轉(zhuǎn)錄因子YY1在肝細(xì)胞癌中的表達(dá)和功能研究
2021-03-26 07:36:34黃貽培羅偉鑫許磊波
嶺南現(xiàn)代臨床外科 2021年1期
黃貽培 ,羅偉鑫 ,許磊波 *
肝細(xì)胞肝癌(Hepatocellular Carcinoma,HCC)是肝臟最常見(jiàn)的原發(fā)惡性腫瘤,具有起病隱匿及容易復(fù)發(fā)轉(zhuǎn)移的特點(diǎn),往往預(yù)后不良[1-3]。目前根治性手術(shù)及手術(shù)替代方案(如靶向治療)均有較大的缺陷和不確定性。弄清HCC 發(fā)生發(fā)展機(jī)制仍然是科研人員不斷追求的目標(biāo)。Yin Yang 1(YY1)是一類含有C2H2 型鋅指結(jié)構(gòu)的轉(zhuǎn)錄因子,屬于GL1-Kruppel 蛋白質(zhì)家族,通過(guò)不同的活性區(qū)域與靶基因啟動(dòng)子結(jié)合而發(fā)揮轉(zhuǎn)錄調(diào)節(jié)功能,參與調(diào)控細(xì)胞干性、分化以及細(xì)胞復(fù)制、增殖等生物學(xué)過(guò)程[4-6]。近來(lái)的研究顯示,YY1 可通過(guò)抑制腫瘤細(xì)胞增殖、侵襲轉(zhuǎn)移或誘導(dǎo)腫瘤細(xì)胞凋亡,在乳腺癌、胰腺導(dǎo)管腺癌及食管鱗狀細(xì)胞癌起抑癌基因的作用[7-11];與此相反,在肺癌、胃癌及結(jié)直腸癌等腫瘤的發(fā)生發(fā)展中則發(fā)揮原癌基因的作用[11-13],表明YY1 在腫瘤進(jìn)展中功能的發(fā)揮具有背景依賴性。YY1 在肝癌中的作用尚存在爭(zhēng)議,Kim JS 等學(xué)者報(bào)道YY1 分布在肝癌細(xì)胞核及細(xì)胞漿中,細(xì)胞核中YY1 高表達(dá)的肝癌患者腫瘤分期更晚,且多合并血管侵犯,患者的生存預(yù)后更差,表明YY1在肝癌中發(fā)揮促癌作用[14];而Wang LM 等的研究則顯示,YY1 在肝癌組織中低表達(dá),并于體外實(shí)驗(yàn)證實(shí)抑制YY1 表達(dá)后肝癌細(xì)胞凋亡減少、侵襲轉(zhuǎn)移能力顯著增強(qiáng),表明YY1在肝癌的進(jìn)展中發(fā)揮抑癌基因作用[15]。因此,YY1 在肝癌的發(fā)生、發(fā)展過(guò)程中的作用亟待更多臨床樣本驗(yàn)證及深入的分子生物學(xué)機(jī)制研究。本研究通過(guò)公共數(shù)據(jù)庫(kù)及臨床組織樣本分析YY1 在肝癌中的預(yù)后意義,同時(shí)探討YY1 對(duì)肝癌細(xì)胞生物學(xué)行為的影響,觀察其對(duì)肝癌細(xì)胞體外增殖、遷移及侵襲能力的影響。
1 材料與方法
1.1 肝癌轉(zhuǎn)錄組測(cè)序數(shù)據(jù)
從腫瘤基因組圖譜計(jì)劃(The Cancer Genome Atlas,TCGA)數(shù)據(jù)庫(kù)中查找并下載肝癌的轉(zhuǎn)錄組測(cè)序數(shù)據(jù)TCGA-LIHC 及相關(guān)臨床參數(shù)進(jìn)行分析,得到374 例肝癌及配對(duì)的50 例癌旁組織的mRNA表達(dá)數(shù)據(jù)。同時(shí)從美國(guó)國(guó)家生物信息中心(National Center for Biotechnology Information,NCBI)創(chuàng)建的基因表達(dá)數(shù)據(jù)庫(kù)(Gene Expression Omnibus,GEO)和國(guó)際癌癥基因組聯(lián)盟數(shù)據(jù)庫(kù)(The International Cancer Genome Consortium,ICGC)中下載肝癌mRNA 片數(shù)據(jù)作為驗(yàn)證隊(duì)列,獲取GSE14520、GSE25097 及LIRI-JP 轉(zhuǎn)錄組測(cè)序數(shù)據(jù)。其中GSE14520 有225 例肝癌及220 例配對(duì)癌旁組織的mRNA 表達(dá)數(shù)據(jù);GSE25097 有 6 例正常肝組織,40 例肝硬化組織,268 例肝癌及243 例配對(duì)癌旁組織的 mRNA 表達(dá)數(shù)據(jù);LIRI-JP 包括有 243 例肝癌及配對(duì)的202 例癌旁組織的mRNA 表達(dá)數(shù)據(jù)。
