羊棲菜多糖對3T3-L1 脂肪細胞脂代謝的影響及其機制
2021-03-30 08:53:16張杰楊旭東石杰楊驕霞王桂云宋高臣
智慧健康 2021年4期
張杰,楊旭東★,石杰,楊驕霞,王桂云,宋高臣
(1.牡丹江醫學院,黑龍江 牡丹江 157011;2.牡丹江醫學院附屬紅旗醫院,黑龍江 牡丹江 157011)
0 引言
羊棲菜是一種隸屬于褐藻門的食用海藻,富含氨基酸、維生素和多糖等,其重要成分羊棲菜多糖(Sargassum fusiforme polysaccha-ride,SFPS)是一種具有多種生物活性的物質,具有抗氧化、抗衰老、抑制腫瘤、降血脂等作用[1-3]。本課題組前期動物實驗發現,SFPS 具有改善糖尿病大鼠胰島素抵抗的作用[4-5],但其對體外脂肪細胞的增殖及糖脂代謝的影響及其機制未見報道。本實驗進一步觀察羊棲菜多糖對3T3-L1 脂肪細胞脂代謝的影響并初步研究其作用機制。
1 材料與方法
1.1 主要試劑
羊棲菜多糖由牡丹江醫學院中心實驗室提供;3T3-L1 前脂肪細胞(美國ATCC 公司);DMEM 高糖培養液、胎牛血清(美國Gibco 公司);DOMO、MTT試劑(Sigma 公司);胰蛋白酶、引物核苷酸片段(上海生工),其他試劑為國產分析純。
1.2 主要儀器和設備
CO2恒溫培養箱(美國G.S.公司);PCR 儀(德國,Biometra);紫外分光光度計(上海元析);酶標儀(美國Biotek公司);凝膠成像系統(美國Bio-Rad公司);PCR 儀(德國,Biometra)。
1.3 實驗方法
1.3.1 細胞培養
3T3-L1 前脂肪細胞培養條件[6],37℃飽和濕度,5%CO2環境,10%胎牛血清DMEM 高糖培養液(鏈霉素100mg/L、青霉素100IU/L)培養。
1.3.2 胰島素抵抗模型建立
待分化細胞37℃飽和濕度,5%CO2培養箱,DMEM培養基中加入誘導液1(10μg/mL 胰島素、0.5mmol/LIBMX、1mmol/mL 地塞米松)培養2d。棄上清,DMEM培養基中加入誘導液2(10mg/L 胰島素),培養2d。更換DMEM 培養基,37℃,5%CO2培養箱,培養9d,細胞接近圓形,脂滴明顯聚集,細胞分化完成。分化成熟的3T3-L1 脂肪細胞分為對照組(高糖DMEM 培養基)和模型組(1mmol/mL 地塞米松),96h 后,GOD-POD方法測定培養基中葡萄糖消耗量確定模型是否成功。
1.3.3 胰島素抵抗細胞葡萄糖消耗量、甘油三酯和游離脂肪酸的測定
造模成功細胞分為3 組:模型組、SFPS 100mg/L 組 和SFPS 200mg/L 組,另 設 分 化 成 熟3T3-L1 脂肪細胞為正常對照組。上述各組繼續培養48h 后,按照試劑盒說明測定葡萄糖消耗量(glucose consumption,Glu)、游離脂肪酸(Free fatty acid,FFA)的生成量和細胞中甘油三酯(triglyceride,TG)含量。
1.3.4 MTT 法檢測細胞相對增殖率
正常對照組、模型組、SFPS 100mg/L 組和SFPS 200mg/L 組細胞,每組6 個平行孔,分別培養24h、48h、72h,加入MTT(5g/L)20μL,孵育4h,棄上清,加150μL DMSO,完全溶解,輕微振蕩12min。酶標儀570nm 測光密度(OD)值,并計算細胞相對增殖率(Relative Growth Rate,RGR)。RGR(細胞相對增殖率)=實驗組OD 值/對照組OD 值×100%
