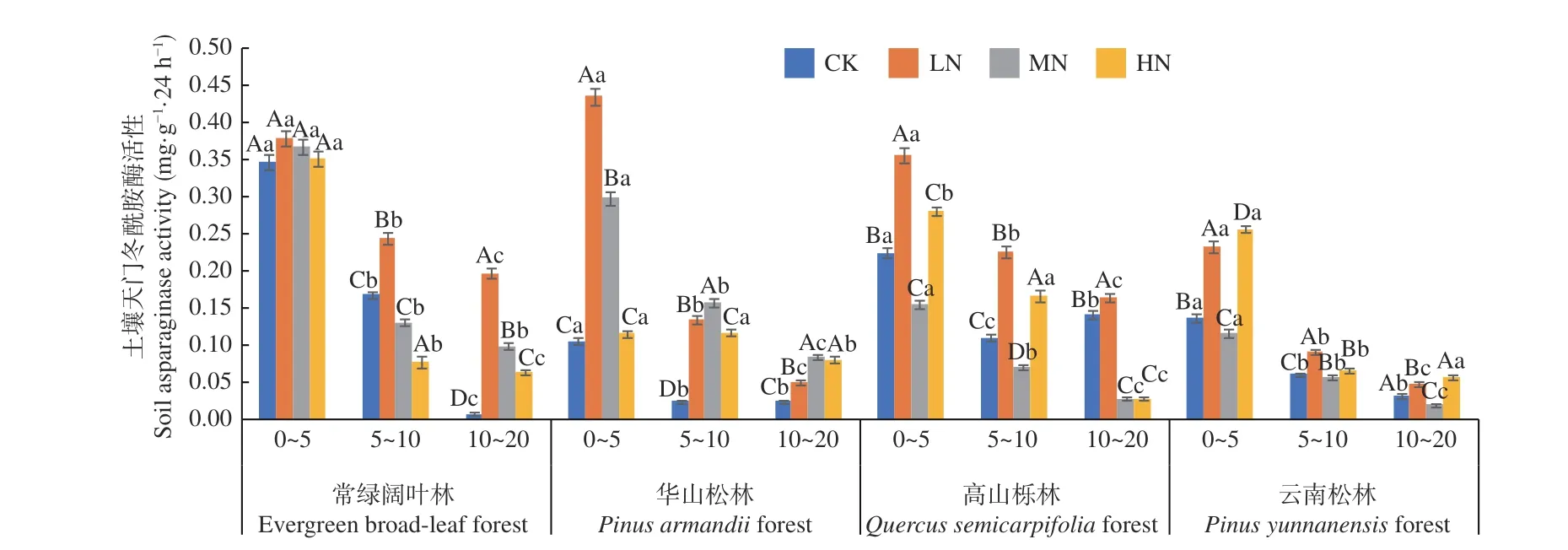
圖4 不同施N 水平4 種林地各土層土壤天門冬酰胺酶活性變化Fig. 4 Changes of asparaginase activity in different soil layers of four forest types under different N deposition
圖5 表明,在土壤深度、施N 水平和森林類型三者交互作用下,常綠闊葉林CK 處理下0~5 cm 土層酸性磷酸酶活性最高(2.879 mg·g-1·24 h-1),高山櫟林LN 處理下10~20 cm 土層活性最低(1.372 mg·g-1·24 h-1)。酸性磷酸酶活性在4 種林分中均表現(xiàn)為隨土層深度增加逐漸降低,降幅分別在34.0%~5.6%、35.0%~13.6%、41.0%~10.9%、和40.2%~11.1%,其中高山櫟林LN 處理降幅最高,4 種林分不同土層平均值為0~5 cm (2.47~2.14 mg·g-1·24 h-1) > 5~10 cm (2.23~1.74 mg·g-1·24 h-1) > 10~20 cm (1.92~1.54 mg·g-1·24 h-1),其變化范圍為1.372~2.879 mg·g-1·24 h-1,且存在顯著差異(P< 0.05)。在不同施N 濃度中,4 種林分土壤酸性磷酸酶活性在各處理下存在顯著差異(P< 0.05)。其中常綠闊葉林在HN 處理下受到抑制,降幅在15.1%~5.1%;華山松在HN 處理下受到促進(jìn),增幅在1.9%~22.1%;高山櫟林與云南松林LN 處理在10~20 cm 土壤酸性磷酸酶受到抑制,降幅為17.2%和11.8%,在其他土層則表現(xiàn)為促進(jìn)作用,增幅為20.0%~20.7%和6.7%~9.3%。常綠闊葉林表現(xiàn)最高,總體平均值為:常綠闊葉林(2.21 mg·g-1·24 h-1) > 華山松林(2.04 mg·g-1·24 h-1) >高山櫟林(1.96 mg·g-1·24 h-1) >云南松林(1.81 mg·g-1·24 h-1),常綠闊葉林土壤酸性磷酸酶活性高于云南松林22.1%。
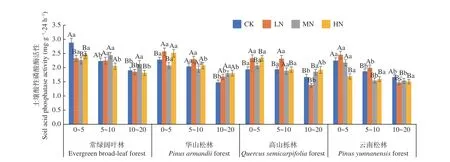
圖5 不同施N 水平4 種林地各土層土壤酸性磷酸酶活性變化Fig. 5 Changes of soil acid phosphatase activity in different soil layers of four forest types under different N deposition
圖6 表明,在土壤深度、施N 水平和森林類型三者交互作用下,高山櫟林LN 處理下0~5 cm 土 層 蛋 白 酶 活 性 最 高(1.297 mg·g-1·24 h-1),云南松林MN 處理下10~20 cm 土層活性最低(0.625 mg·g-1·24 h-1)。