1.2 肝癌組織樣本
本研究經(jīng)中山大學(xué)孫逸仙紀(jì)念醫(yī)院臨床研究倫理委員會(huì)批準(zhǔn),并獲得受試者的知情同意。本研究采用自 2019 年 9 月~2019 年 12 月在中山大學(xué)孫逸仙紀(jì)念醫(yī)院行手術(shù)切除治療的30 對(duì)肝癌組織及配對(duì)的正常肝組織標(biāo)本,經(jīng)10%福爾馬林溶液固定后用石蠟包埋長(zhǎng)期保存,并制成4 μm 厚組織切片行免疫組化染色,所有患者術(shù)后病理學(xué)報(bào)告證實(shí)為肝細(xì)胞癌。
1.3 肝癌細(xì)胞系及細(xì)胞培養(yǎng)
人肝癌細(xì)胞系 Huh7、PLC/PRF/5、HepG2 和Hep3B 購(gòu)自美國(guó)模式培養(yǎng)物集存庫(kù)(American type culture collection,ATCC),正常肝細(xì)胞系L02、肝癌細(xì)胞系 BEL-7402、MHCC97H、HCCLM3 均購(gòu)自中國(guó)科學(xué)院典型培養(yǎng)物保藏委員會(huì)細(xì)胞庫(kù),在含有10%胎牛血清(Gibco,USA)的DMEM(BI,Israel)培養(yǎng)基中培養(yǎng),常規(guī)添加100 μg/mL 鏈霉素及100 U/mL青霉素(BI,Israel),于5% CO2、37℃恒溫培養(yǎng)箱中培養(yǎng)。
1.4 實(shí)驗(yàn)試劑
YY1 和GAPDH 兔抗人單克隆抗體分別購(gòu)自美國(guó) Abcam 和 Cell Signaling Technology 公司;免疫組化試劑盒(Dako,Denmark);BCA 蛋白定量試劑盒(Invitrogen,USA);反轉(zhuǎn)錄試劑盒及TB Green染料法標(biāo)準(zhǔn)型定量試劑盒(Takara,Japan);Cell Counting Kit-8(CCK-8)檢測(cè)試劑盒(Yeasen,China);Edu 檢測(cè)試劑盒(Beyotime,China)8 μm 孔徑的 Transwell 小室及 Matrigel 購(gòu)自美國(guó) Corning 公司 ;YY1 及 GAPDH 引 物 序 列 為 :GAPDH-F:GGAGCGAGATCCCTCCAAAAT;GAPDH-R:GGCTGTTGTCATACTTCTCATGG;YY1-F:AAGAGCGGCAAGAAGAGTTAC;YY1-R:CAACCACTGTCTCATGGTCAATA。
1.5 免疫組化
肝癌組織石蠟切片經(jīng)60℃烘烤2 h,趁熱置于二甲苯中10 min 進(jìn)行脫蠟處理,使用不同濃度梯度的酒精(無(wú)水酒精,95%,80%,75%)行水化處理,經(jīng)0.3%的過(guò)氧化氫處理10 min 去除內(nèi)源性過(guò)氧化氫酶后,使用Tris-HCl(pH=9.2)抗原修復(fù)液于高壓鍋中修復(fù)10 min,YY1 抗體按1∶1 000 濃度4℃于濕盒孵育過(guò)夜。次日PBS 洗去一抗,37℃溫箱孵育二抗30 min,DAB(DAKO,Denmark)顯色后用蘇木素進(jìn)行細(xì)胞核染色。根據(jù)染色強(qiáng)度,將YY1組織染色強(qiáng)度評(píng)分分為:0=無(wú)染色,1=染色弱,2=染色中等,3=染色強(qiáng),染色范圍按百分比計(jì)算(0~100)。強(qiáng)度評(píng)分乘以染色范圍即為YY1 的染色評(píng)分。
1.6 蛋白免疫印跡實(shí)驗(yàn)