1.3.5 RT-PCR 檢測3T3-L1 細胞中GLUT-4、HSL的mRNA 表達情況
各組細胞棄培養液,Trizol 法提取3T3-L1 細胞細胞總RNA,鑒定RNA 純度后逆轉錄生成cDNA。PCR 反應條件:預變性95℃60s,變性95℃15s,復性60℃25s,延伸72℃45s,共40 個循環,引物序列,見表1。凝膠成像系統分析,分別計算GLUT-4/β-actin 和HSL/β-actin 的灰度比值。
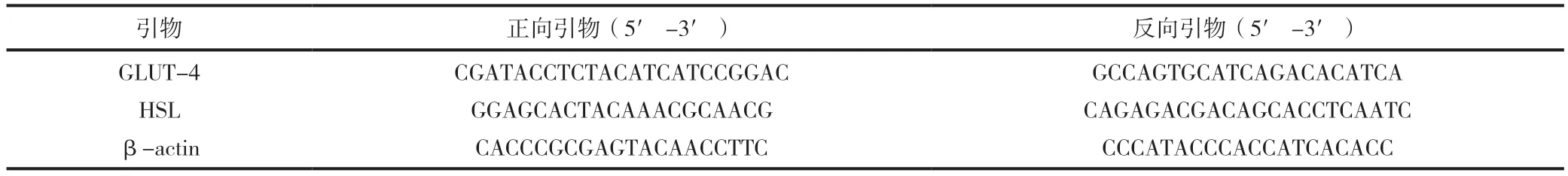
表1 GLUT-4、HSL 的引物序列
1.4 統計學處理
2 結果
2.1 SFPS 對3T3-L1 細胞胰島素抵抗模型葡萄糖消耗量的影響
造模細胞分別培養24h、48h、72h,與對照組比較,培養48h、72h 的模型組細胞培養基中葡萄糖濃度顯著升高(P<0.01),表明細胞對葡萄糖的消耗量顯著降低,見表2。
表2 3T3-L1 細胞培養基中葡萄糖濃度(,n=6,mmol/L)
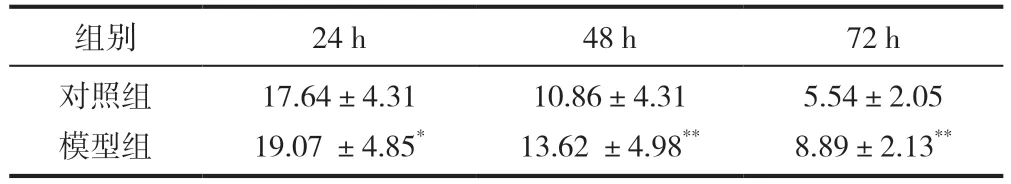
表2 3T3-L1 細胞培養基中葡萄糖濃度(,n=6,mmol/L)
注:與對照組比較,*P<0.05,**P<0.01。
2.2 SFPS 對胰島素抵抗3T3-L1 細胞糖脂代謝的影響
由表3 可知,與模型組比較,SFPS 100mg/L 組和SFPS 200mg/L 組也能顯著增加胰島素抵抗3T3-L1細胞葡萄糖消耗量的量 (P<0.01)。與模型組比較,SFPS 100mg/L 組和SFPS 200mg/L 組也顯著降低胰島素抵抗3T3-L1 細胞生成FFA 的量(P<0.01)。
表3 SFPS 對胰島素抵抗3T3-L1 細胞中糖脂代謝的影響(,n=6)
注:與正常對照組比較,**P<0.01;與模型組比較,##P<0.01。
2.3 SFPS 對胰島素抵抗3T3-L1 細胞相對增殖率的影響
與正常對照組比較,模型組胰島素抵抗3T3-L1細胞24h,48h 和72h 相對增值率降低(P<0.01,P<0.05),24h,48h 細胞相對增殖率降低顯著(P<0.01)。與模型組比較,SFPS 100mg/L 組和SFPS 200mg/L 組能提高胰島素抵抗3T3-L1 細胞相對增值率,提高細胞活力,24h 細胞和48h 細胞相對增值率顯著增加(P<0.01),見表4。
2.4 SFPS 對3T3-L1 細胞中GLUT-4、HSL mRNA表達的影響
與正常對照組比較,模型組胰島素抵抗3T3-L1 細胞GLUT-4 mRNA 表達顯著降低,HSL mRNA 表達顯著增加(P<0.01)(見表5,圖1、2)。與模型組比較,SFPS 100mg/L 組和SFPS 200mg/L 組GLUT-4mRNA 表達顯著增加,HSL mRNA 表達顯著降低(P<0.01,P<0.05)(見表5,圖1、2)。
表4 SFPS 對胰島素抵抗3T3-L1 細胞相對增殖率的影響(,n=6)
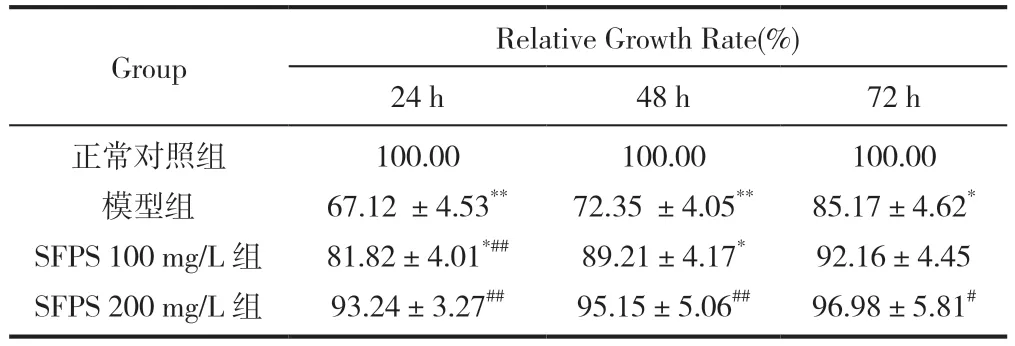
表4 SFPS 對胰島素抵抗3T3-L1 細胞相對增殖率的影響(,n=6)
注:與正常對照組比較,**P<0.01,* P<0.05;與模型組比較,##P<0.01,#P<0.05。
表5 SFPS 對3T3-L1 細胞中GLUT-4、HSL mRNA 表達的影響(,n=6)

表5 SFPS 對3T3-L1 細胞中GLUT-4、HSL mRNA 表達的影響(,n=6)