除HN 處理下的華山松林和高山櫟林外,其他土層蛋白酶活性均表現(xiàn)為隨土層深度增加逐漸降低,降幅分別在33.8%~22.8%、17.4%~13.4%、41.5%~3.2%、和27.3%~4.9%,其中高山櫟林MN 處理降幅最大,4 種林分不同土層 平 均 值 為0 ~5 cm (1.21~0.85 mg·g-1·24 h-1) >5~10 cm (1.14~0.80 mg·g-1·24 h-1) > 10~20 cm(1.02~0.73 mg·g-1·24 h-1),其變化范圍為0.625~1.297 mg·g-1·24 h-1,且存在顯著差異(P< 0.05)。在不同施N 濃度中,常綠闊葉林土壤蛋白酶活性表現(xiàn)為先增加后降低的趨勢(shì),在LN 與MN 處理下高于CK 處理,增幅在8.4%~30.2%。高山櫟林在LN 處理下表現(xiàn)為促進(jìn)作用,增幅在9.5%~23.8%。華山松林與云南松林在各處理下差異不顯著。其中,高山櫟林表現(xiàn)最高,總體平均值為:高山櫟林(1.12 mg·g-1·24 h-1) > 常綠闊葉林(0.91 mg·g-1·24 h-1) > 華山松林(0.83 mg·g-1·24 h-1) >云南松林(0.80 mg·g-1·24 h-1),高山櫟林土壤蛋白酶活性高于云南松林34.9%。
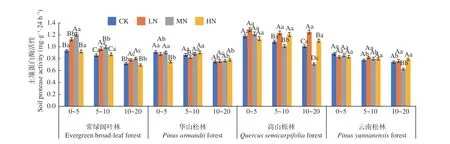
圖6 不同施N 水平4 種林地各土層蛋白酶活性變化Fig. 6 Changes of protease activities in different soil layers of four forest types under different N deposition
圖7 表明,在土壤深度、施N 水平和森林類型三者交互作用下,華山林LN 處理下0~5 cm 土層過(guò)氧化氫酶活性最高(0.973 mg·g-1·20 min-1),云南松林LN 處理下0~5 cm 土層活性最低(0.209 mg·g-1·20 min-1)。常綠闊葉林與云南松林土壤過(guò)氧化氫酶活性表現(xiàn)為5~10 cm 土層顯著高于0~5 cm 土層(P< 0.05),不同土層平均值為5~10 cm (0.48~0.45 mg·g-1·20 min-1) > 0~5 cm (0.44~0.33 mg·g-1·20 min-1) > 10~20 cm (0.41~0.29 mg·g-1·20 min-1)。華山松林與高山櫟林土壤過(guò)氧化氫酶活性則表現(xiàn)為隨土層深度增加逐漸降低,降幅分別在55.6%~14.4%和28.1%~14.4%,兩種林分不同土層平 均 值 為0~5 cm (0.47~0.36 mg·g-1·20 min-1) >5~10 cm (0.32~0.30 mg·g-1·20 min-1) > 10~20 cm(0.28~0.27 mg·g-1·20 min-1),且存在顯著差異(P<0.05)。4 種林分土壤過(guò)氧化氫酶活性變化范圍為0.209~0.673 mg·g-1·20 min-1。在不同施N 濃度中,常綠闊葉林與華山松林土壤過(guò)氧化氫酶活性隨著施N 水平的增加先增加后減小。高山櫟林在HN 處理下顯著高于CK 處理(P< 0.05),增幅為3.9%、16.9%、21.1%。云南松林在HN 處理下顯著高于CK處理,增幅為30.1%、57.9%、9.1%(P<0.05)。其中,常綠闊葉林表現(xiàn)最高,總體平均值為:常綠闊葉林(0.45 mg·g-1·20 min-1) > 云南松林(0.36 mg·g-1·20 min-1) > 華山 松林(0.34 mg·g-1·20min-1) > 高山櫟林(0.32 mg·g-1·20 min-1),常綠闊葉林土壤過(guò)氧化氫酶活性高于高山櫟林40.6%。
