玉木耳絡氨酸酶基因的克隆及表達分析
2021-03-23 08:38:32蘇文英楊和川譚一羅惠林沖秦裕營
江西農業學報 2021年3期
關鍵詞:分析
蘇文英,楊和川,譚一羅,惠林沖,秦裕營,李 曉
(1.連云港市農業科學院,江蘇 連云港 222006;2.吉林農業大學 食藥用菌教育部工程中心,吉林 長春 130118)
酪氨酸酶(Tyrosinase)是一種含銅糖蛋白,一般也稱為多酚氧化酶,歸類于酪氨酸酶相關蛋白家族(Tyrosinase-related Protein)[1]。酪氨酸酶廣泛存在于微生物、動植物及人體中,并發揮著重要的生理功能[2]。在真菌中,雙孢蘑菇的酪氨酸酶較早受到關注,在20世紀70年代,Turner 等就提出酪氨酸酶可能在雙孢蘑菇(Agaricusbisporus)黑色素的合成中發揮相當重要的作用[3];絡氨酸酶在氧氣的參與下,將酪氨酸氧化為多酚類化合物(如L-多巴)及多巴醌,多巴醌在一系列反應后最終生成黑色素[4]。2016年賓夕法尼亞大學的楊亦農教授利用基因編輯技術對雙孢蘑菇中控制褐變的多酚氧化酶基因進行了基因編輯,并將該酶的活性降低了30%,使其抗褐變[5]。此外,真菌絡氨酸酶還與孢子的形成及穩定性相關。有研究表明,絡氨酸酶在真菌菌絲無性生長時期發揮重要的作用,其表達量在菌絲生長期間逐漸上升,至子實體形成時開始下降[6]。目前,香菇(Lentinusedodes)[7]、漏斗多孔菌(Polyporusarcularius)[8]等真菌中的絡氨酸酶基因已被克隆鑒定,為其在真菌內的表達模式分析及功能研究提供了方便。
玉木耳(AuriculariacorneaEhrenb.),隸屬于木耳科木耳屬,是毛木耳的白色變種[9],其質地脆嫩,美味可口,營養豐富,且具有抗腫瘤等活性[10-11]。目前關于玉木耳的研究集中在栽培、生理活性及成分分析方面,對其分子遺傳學的研究還鮮有報道。鑒于此,我們在克隆玉木耳絡氨酸酶基因(Actyr)的基礎上,還測定了玉木耳菌絲由營養生長到生殖生長整個過程中關鍵時期絡氨酸酶基因的相對表達量,從而對所有絡氨酸酶基因的功能進行了系統分析,以期為深入研究絡氨酸酶基因在玉木耳子實體發育過程中的生物學功能提供科學依據。
1 材料與方法
1.1 供試菌株
玉木耳菌絲材料由吉林農業大學食藥用菌教育部工程研究中心提供;大腸桿菌(E.coli)DH5α購自北京全式金生物技術有限公司。
1.2 試劑
UNlQ-10柱式Trizol總RNA抽提試劑盒(生工公司,上海); Phanta Max Super-Fidelity DNA Polymerase(諾唯贊生物科技股份有限公司,南京); SYBR PrimeScript TM RT-PCR Kit熒光定量試劑盒[寶生物工程(大連)有限公司,大連]; Peasy-T1 cloning vector(北京全式金生物技術有限公司,北京); AxyPrep DNA凝膠回收試劑盒(愛思進生物技術有限公司,杭州)。
1.3 試驗方法
1.3.1 玉木耳總RNA的提取及cDNA第一鏈的合成 用總RNA提取試劑盒提取玉木耳的RNA,具體操作參照說明書進行。采用凝膠電泳及微量測定儀NanoDrop ND-1000對提取獲得的RNA的濃度和純度進行檢測。在檢測合格后于-80 ℃保存備用。根據試劑盒說明書進行反轉錄,所得cDNA于-20 ℃冰箱中保存備用。
1.3.2 玉木耳Actyr基因全長序列的克隆 以逆轉錄獲得的cDNA為模板,利用RT-PCR技術擴增得到Actyr基因的全長cDNA序列。PCR引物序列如下:Actyr-F, ATGCCACAAGGTCACAGG;Actyr-R, TCAATCGTACTCGTAGCAC。引物由金唯智生物技術有限責任公司合成。PCR反應體系為: ddH2O 17 μL、2 × Phanta Max Buffer 25 μL、dNTP Mix (10 mmol/L each) 1 μL、模板cDNA 2 μL、上下游引物各2 μL、Phanta Max Super-Fidelity DNA Polymerase (1 μg/μL) 1 μL。PCR擴增反應條件為:95 ℃預變性3 min;95 ℃變性30 s,55 ℃退火15 s,72 ℃延伸1 min,共30個循環。在PCR反應結束后,將PCR產物經1%瓊脂糖凝膠電泳檢測正確后,利用凝膠回收試劑盒回收PCR產物,具體操作參照試劑盒說明書。