處于對(duì)數(shù)生長(zhǎng)期的細(xì)胞用預(yù)冷的PBS 洗去培養(yǎng)基,甩干后加入預(yù)先配好的細(xì)胞裂解液(RIPA裂解液+1∶100 蛋白酶抑制劑+1∶100 磷酸酶抑制劑,均購(gòu)自中國(guó)康為世紀(jì)公司),冰上裂解15 min,15 000 rpm 離心20 min 后取上清經(jīng)BCA 法測(cè)定蛋白濃度。按細(xì)胞總蛋白量20 μg 進(jìn)行上樣,經(jīng)10%的SDS-PAGE 進(jìn)行蛋白電泳,然后經(jīng)電轉(zhuǎn)至PVDF膜(Millipore,USA)上。加入一抗(YY1 與GAPDH均按照1∶1000 稀釋使用)后置于4℃冰箱過(guò)夜孵育,次日TBST 洗去一抗后,抗兔二抗(Cell Signaling Technology,USA)室溫孵育1 h。BCL 超敏發(fā)光液(Millipore,USA)曝光并拍攝。
1.7 實(shí)時(shí)熒光定量PCR
按照試劑說(shuō)明書使用RNAiso Plus(Invitrogen,USA)提取細(xì)胞RNA,采用PrimerScript RT Master Mix 試劑盒(Takara,Japan)進(jìn)行逆轉(zhuǎn)錄得到cDNA。實(shí)時(shí)定量PCR 實(shí)驗(yàn)按照TB Green Premix Ex Taq 試劑盒說(shuō)明書(Takara,Japan)進(jìn)行,操作儀器為Roche LightCycler 480Ⅱ。PCR 實(shí)驗(yàn)以 GAPDH 作為內(nèi)源性對(duì)照,每個(gè)反應(yīng)的溶解曲線只有一個(gè)峰才可視為有效結(jié)果。實(shí)驗(yàn)重復(fù)3 次。
1.8 CCK-8 法檢測(cè)細(xì)胞增殖
處于對(duì)數(shù)生長(zhǎng)期的細(xì)胞,經(jīng)胰酶消化及離心后,用完全培養(yǎng)基重懸成單細(xì)胞懸液,計(jì)數(shù)后將細(xì)胞接種至96 孔板,密度為1000 個(gè)/孔,每孔完全培養(yǎng)基為100 μL。置于細(xì)胞培養(yǎng)箱中培養(yǎng)。待細(xì)胞貼壁后,沿孔壁緩慢加入10 μL CCK-8 試劑,輕微搖勻后置于細(xì)胞培養(yǎng)箱中,待孵育1 h 后使用酶標(biāo)儀在450 nm 處檢測(cè)每個(gè)孔的吸光值。同樣的方法檢測(cè)第24 h、48 h、72 h、96 h 的吸光值。實(shí)驗(yàn)重復(fù)3 次。
1.9 Edu 法檢測(cè)細(xì)胞增殖
按3000 個(gè)/孔的密度將細(xì)胞接種于96 孔板中,置于細(xì)胞培養(yǎng)箱中培養(yǎng)過(guò)夜。按照試劑盒說(shuō)明書加入預(yù)先配置好的1X Edu 溶液,細(xì)胞培養(yǎng)箱中孵育2 h,棄去Edu 溶液,4%多聚甲醛固定15 min,用0.3%TritonX-100/PBS 溶液室溫孵育15 min 進(jìn)行膜通透處理,用Hoechst 給細(xì)胞核染色后于熒光顯微鏡下拍照保存、計(jì)數(shù)。實(shí)驗(yàn)重復(fù)3 次。
1.10 克隆形成實(shí)驗(yàn)
按每孔1000 個(gè)細(xì)胞接種于6 孔板中,混勻后置于細(xì)胞培養(yǎng)箱中,培養(yǎng)箱溫度保持37℃,二氧化碳體積濃度比維持5%,每2~3 d 給細(xì)胞換液。待細(xì)胞長(zhǎng)出明顯的單細(xì)胞克隆后,吸去孔中培養(yǎng)基,PBS 清洗后加入4%多聚甲醛固定15 min,加入0.1%結(jié)晶紫溶液給細(xì)胞染色30 min,用ddH2O 洗去殘留的結(jié)晶紫溶液,晾干后拍照、計(jì)數(shù)。實(shí)驗(yàn)重復(fù)3 次。
1.11 Transwell 細(xì)胞遷移侵襲實(shí)驗(yàn)
基質(zhì)膠提前一晚放于冰上化凍,用預(yù)冷的無(wú)血清DMEM 培養(yǎng)基按1∶3 的比例稀釋基質(zhì)膠,將80 μL 稀釋后的凝膠滴入 8 μm 孔徑的 Transwell 小室上室面,使凝膠鋪勻后,室溫靜置2 min 后吸去60 μL,上室面留下 20 μL 稀釋后的基質(zhì)膠,放入細(xì)胞培養(yǎng)箱2 h 待其凝固備用。將細(xì)胞消化離心,用PBS 洗去殘留的培養(yǎng)基,再次離心后用無(wú)血清的DMEM 重懸計(jì)數(shù),調(diào)整細(xì)胞密度為5×105個(gè)/mL,分別取 200 μL 加入上室,下室加入 700 μL 含 10%血清的DMEM 完全培養(yǎng)基,置于細(xì)胞培養(yǎng)箱中培養(yǎng)24 h。