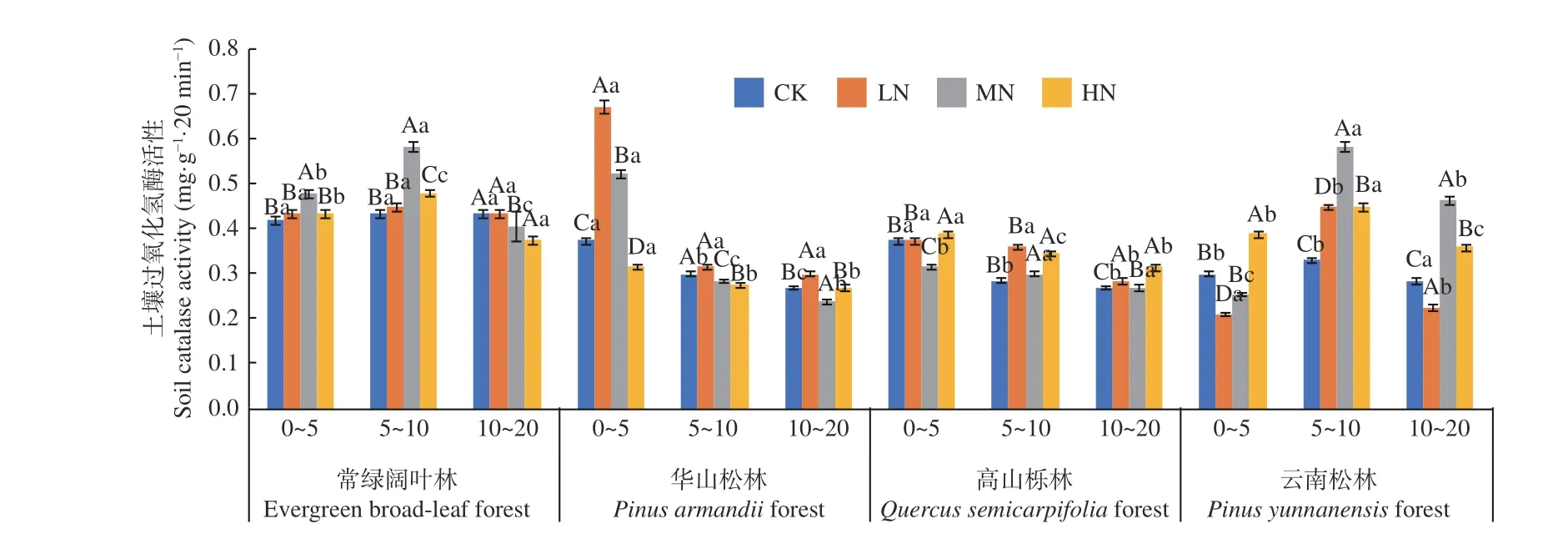
圖7 不同施N 水平4 種林地各土層過(guò)氧化氫酶活性變化Fig. 7 Changes of catalase activity in different soil layers of four forest types under different N deposition
圖8 表明,在土壤深度、施N 水平和森林類型三者交互作用下,云南松林CK 處理下0~5 cm土層多酚氧化酶活性最高(1.969 mg·g-1·20 min-1),高山櫟林MN 處理下0~5 cm 土層活性最低(0.156 mg·g-1·20 min-1)。常綠闊葉林與華山松林土壤多酚氧化酶活性表現(xiàn)為隨土層深度增加逐漸降低,降幅分別在73.3%~52.6%和50.0%~11.1%,兩種林分不同土層平均值為0~5 cm (1.49~1.23 mg·g-1·20 min-1) > 5~10 cm (1.02~0.87 mg·g-1·20 min-1) >10~20 cm (0.73~0.59 mg·g-1·20 min-1),且存在顯著差異(P< 0.05)。高山櫟林與云南松林則大部分表現(xiàn)為5~10 cm > 0~5 cm > 10~20 cm,10~20 cm 土 層 為5 ~10 cm 的23.6%~74.4%,0~5 cm 土層為5~10 cm 的15.4%~94.7%。4 種林分土壤多酚氧化酶變化范圍為0.156~1.969 mg·g-1·20 min-1。在不同施N 濃度中,常綠闊葉林土壤多酚氧化酶活性在HN 處理下顯著抑制(P < 0.05),降幅為42.7%~35.7%;華山松林與高山櫟林在LN、MN 和HN 處理下基本表現(xiàn)為抑制,降幅在49.4%~9.1%;云南松林在HN 處理下高于CK 處理,增幅在80.0%~27.7%。其中,常綠闊葉林表現(xiàn)最高,總體平均值為:常綠闊葉林(1.03 mg·g-1·20 min-1) >華山松林(0.94 mg·g-1·20 min-1) > 云南松林(0.89mg·g-1·20 min-1) > 高山櫟林(0.84 mg·g-1·20 min-1),常綠闊葉林土壤過(guò)氧化氫酶活性高于高山櫟林22.6%。

圖8 不同施N 水平4 種林地各土層多酚氧化酶活性變化Fig. 8 Changes of polyphenol oxidase activities in different soil layers of four Forest types under different N deposition
2.2 不同土壤深度、不同施N 水平和不同森林類型對(duì)土壤酶活性的影響