注:與正常對照組比較,**P<0.01,* P<0.05;與模型組比較,##P<0.01,#P<0.05。
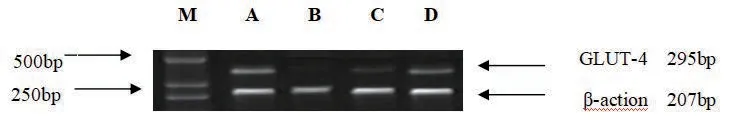
圖1 各組細胞GLUT-4 基因表達

圖2 各組細胞HSL 基因表達
3 討論
2 型糖尿病是一種復雜糖脂代謝紊亂的慢性疾病,以糖耐量異常、脂質代謝異常為特征的胰島素抵抗貫穿于2 型糖尿病的始終。胰島素抵抗是指各種原因引起的外周組織對胰島素的敏感性降低,利用葡萄糖不足[7],主要累及肝臟、脂肪、肌肉等組織,其中脂肪組織分泌的脂肪因子在機制中占重要地位[8-9]。SFPS 是羊棲菜中的多糖成分,具有抗菌、增強免疫力,調節脂代謝等作用[10-11],因此,本實驗進一步研究SFPS 對3T3-L1 細胞胰島素抵抗的作用并初步探討其作用機制。
葡萄糖轉運蛋白(glucose transporter,GLUT)是組織細胞利用調節葡萄糖的重要蛋白,家族重要成員GLUT4 在脂肪、肝臟等組織中均有表達,GLUT4 蛋白將細胞外葡萄糖轉運到細胞內[12-13],對胰島素反應敏感。已有研究顯示,胰島素抵抗的脂肪細胞膜上GLUT4 表達減少,與胰島素抵抗密切相關[14-15]。為了研究SFPS 對3T3-L1 細胞胰島素抵抗的影響,造模成功的細胞培養液中加入不同濃度的SFPS。本研究中,與模型組比較,SFPS 100 和200 mg/L 組細胞中GLUT4mRNA 的表達量顯著增加,表明SFPS 可通過增加GLUT4mRNA 的表達,改善3T3-L1 中胰島素抵抗。激素敏感性脂肪酶(hormone-sensitive triglayceride lipase,HSL)是脂肪分解的限速酶之一,在脂肪組織中受多種激素的調控,主要催化甘油三脂分解為甘油和脂肪酸[16]。為了研究SFPS對3T3-L1 細胞中甘油三酯分解利用的影響,造模成功的細胞培養液中加入不同濃度的SFPS。本研究中,與模型組比較,SFPS 100 和200 mg/L 組細胞中HSL mRNA 的表達量顯著降低,表明SFPS 可通過降低HSL mRNA 的表達,抑制3T3-L1 中脂質的分解利用。
本研究結果顯示:①SFPS 能顯著增加胰島素抵抗3T3-L1 細胞葡萄糖消耗量,顯著降低胰島素抵抗3T3-L1 細胞生成FFA 的量。②SFPS 能顯著增加GLUT-4 mRNA 的表達,顯著降低HSL mRNA 的表達。實驗結果提示,SFPS 改善3T3-L1 脂肪細胞胰島素抵抗的分子機制可能與調控GLUT-4 和HSL mRNA 的表達有關。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
人人健康(2023年26期)2023-12-07 03:55:46
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中國生殖健康(2019年2期)2019-08-23 08:12:10
光學精密工程(2016年6期)2016-11-07 09:07:19
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
核科學與工程(2015年4期)2015-09-26 11:59:03
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
中國醫藥科學(2015年15期)2015-02-27 12:32:27