取出Transwell 小室,棄去上室中培養(yǎng)液,PBS 清洗后置于4%多聚甲醛中固定15 min,PBS 清洗后置于0.1%濃度的結(jié)晶紫中染色30 min,ddH2O洗去結(jié)晶紫,晾干后于顯微鏡下拍照計(jì)數(shù)。遷移實(shí)驗(yàn)不鋪基質(zhì)膠,上室加入100 μL 細(xì)胞懸液,余實(shí)驗(yàn)步驟相同。實(shí)驗(yàn)重復(fù)3 次。
1.12 基因集富集分析
將TCGA 數(shù)據(jù)庫(kù)中的肝癌mRNA 表達(dá)數(shù)據(jù)根據(jù)YY1 表達(dá)高低進(jìn)行分組,YY1 表達(dá)高于總體75%者歸于YY1 高表達(dá)組,表達(dá)低于總體25%者歸于YY1低表達(dá)組,兩組間的基因表達(dá)矩陣用于基因集富集分析(gene set enrichment analysis,GSEA)[16],選用c2.cp.kegg.v6.0.symbols.gmt 作為預(yù)定義基因集,分析與YY1 表達(dá)最相關(guān)的生物過(guò)程及信號(hào)通路。
1.13 基因富集分析
利用R 軟件包“clusterProfiler”對(duì)篩選出來(lái)的93 個(gè)YY1 共表達(dá)基因進(jìn)行KEGG 富集分析和GO富集分析[17],分析共表達(dá)基因的生物學(xué)功能。繪制柱狀圖:橫坐標(biāo)為基因數(shù),縱坐標(biāo)為各信號(hào)通路,柱狀圖顏色代表p 值。
1.14 蛋白質(zhì)互作網(wǎng)絡(luò)構(gòu)建
利用String 數(shù)據(jù)庫(kù)(https://string-db.org/)對(duì)93個(gè)YY1 共表達(dá)基因構(gòu)建蛋白質(zhì)互作網(wǎng)絡(luò)(protein -protein interaction network,PPI)[18],置信度設(shè)置為0.7(高度可信),所得結(jié)果用Cytoscape 軟件行可視化分析。
1.15 統(tǒng)計(jì)學(xué)分析
本研究采用GraphPad Prism 8.0 及R 3.6.1 軟件對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)學(xué)分析。生存分析使用Kaplan-Meier法。體外細(xì)胞功能學(xué)實(shí)驗(yàn)均重復(fù)3次,所有實(shí)驗(yàn)數(shù)據(jù)顯示為3 次獨(dú)立實(shí)驗(yàn)的均數(shù)±標(biāo)準(zhǔn)差。兩樣本之間比較,數(shù)據(jù)呈正態(tài)分布且方差齊采用Studentt 檢驗(yàn);方差不齊的數(shù)據(jù)采用校正t檢驗(yàn)。P<0.05 時(shí)差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié) 果
2.1 YY1 在肝癌中表達(dá)上調(diào)且與不良預(yù)后相關(guān)
在TCGA 數(shù)據(jù)庫(kù)的肝癌隊(duì)列中發(fā)現(xiàn),YY1 在肝癌中表達(dá)量顯著上調(diào)(圖1A),高表達(dá)YY1 的患者總體生存期(overall survival,OS)較短(圖1B)。其中,低表達(dá)YY1 的肝癌患者中位生存期為5.84 年,而高表達(dá)YY1 的肝癌患者中位生存期僅為3.14年。進(jìn)一步分析發(fā)現(xiàn),在腫瘤體積分期為T3 及T4的患者中YY1 表達(dá)更高(10.36±3.64 vs 9.04±2.75,P<0.01,圖1C);腫瘤分期越晚(Ⅲ+Ⅳ)、組織學(xué)分化更差(Ⅲ+Ⅳ)的患者中YY1 表達(dá)顯著增加(圖1D-1E,P<0.05),提示YY1 表達(dá)可能與肝癌增殖、惡性進(jìn)展相關(guān)。在GSE14520、GSE25097 及ICGCLIRI-JP 肝癌驗(yàn)證隊(duì)列中,我們發(fā)現(xiàn)YY1 在肝癌組織中表達(dá)量顯著高于癌旁組織,差異均有統(tǒng)計(jì)學(xué)意義(P<0.0001,圖1F~1H)