多因素方差分析表明(表2):除施N 水平對(duì)酸性磷酸酶以及土壤深度、施N 水平和森林類型三者交互作用對(duì)脲酶無(wú)顯著影響外(P>0.05),土壤深度、施N 水平、森林類型和三者交互作用對(duì)土壤蔗糖酶、淀粉酶、脲酶、天門冬酰胺酶、酸性磷酸酶、蛋白酶、過(guò)氧化氫酶和多酚氧化酶均具有極顯著影響(P< 0.01)。其中,蔗糖酶、淀粉酶、蛋白酶和過(guò)氧化氫酶表現(xiàn)為:森林類型的影響最大,土壤深度次之,施N 水平和三者交互作用最小;脲酶表現(xiàn)為:森林類型影響最大,施N 水平次之,土壤深度和三者交互作用最小;天門冬酰胺酶和酸性磷酸酶表現(xiàn)為:土壤深度影響最大,森林類型次之,施N 水平和三者交互作用最小;多酚氧化酶表現(xiàn)為:土壤深度的影響最大,施N 水平次之,森林類型和三者交互作用最小。
土壤深度對(duì)土壤酶活性的影響力為:天門冬酰胺酶>蔗糖酶>淀粉酶>多酚氧化酶>過(guò)氧化氫酶>蛋白酶>酸性磷酸酶>脲酶;施N 水平對(duì)土壤酶活性的影響力為:蔗糖酶>天門冬酰胺酶>淀粉酶>多酚氧化酶>過(guò)氧化氫酶>蛋白酶>脲酶>酸性磷酸酶;森林類型對(duì)土壤酶活性的影響力為:蔗糖酶>淀粉酶>天門冬酰胺酶>過(guò)氧化氫酶>蛋白酶>多酚氧化酶>酸性磷酸酶>脲酶。三者交互作用對(duì)土壤酶活性的影響力為:天門冬酰胺酶>過(guò)氧化氫酶>淀粉酶>蔗糖酶>多酚氧化酶>蛋白酶>酸性磷酸酶>脲酶。
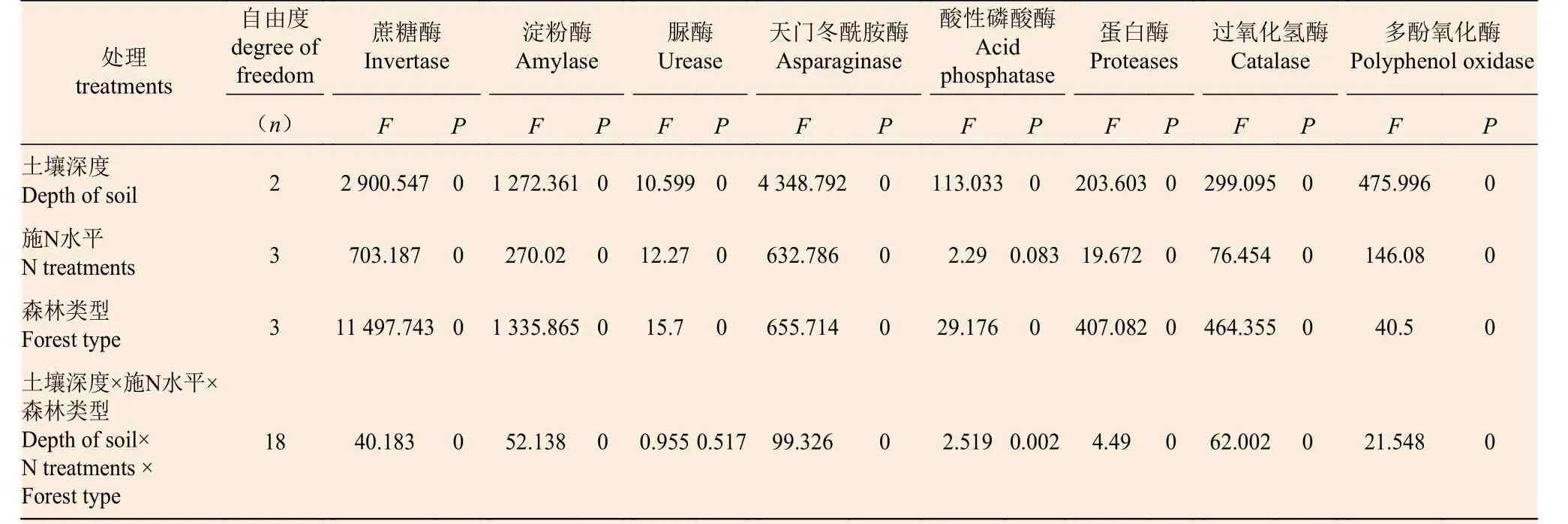
表2 不同土壤深度、不同施N 水平和不同森林類型對(duì)土壤酶活性的多因素方差分析Table 2 Multivariate ANOVA of soil enzyme activity by different soil depth,different N treatments and different forest types
3 討論
3.1 土壤酶活性對(duì)不同土壤深度的響應(yīng)特征
本研究中,土層深度對(duì)8 種土壤酶活性均表現(xiàn)為極顯著影響。其中4 種林分下土壤淀粉酶、脲酶、天門冬酰胺酶、酸性磷酸酶活性影響均表現(xiàn)為:隨著土層加深而逐漸降低,N 沉降并未改變土壤酶活性的垂直分布格局。這與陳影[23]對(duì)楓楊-桑樹混交林(Pterocarya stenopteraC. DC.-Morus albaLinn.)、柳樹純林(Salix babylonicaL.)、女貞純林(Ligustrum lucidumAit.)和欒樹純林(Koelreuteria paniculataLaxm.)4 種溫帶樹種下土壤蔗糖酶、過(guò)氧化氫酶、蛋白酶活性研究,以及江玉梅[24]對(duì)江西馬尾松純林(Pinus massonianaLamb.)、馬尾松-楓香-木荷混交林(Pinus massonianaLamb.-Liquidamber formosanaHance-Schima superbaGardn. )、 木 荷 純 林(Schima superbaGardn.)和楓香純林(Liquidambar formosanaHance)4 種人工林林分下土壤天門冬酰胺酶活性的垂直性變化研究結(jié)果一致。這可能是因?yàn)殡S著土層加深,土壤物理性質(zhì)發(fā)生變化,微生物含量降低,進(jìn)而導(dǎo)致土壤酶活性降低;同時(shí)土壤全N 含量的表聚性,使表層土壤含氮有機(jī)物轉(zhuǎn)化速率增加,從而使土壤酶活性進(jìn)一步增強(qiáng)[25]。
不同林分下土壤脲酶活性垂直分布差異不顯著,但總體上仍表現(xiàn)為隨土層深度增加而降低。李佳等[26]和劉星[27]分別對(duì)羅甸縣米槁(Cinnamomum migaoH. W. Li)及油松天然林(Pinus tabuliformisCarriere)模擬N 沉降下土壤酶的研究同樣表明,不同土層之間脲酶活性差異不顯著(P> 0.05)。有研究表明,當(dāng)森林土壤N 含量較高時(shí),外源N 抑制土壤脲酶活性;反之,則促進(jìn)土壤脲酶活性,在本研究中,土壤脲酶活性未受到N 處理的顯著影響,推測(cè)施N 前后土壤N 濃度均處于平均范圍內(nèi),今后研究可考慮適當(dāng)增加施N 濃度,以研究N 沉降增加對(duì)土壤脲酶活性的影響。而HN 處理下華山松林和高山櫟林土壤蛋白酶活性、各施N 處理下常綠闊葉林和云南松林土壤過(guò)氧化氫酶活性、各施N 處理下高山櫟林與云南松林土壤多酚氧化酶活性則多表現(xiàn)出0~5 cm、5~10 cm和10~20 cm 土層先降低后升高的現(xiàn)象,這可能是由于過(guò)量的N 施入土壤表層使土壤發(fā)生負(fù)反饋機(jī)制[28],從而抑制了土壤蛋白酶、過(guò)氧化氫酶和多酚氧化酶的活性,但其抑制機(jī)理還有待進(jìn)一步研究。
3.2 土壤酶活性對(duì)不同施N 水平的響應(yīng)特征
本研究對(duì)8 種酶的研究中,蔗糖酶、天門冬酰胺酶與淀粉酶對(duì)施N 水平的響應(yīng)最顯著。土壤蔗糖酶活性與土壤有機(jī)質(zhì)、氮、磷含量,微生物數(shù)量及土壤呼吸強(qiáng)度有關(guān)[6],土壤淀粉酶則是碳素循環(huán)中的一種重要的酶。其中,LN 處理顯著增加了常綠闊葉林土壤蔗糖酶活性和云南松林土壤淀粉酶活性(P< 0.01),這與肖永翔等[29]對(duì)四川雅安常綠闊葉林土壤蔗糖酶活性變化的研究結(jié)果一致,原因可能是相對(duì)較低的N 沉降促進(jìn)了林下植被的生長(zhǎng),進(jìn)而向土壤中提供了更多的C;MN 與HN 處理對(duì)土壤蔗糖酶與淀粉酶活性多表現(xiàn)為抑制作用,如HN 處理抑制了高山櫟林、云南松林土壤蔗糖酶與土壤淀粉酶活性,但未達(dá)到顯著性(P>0.05),可能是高濃度的N 抑制了植物根系活性,使土壤pH降低,土壤硝化作用增強(qiáng),土壤有效C降低[30],從而抑制了土壤蔗糖酶與淀粉酶活性,而土壤蔗糖酶活性的降低,減少了土壤植物與微生物營(yíng)養(yǎng)源,進(jìn)一步影響了土壤生物學(xué)活性強(qiáng)度。