2.2 YY1 在肝癌組織及細(xì)胞系中的表達(dá)情況
分析我院30 對(duì)肝癌及配對(duì)癌旁組織的YY1免疫組化染色發(fā)現(xiàn),YY1 集中表達(dá)在細(xì)胞核中,相比于癌旁組織,YY1 在肝癌組織內(nèi)陽(yáng)性區(qū)域更多、染色更強(qiáng)(圖2A),肝癌組織的免疫評(píng)分顯著高于癌旁組織(P<0.05,圖2B)。通過(guò)蛋白質(zhì)免疫印跡實(shí)驗(yàn),我們發(fā)現(xiàn)YY1 在BEL-7402、HepG2、Hep3B、Huh7、MHCC97H、HCCLM3 等肝癌細(xì)胞系中高表達(dá),在PLC/PRF/5 中低表達(dá)(圖2C)。因此,我們使用敲降表達(dá)及過(guò)表達(dá)YY1 的慢病毒載體構(gòu)建穩(wěn)定轉(zhuǎn)染的肝癌細(xì)胞株;其中,肝癌細(xì)胞Huh7 敲降 YY1 表達(dá),低表達(dá) YY1 的 PLC/PRF/5 細(xì)胞則過(guò)表達(dá) YY1。如圖 2D 中顯示,Huh7 敲低 YY1 表達(dá)后,蛋白水平明顯下降;PLC/PRF/5 經(jīng)過(guò)表達(dá)YY1后,蛋白水平也明顯增加。實(shí)時(shí)熒光定量PCR 實(shí)驗(yàn)證實(shí),在Huh7 中敲低YY1 表達(dá)后mRNA 水平明顯下降(P<0.05,圖 2E);PLC/PRF/5 中過(guò)表達(dá)YY1 組較對(duì)照組mRNA 的表達(dá)明顯上調(diào)(P<0.05,圖2F)。
2.3 YY1 對(duì)肝癌細(xì)胞體外增殖功能的影響
在TCGA 數(shù)據(jù)庫(kù)肝癌研究隊(duì)列中,YY1 的表達(dá)與患者腫瘤大小顯著相關(guān)(圖1C)。進(jìn)一步研究發(fā)現(xiàn),YY1 的表達(dá)水平與常見(jiàn)的增殖相關(guān)基因MKI67、PCNA 及MCM2 表達(dá)水平呈顯著正相關(guān)(Spearman相關(guān)系數(shù)分別為:Ki67:R=0.46,P<0.0001;PCNA:R=0.44,P<0.0001;MCM2:R=0.46,P<0.0001;圖3A-3C)。體外CCK-8 檢測(cè)實(shí)驗(yàn)顯示,在Huh7 中干擾YY1 表達(dá)后,細(xì)胞增殖減慢,在第 96 h 的 OD450 顯著低于對(duì)照組(1.040±0.092vs1.447±0.099,P<0.01,圖 3D);PLC/PRF/5 過(guò)表達(dá)YY1 后細(xì)胞增殖顯著加快,PLC/PRF/5-YY1 在第96 h 的吸光值顯著高于對(duì)照組(1.266±0.053vs0.963±0.041,P<0.001,圖3F)。在 Edu 增殖檢測(cè)實(shí)驗(yàn)中,Huh7-shYY1 組較對(duì)照Huh7-shCon 組Edu 陽(yáng)性的細(xì)胞比例明顯減少(P<0.001,圖3E),提示細(xì)胞增殖比例下降;在PLC/PRF/5-YY1 相比于對(duì)照組細(xì)胞,Edu 陽(yáng)性細(xì)胞占比則有所增加(P<0.01,圖3G)。克隆形成實(shí)驗(yàn)中發(fā)現(xiàn),Huh7 干擾YY1 表達(dá)后,其克隆形成數(shù)目(392±55.6)少于對(duì)照組(151.3±66.9),差異有統(tǒng)計(jì)學(xué)意義(P<0.01,圖3H)。而過(guò)表達(dá)YY1 的PLC/PRF/5 細(xì)胞克隆形成數(shù)目(446.7±39.5)顯著多于對(duì)照組(306±41,P<0.05,圖3I)。以上實(shí)驗(yàn)結(jié)果提示YY1 可影響肝癌細(xì)胞的增殖能力,提示YY1 可能在促進(jìn)肝癌增殖、生長(zhǎng)中發(fā)揮了重要的作用。
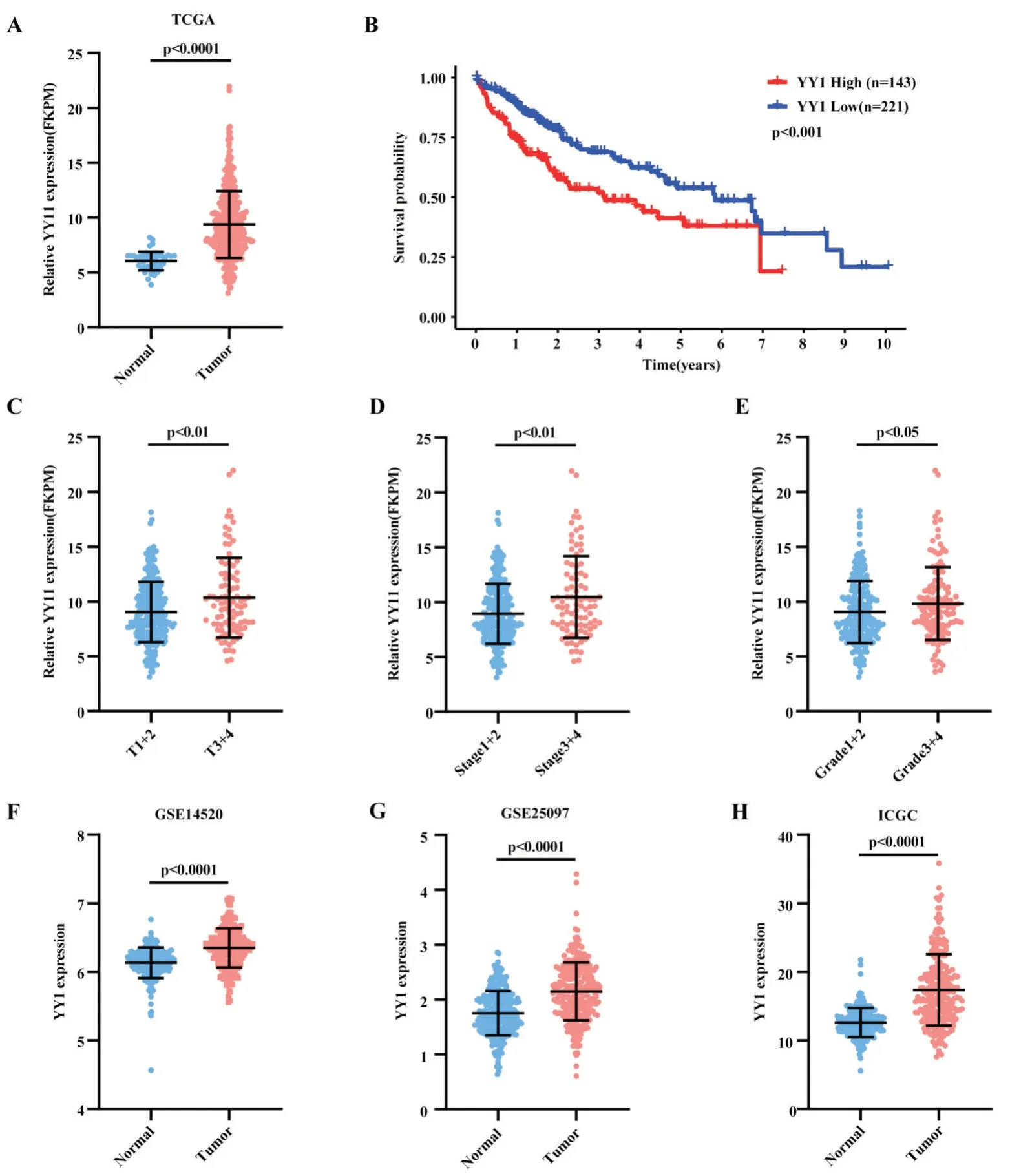