此外,不同森林類型的植株對(duì)碳的吸收和利用能力不同[31],也可能是造成土壤蔗糖酶與淀粉酶活性隨著N 濃度的增加出現(xiàn)不同的變化趨勢(shì)的原因。土壤天門冬酰胺酶在氮代謝中起著重要作用,能夠反映土壤中含氮有機(jī)化合物的轉(zhuǎn)化能力。4 種N 處理對(duì)4 種林分土壤天門冬酰胺酶活性均造成了顯著影響,其中,LN 處理顯著提高了4 種林分表層土壤的天門冬酰胺酶活性。可能是表層土壤很大程度上受土壤有機(jī)質(zhì)和微生物含量的影響[26],而LN 處理下N 的輸入提高了土壤N 含量,促進(jìn)酶活性,但土層的增加使土壤通透性下降,微生物量降低,抑制了土壤天門冬酰胺酶活性。
本研究對(duì)8 種酶的研究中,脲酶、酸性磷酸酶與蛋白酶對(duì)施N 水平的響應(yīng)最低。土壤脲酶在土壤中較為活躍,其活性與土壤中尿素、有機(jī)質(zhì)含量和微生物含量有關(guān)。脲酶能將土壤中的尿素水解為NH3和CO2,為植物直接提供N 素營(yíng)養(yǎng),是促進(jìn)土壤N 循環(huán)的重要組成部分,可以反應(yīng)土壤的供氮能力[32]。本研究中不同林分下土壤脲酶活性在各施N 處理間差異不顯著。李瑞瑞等[33]對(duì)墨西哥柏 (Cupressus lusitanicaMiller) 模擬N 沉降下土壤酶的研究同樣表明,不同N 沉降處理之間脲酶活性差異不顯著(P> 0.05)。這可能是由于土壤脲酶對(duì)環(huán)境條件的抗性,即脲酶具有較強(qiáng)穩(wěn)定性。有研究表明土壤脲酶的保護(hù)容量是相對(duì)恒定的值[6],外界因子對(duì)脲酶保護(hù)容量的改變具有時(shí)效性,最終都會(huì)恢復(fù)到初始狀態(tài)[34],使不同處理間土壤脲酶活性差異較小。
土壤酸性磷酸酶與土壤碳、氮、磷含量有關(guān),是評(píng)價(jià)土壤磷素生物轉(zhuǎn)化方向與強(qiáng)度的指標(biāo)。LN 處理促進(jìn)了華山松林、高山櫟林和云南松林土壤酸性磷酸酶活性,HN 則抑制了常綠闊葉林與云南松林土壤酶活性。可能是由于亞熱帶森林中有效N 富集,使LN 處理的N 增加量直接作用于土壤酶,促進(jìn)酶活性[35],而高N 下抑制則可能是因?yàn)楦逳 鹽的毒害作用,使土壤微生物群落功能受到影響[36]。土壤蛋白酶能夠水解蛋白質(zhì)肽鏈,促進(jìn)土壤微生物氮素代謝,可促進(jìn)土壤N 循環(huán),是土壤中氮礦化過(guò)程的限速酶[37]。本研究中N 沉降對(duì)土壤蛋白酶活性影響并不顯著,隨著N 濃度的增加,酶活性峰值出現(xiàn)在LN、MN 下,這與于濟(jì)通[38]等的研究結(jié)果,土壤蛋白酶活性隨著N 沉降量的增加逐漸增強(qiáng)相一致。而HN 下抑制則可能是因?yàn)橥寥繬 含量超出N 飽和點(diǎn),酶活性受到過(guò)量N 沉降的抑制作用;也可能是因?yàn)楦叩}的毒害作用,使土壤微生物群落功能受到影響,降低了酶活性[39]。
土壤過(guò)氧化氫酶有利于防止過(guò)氧化氫對(duì)生物體的毒害作用,其活性與土壤有機(jī)質(zhì)含量、微生物數(shù)量有關(guān)[40]。Frey 等[38]與本研究中常綠闊葉林研究結(jié)果類似,N 沉降對(duì)土壤過(guò)氧化氫酶活性影響不顯著。本研究中,LN 處理促進(jìn)了華山松林、高山櫟林下土壤過(guò)氧化氫酶活性,MN 處理促進(jìn)了常綠闊葉林、云南松林下土壤酶活性。從不同研究中可以發(fā)現(xiàn),施N 濃度不同會(huì)對(duì)土壤過(guò)氧化物酶活性造成促進(jìn)[41]或抑制[27,42]作用,原因可能是N 沉降雖然在一定程度上對(duì)微生物活性產(chǎn)生了影響,但由于不同森林類型下土壤環(huán)境、土壤生物及森林類型的差異性,使微生物活性的變化方向具有不確定性,進(jìn)而使土壤過(guò)氧化氫酶活性變化不同[43]。土壤多酚氧化酶屬于木質(zhì)素分解酶,其活性與凋落物分解密切相關(guān)。本研究中,N 沉降對(duì)常綠闊葉林、華山松林和高山櫟林下土壤酶活性有顯著抑制作用,并且隨著N 濃度的升高,抑制作用逐漸增強(qiáng)。這與孫宇等[44]對(duì)濕性常綠闊葉次生林的研究結(jié)果一致。原因可能是N 沉降下微生物中有關(guān)木質(zhì)素分解基因的表達(dá)和豐度的相關(guān)基因下降[45],而N 水平的逐級(jí)增加,抑制了土壤多酚氧化酶的生產(chǎn)作用,使木質(zhì)素在土壤中大量堆積,引起進(jìn)一步的抑制作用[46]。
3.3 土壤酶活性對(duì)不同森林類型的響應(yīng)特征
本研究結(jié)果表明森林類型對(duì)土壤蔗糖酶、淀粉酶、脲酶、天門冬酰胺酶、酸性磷酸酶、蛋白酶過(guò)氧化氫酶和多酚氧化酶活性均表現(xiàn)為極顯著影響,且為蔗糖酶、淀粉酶、脲酶、蛋白酶和過(guò)氧化氫酶影響最大的因素,原因可能是不同森林類型下土壤通透性和微生物種類與數(shù)量等差異,造成4 種林分對(duì)N 沉降的轉(zhuǎn)化程度各不同,施N 前后土壤N 含量出現(xiàn)差異,進(jìn)而極大程度上影響了不同土壤酶活性的變化。其中土壤天門冬酰胺酶、酸性磷酸酶、過(guò)氧化氫酶和多酚氧化酶活性在常綠闊葉林中表現(xiàn)最高,可能原因是常綠闊葉林中植物種類較多,包括米櫧(Castanopsis carlesii(Hemsl.) Hayata.)、光葉柯(Lithocarpus mairei(Schottky) Rehder)、毛蕊紅山茶(Camellia mairei(Lévl.) Melch.)等,凋落物積累較為豐富,相比其他樹種土壤碳儲(chǔ)量較高[47],能夠促進(jìn)土壤微生物活性。脲酶在華山松林中活性最高,原因可能是脲酶能為植物直接供N,而華山松林為人工林,土壤通透性較高,且處于中幼齡林階段(20 年生),生長(zhǎng)較快需要合成大量干物質(zhì),對(duì)N、P 的需求相對(duì)較高[48]。
3.4 土壤酶活性對(duì)不同土壤深度、不同施N 水平和不同森林類型交互作用的響應(yīng)特征