圖1 YY1 在肝癌組織中的表達(dá)情況圖 A:YY1 在TCGA-LIHC 中的表達(dá)情況(t 檢驗(yàn));B:TCGA-LIHC 中YY1 表達(dá)與患者總體生存期相關(guān)(Kaplan-Meier 法,log-rank 檢驗(yàn));C、D、E:YY1 在TCGA-LIHC 中與腫瘤體積、腫瘤分期、病理學(xué)分級(jí)相關(guān)(t 檢驗(yàn));F:YY1 在GSE14520 中的表達(dá)情況(t 檢驗(yàn));G:YY1 在GSE25097 中的表達(dá)情況(t 檢驗(yàn));H:YY1 在ICGC-LIRI-JP 中的表達(dá)情況(t 檢驗(yàn))
2.4 YY1 對(duì)肝癌細(xì)胞體外遷移侵襲能力的影響
Transwell 細(xì)胞遷移、侵襲實(shí)驗(yàn)顯示,Huh7 干擾YY1 表達(dá)后,Huh7-shYY1 組 24 h 的細(xì)胞遷移數(shù)(62.7±11.7)較對(duì)照組(118.3±7.6)明顯減少;侵襲細(xì)胞數(shù) Huh7-shYY1 組為 49±2,對(duì)照組為 100.7±10.5,遷移、侵襲細(xì)胞數(shù)兩組差異均有統(tǒng)計(jì)學(xué)意義(P<0.05,圖 4A)。過(guò)表達(dá) YY1 的 PLC/PRF/5 細(xì)胞中,24 h 遷移及侵襲細(xì)胞數(shù)較對(duì)照組均明顯增加(遷移細(xì)胞數(shù):220.7±14.3vs59.7±13;侵襲細(xì)胞數(shù):176.3±15.5vs38±8),兩組差異均有統(tǒng)計(jì)學(xué)意義(P<0.05,圖4B)。因此,體外實(shí)驗(yàn)證實(shí)YY1 可以促進(jìn)肝癌細(xì)胞的侵襲轉(zhuǎn)移能力。
2.5 YY1 與基因集富集分析
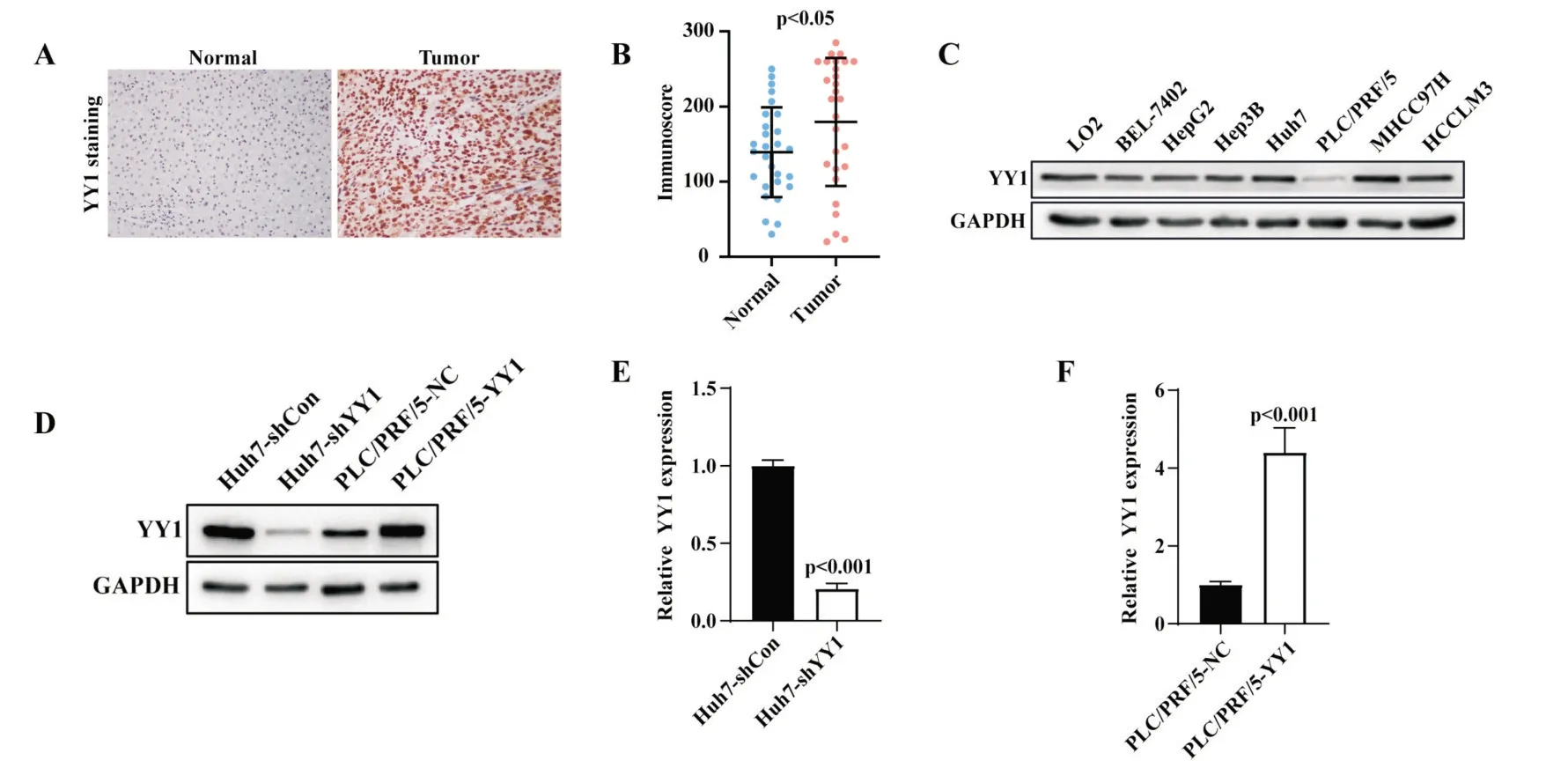
圖2 YY1 在肝癌組織及肝癌細(xì)胞系中的表達(dá)情況 A:YY1 在肝癌組織及配對(duì)癌旁組織的免疫組化染色;B:YY1 在30 對(duì)肝癌組織及配對(duì)癌旁組織的免疫評(píng)分(t 檢驗(yàn));C:肝癌細(xì)胞系中YY1 的本底表達(dá)情況;D:蛋白免疫印跡實(shí)驗(yàn)檢測(cè)YY1 敲降表達(dá)及過(guò)表達(dá)效果;E:q-PCR 實(shí)驗(yàn)檢測(cè)YY1 敲降表達(dá)及過(guò)表達(dá)效果(t 檢驗(yàn))
基因集富集分析比較TCGA-LIHC 中高、低表達(dá)YY1 兩組之間差異的信號(hào)通路,我們發(fā)現(xiàn)YY1高表達(dá)組中富集了多種腫瘤相關(guān)的通路,比如KEGG-PATHWAYS-IN-CANCER、KEGG-CELL-CYCLE、KEGG-APOPTOSIS 和 KEGG-VEGF-SIGNALING-PATHWAY。同時(shí),參與腫瘤發(fā)生及進(jìn)展相關(guān)的分子信號(hào)通路,比如KEGG-ERBB-SIGNALINGPATHWAY、KEGG-MAPK-SIGNALING-PATHWAY、KEGG-NOTCH-SIGNALING-PATHWAY 及KEGGWNT-SIGNALING-PATHWAY也顯著富集于YY1高表達(dá)組中(圖5A,P<0.05,F(xiàn)DR<0.05)。這些結(jié)果提示YY1 的表達(dá)上調(diào)可能會(huì)激活這些信號(hào)通路從而促進(jìn)肝癌進(jìn)展。
2.6 YY1 與基因共表達(dá)分析
具有類似表達(dá)模式的基因可能參與相近的分子生物學(xué)功能,因此,我們篩選TCGA、GEO 及ICGC 數(shù)據(jù)庫(kù)中與YY1 表達(dá)顯著相關(guān)的基因,我們將與YY1 表達(dá)相關(guān)系數(shù)絕對(duì)值超過(guò)0.4,p 值小于0.05 的基因定義為YY1 的共表達(dá)基因,四個(gè)不同的肝癌研究隊(duì)列中篩選出93 個(gè)共表達(dá)基因(圖5B)。經(jīng)過(guò)KEGG 通路富集分析,我們發(fā)現(xiàn)這些共表達(dá)基因主要參與了基因復(fù)制及基因轉(zhuǎn)錄相關(guān)的信號(hào)通路,比如DNA 復(fù)制、RNA 轉(zhuǎn)運(yùn)及細(xì)胞周期等(圖5C)。在GO 通路富集分析中,這些共表達(dá)基因參與了P53基因介導(dǎo)的信號(hào)轉(zhuǎn)導(dǎo),tRNA轉(zhuǎn)運(yùn)及DNA 復(fù)制等(圖5D)。進(jìn)一步分析中,我們通過(guò)String 數(shù)據(jù)庫(kù)構(gòu)建YY1 與這些共表達(dá)基因的蛋白質(zhì)互作網(wǎng)絡(luò),在置信度為0.7(高度可信)時(shí),我們發(fā)現(xiàn)93 個(gè)共表達(dá)基因之間的蛋白互作網(wǎng)絡(luò)數(shù)(249)比預(yù)期的互作網(wǎng)絡(luò)數(shù)(60)顯著增加(PPI enrichmentPvalue=1.0×10-16)。其中預(yù)測(cè)到 YY1 與8 個(gè)蛋白存在高度可信的互作網(wǎng)絡(luò),包括FKBP3、RBBP7、H2AFV、ACTL6A、PAXIL1、NCOA6、SUZ12和H2AFZ,提示YY1可能與這8個(gè)共表達(dá)基因存在相互作用關(guān)系,參與調(diào)控相應(yīng)的分子生物學(xué)功能。
3 討 論
YY1 是1991 年被發(fā)現(xiàn)并純化的鋅指類轉(zhuǎn)錄因子,屬于GL1-Kruppel 蛋白質(zhì)家族,編碼基因位于人類14 號(hào)染色體端粒區(qū)q32.2,序列在脊椎動(dòng)物中高度保守[19,20]。作為一類重要的轉(zhuǎn)錄因子,YY1可以影響大約7%的哺乳動(dòng)物基因的轉(zhuǎn)錄活性,通過(guò)調(diào)控基因轉(zhuǎn)錄、大規(guī)模染色體重組及X 染色體失活等參與調(diào)節(jié)細(xì)胞增殖、分化和凋亡等生物學(xué)過(guò)程[21]。多項(xiàng)研究表明YY1 在多種腫瘤中異常表達(dá),發(fā)揮抑癌或促癌作用。如在乳腺癌、胰腺導(dǎo)管腺癌及食管鱗狀細(xì)胞癌中,YY1 能抑制腫瘤進(jìn)展[7-10],而在肺癌、胃癌及結(jié)直腸癌中則發(fā)揮促癌作用[11-13]。在肝癌的研究中,Kim 等人報(bào)道 YY1 在細(xì)胞核中的表達(dá)上調(diào)與肝癌不良預(yù)后相關(guān)[14],Tsang 等人分析50 對(duì)肝癌與配對(duì)癌旁組織免疫組化染色發(fā)現(xiàn),YY1 在68%的肝癌組織中表達(dá)量明顯增加[22]。而在Wang LM 等人的報(bào)道中,他們分析了80 對(duì)肝癌組織及配對(duì)癌旁組織的mRNA 表達(dá)水平,發(fā)現(xiàn)65%肝癌組織中YY1 表達(dá)低于癌旁組織,而且高表達(dá)YY1 的患者肝癌組織病理學(xué)分級(jí)較好,指出YY1高表達(dá)在肝癌中是保護(hù)因素而非危險(xiǎn)因素[15]。因而不能將YY1 籠統(tǒng)地歸為抑癌基因或癌基因,YY1 在不同腫瘤甚至同一腫瘤中功能的差異性可能與其特殊的結(jié)構(gòu)有關(guān),YY1 蛋白包含多個(gè)不同的活性催化區(qū)域一方面可直接識(shí)別結(jié)合啟動(dòng)子區(qū)的YY1 應(yīng)答元件促進(jìn)靶基因轉(zhuǎn)錄,另一方面還可以通過(guò)招募共激活因子(如p300、CBP或PRMT1 等)或共抑制因子(如HDACs、EZH2 或DNMTs)改變下游基因YY1 應(yīng)答元件附近染色質(zhì)的表觀遺傳修飾狀態(tài),促進(jìn)或抑制下游基因的表達(dá)[23,24]。
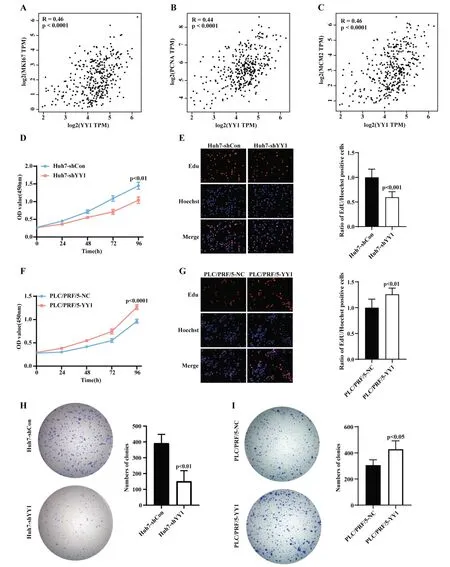