由森林類型、土壤深度和施N 水平對(duì)土壤酶活性的多因素方差分析結(jié)果可知:森林類型和土壤深度是影響N 沉降下土壤蔗糖酶、淀粉酶、脲酶、天門冬酰胺酶、酸性磷酸酶、蛋白酶和過(guò)氧化氫酶活性變化差異的重要因素。土壤多酚氧化酶對(duì)施N 水平反應(yīng)敏感,對(duì)森林類型反應(yīng)較不敏感,這將有利于對(duì)比研究不同區(qū)域?qū) 沉降的響應(yīng)特征。天門冬酰胺酶對(duì)不同土壤深度的響應(yīng)最為敏感,蔗糖酶對(duì)不同施N 水平和不同森林類型的響應(yīng)最為敏感;天門冬酰胺酶對(duì)不同土壤深度、不同施N 水平和不同森林類型三者交互作用的響應(yīng)最為敏感。
天門冬酰胺酶來(lái)源于植物根系,能酶促天門冬酰胺生成天門冬氨酸和氨,和氮循環(huán)密切相關(guān),也是土壤中最活躍的水解酶類之一[49]。本研究中土壤天門冬酰胺酶的垂直差異表現(xiàn)出極顯著水平,而LN 處理顯著提高了4 種林分各土層土壤天門冬酰胺酶活性,原因可能是細(xì)根作為林木根系中最活躍的部分,其分布主要集中于土壤表層,并隨著土壤深度增加逐漸減小[50],使土壤天門冬酰胺酶活性隨土壤深度的增加而逐漸降低,穆蘭[51]對(duì)茶園生態(tài)系統(tǒng)的研究也證明了這一觀點(diǎn);而土壤C 庫(kù)中的活性C 庫(kù)能夠直接影響水解酶活性,外源N 的輸入通過(guò)影響微生物活性與數(shù)量,從而改變了土壤C 儲(chǔ)量[52],使天門冬酰胺酶對(duì)N 沉降的敏感性較高,但具體的作用機(jī)理還有待進(jìn)一步研究。
4 結(jié)論
(1)土壤酶活性的垂直分布存在明顯規(guī)律,8 種土壤酶活性均表現(xiàn)為5~10 cm 土層土壤酶活性大于10~20 cm 土層,降幅最高達(dá)76.8%,常綠闊葉林與云南松林土壤過(guò)氧化氫酶和高山櫟林與云南松林土壤多酚氧化酶活性均在0~5 cm 土層受到抑制,活性最低時(shí)僅為5~10 cm 土層活性的15.4%。
(2)施N 條件下不同林分土壤酶活性變化存在差異:LN 顯著抑制了高山櫟土壤蔗糖酶與淀粉酶活性;脲酶、蛋白酶在各N 處理下差異不顯著;除常綠闊葉林土壤酸性磷酸酶外LN 處理均顯著提高了天門冬酰胺酶和酸性磷酸在0~10 cm 土層土壤酶活性;N 沉降抑制了華山松林土壤過(guò)氧化氫酶活性,促進(jìn)了高山櫟林、云南松林土壤過(guò)氧化氫活性;對(duì)土壤多酚氧化酶則表現(xiàn)為抑制。
(3)土壤蔗糖酶、淀粉酶、脲酶、天門冬酰胺酶、酸性磷酸酶、蛋白酶、過(guò)氧化氫酶和多酚氧化酶在不同林分中表現(xiàn)出明顯差異性(P< 0.01);其中土壤天門冬酰胺酶、酸性磷酸酶、過(guò)氧化氫酶和多酚氧化酶活性在常綠闊葉林中表現(xiàn)最高,蔗糖酶和蛋白酶在高山櫟林中表現(xiàn)最高,淀粉酶在云南松林中表現(xiàn)最高,脲酶在華山松林中表現(xiàn)最高。
(4)8 種土壤酶中,蔗糖酶分別對(duì)不同施N 水平和不同森林類型的響應(yīng)最為敏感,天門冬酰胺酶分別對(duì)不同土壤深度及不同土壤深度、施N 水平和森林類型三者交互作用的響應(yīng)最為敏感。