圖3 YY1 對(duì)肝癌細(xì)胞增殖和克隆形成能力的影響 A、B、C:YY1 在TCGA-LIHC 中與增殖相關(guān)基因MKI67、PCNA 和MCM2 顯著正相關(guān)(Spearman 相關(guān)性分析);D、F:CCK-8 實(shí)驗(yàn)檢測(cè)敲降或過(guò)表達(dá)YY1 后細(xì)胞增殖情況(t 檢驗(yàn));E、G:Edu 染色法檢測(cè)敲降或過(guò)表達(dá)YY1后細(xì)胞增殖情況(t 檢驗(yàn));H、I:敲降或過(guò)表達(dá)YY1 后細(xì)胞克隆形成能力改變情況(t 檢驗(yàn))
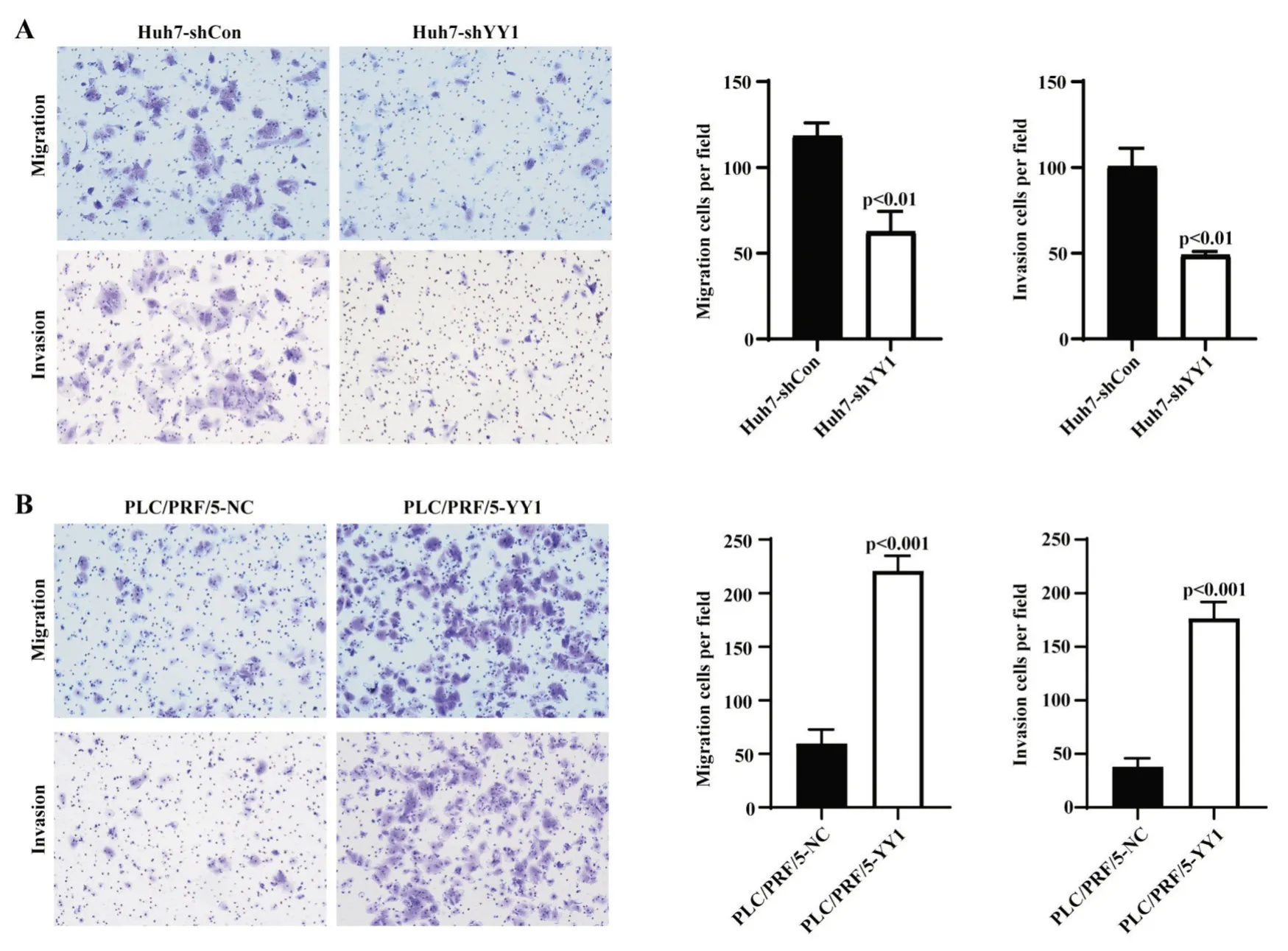
圖4 YY1 對(duì)肝癌細(xì)胞遷移、侵襲能力的影響 A:敲降YY1 表達(dá)后Huh7 遷移、侵襲能力顯著降低(t 檢驗(yàn));B:過(guò)表達(dá)YY1 后PLC/PRF/5 遷移、侵襲能力顯著增強(qiáng)(t 檢驗(yàn))
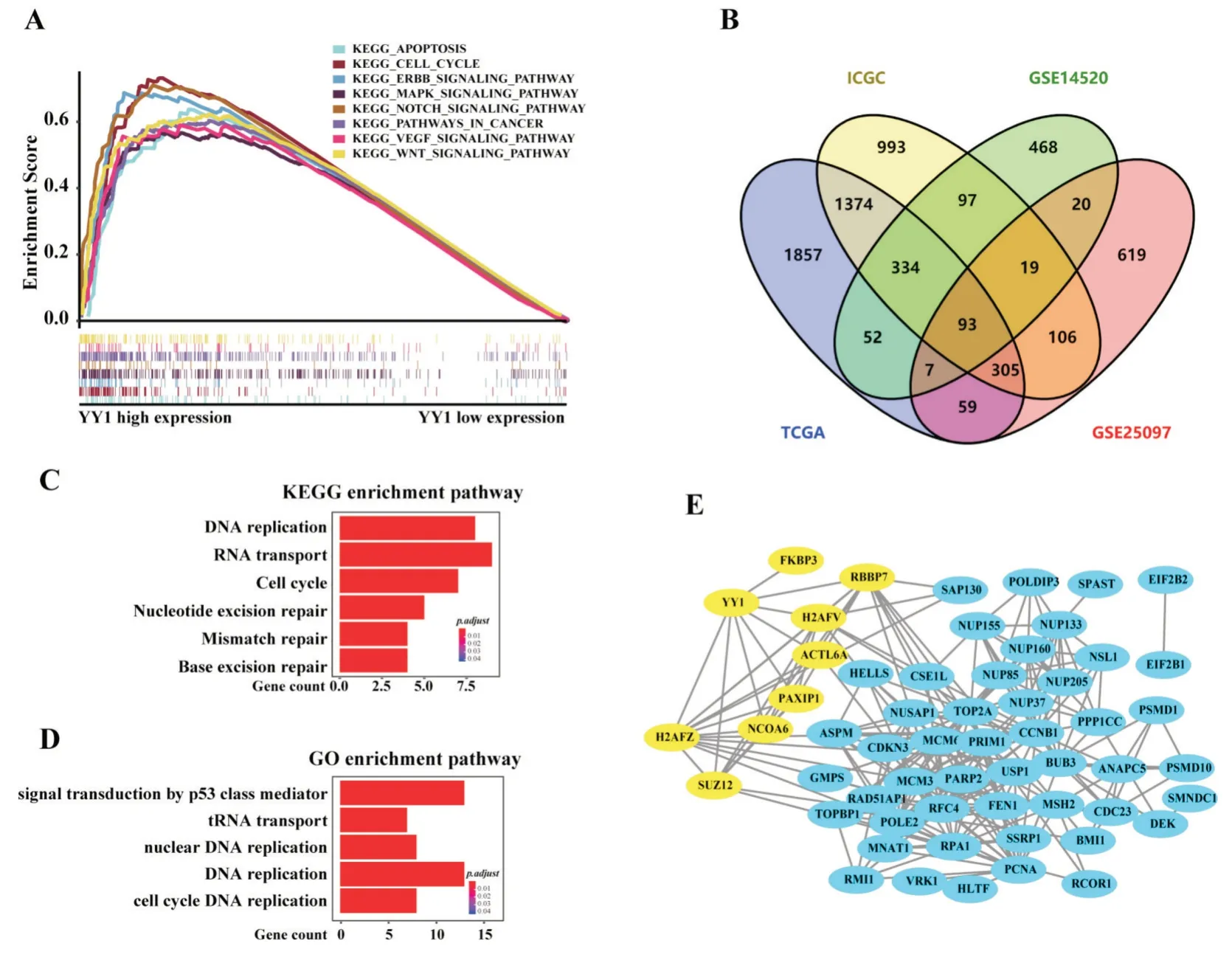
圖5 基因集富集分析、基因共表達(dá)分析及蛋白質(zhì)互作網(wǎng)絡(luò)構(gòu)建 A:YY1 高表達(dá)的HCC 患者明顯富集多種與腫瘤發(fā)生發(fā)展相關(guān)的信號(hào)通路;B:篩選并確定93 個(gè)TCGA-LIHC、GSE14520、GSE25097 及ICGC-LIRI-JP 數(shù)據(jù)集中的YY1 過(guò)表達(dá)基因;C、D:93 個(gè)YY1 共表達(dá)基因的KEGG、GO 通路富集分析;E:構(gòu)建YY1 與共表達(dá)基因之間的蛋白質(zhì)互作網(wǎng)絡(luò)
本研究分析TCGA-LIHC、GSE14520、GSE25097及ICGC-LIRI-JP 四個(gè)肝細(xì)胞癌研究隊(duì)列中YY1 的表達(dá)及臨床意義,并在本院收集的樣本中進(jìn)行驗(yàn)證,分析結(jié)果均顯示YY1 在肝癌組織中的表達(dá)顯著高于正常肝組織,且YY1 高表達(dá)能指示患者的不良預(yù)后。值得注意的是YY1 與腫瘤體積、腫瘤分期、腫瘤病理學(xué)分級(jí)密切相關(guān),提示YY1 可能在多方面影響腫瘤進(jìn)展。我們的體外實(shí)驗(yàn)結(jié)果顯示,YY1 可以促進(jìn)肝癌細(xì)胞增殖、遷移及侵襲,進(jìn)一步證實(shí)YY1 確實(shí)可以從多個(gè)方面影響到肝癌細(xì)胞。在探討YY1 在肝癌中發(fā)揮作用的潛在機(jī)制時(shí),我們分析了TCGA-LIHC 研究隊(duì)列的轉(zhuǎn)錄組測(cè)序數(shù)據(jù),與預(yù)期一致,結(jié)果顯示YY1 高表達(dá)組富集到更多與腫瘤發(fā)生及進(jìn)展相關(guān)的信號(hào)通路。此外,分析YY1 的共表達(dá)基因并構(gòu)建蛋白質(zhì)互作網(wǎng)絡(luò)發(fā)現(xiàn),YY1 與這些共表達(dá)基因參與DNA 復(fù)制、RNA 轉(zhuǎn)運(yùn)及細(xì)胞周期等生物學(xué)過(guò)程,進(jìn)一步提示轉(zhuǎn)錄因子YY1 在肝癌中可能廣泛參與調(diào)控基因復(fù)制及轉(zhuǎn)錄;預(yù)測(cè)到與YY1 存在高度可信的蛋白質(zhì)互作網(wǎng)絡(luò)的基因可能是YY1 的下游靶基因,這可以為我們后續(xù)的機(jī)制研究提供參考和方向。
綜上所述,本研究初步探討了YY1 基因在肝癌中的預(yù)后意義,體外實(shí)驗(yàn)證明YY1 可以促進(jìn)肝癌細(xì)胞的增殖及惡性轉(zhuǎn)化能力,靶向干擾肝癌組織中的YY1 表達(dá)可能是潛在的藥物治療靶點(diǎn)。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
天津醫(yī)科大學(xué)學(xué)報(bào)(2019年3期)2019-08-13 06:53:08
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
腫瘤預(yù)防與治療(2015年1期)2015-09-26 07:26:20
中國(guó)當(dāng)代醫(yī)藥(2015年16期)2015-03-01 02:03:11