1.3.3 擴增產物的克隆及測序 將回收純化的產物連接T-vector,連接反應體系為: Peasy-T1 cloning vector 1 μL、回收產物4 μL,反應條件為25 ℃ 20 min。獲得的連接產物直接用于大腸桿菌轉化,轉化方法為熱激法。將轉化產物涂布于平板,在37 ℃過夜培養;挑取單菌落于5 mL LB(含Amp+)液體培養基中,在200 r/min、37 ℃條件下震蕩培養過夜。對重組克隆載體進行菌液PCR鑒定。委托生工公司進行菌液PCR測序。
1.3.4 基因序列的分析 根據克隆獲得的Actyr的核酸序列以及對應的氨基酸序列,通過在線分析軟件在理論上對其進行基因信息分析。
利用DNAMAN軟件對測序所得序列與Actyr基因序列進行序列同源性分析;通過NCBI Conserved Domains進行保守功能域分析;應用PredictProtein軟件對Actyr蛋白質的二級結構進行分析;應用Protparam (http://expasy.org/tools/protparam.htmL)對其進行分子量和等電點的預測;應用TMpred軟件(http://www.isrec.isb-sib.ch/software/tmpred_from.html)分析預測蛋白序列的跨膜區;應用SignalP 4. 1 (http://www.cbs.dtu.dk/services/SignalP/)預測信號肽;應用Swiss-MODEL在線工具軟件對蛋白質的三級結構進行成像分析。
1.3.5 基因表達量的測定 收集耳芽期、原基期、耳片期(耳片直徑2、3、4 cm)5個時期的材料,提取RNA,反轉錄后形成cDNA,保存于-20 ℃待用。以ACTIN基因為內參基因,采用SYBR PrimeScript TM RT-PCR Kit熒光定量試劑盒在ABI StepOne Plus型熒光定量PCR儀上進行熒光定量檢測。擴增條件: 95 ℃ 30 s; 95 ℃ 5 s,60 ℃ 20 s,循環40次;在72 ℃下單點檢測信號。對每個樣品設3次技術重復,采用2-△△Ct法對各基因進行相對定量分析。
2 結果與分析
2.1 目的基因的擴增與克隆鑒定
以玉木耳的基因組cDNA為模板,擴增得到Actyr基因的全長cDNA序列,經1%瓊脂糖凝膠電泳,所得結果與預期大小相符(圖1)。將PCR擴增獲得的反應產物經電泳、回收、連接、轉化DH5α后,對獲得的單菌落進行菌液PCR及測序,測序結果表明,克隆獲得的Actyr基因cDNA序列長度為1113 bp,編碼371個氨基酸(圖2 );同時,NCBI數據庫的Conserved Domains分析結果(圖3)表明,Actyr屬于Tyrosinase基因家族。
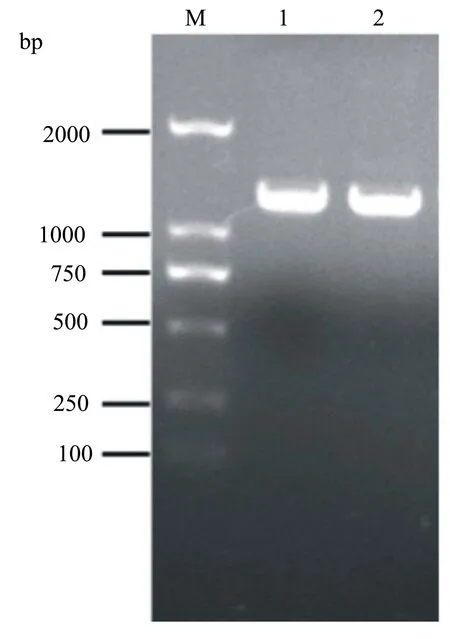
M: DNA分子量標準DNA marker DL2000。1~2: PCR產物。
2.2 Actyr基因的生物信息學分析
應用PredictProtein軟件對Actyr蛋白質進行分析,二級結構分析結果見圖4a,具有α螺旋、β折疊、無規則卷曲;氨基酸組成分析結果見圖4b,其中亮氨酸含量最多,纈氨酸含量最少;亞細胞定位預測結果表明該基因定位于細胞質,見圖4c;通過Protparam軟件預測分析結果顯示,Actyr的等電點為7.64,分子量MW=41.11 kDa。在體外的半衰期為30 h;不穩定指數為48.2,表明該蛋白是不穩定的。通過SWISS-MODEL軟件對Actyr蛋白質的三級結構進行預測,結果如圖4d所示,為下一步對該基因功能的研究奠定了基礎。TMpred軟件的分析結果顯示, Actyr蛋白不存在跨膜區(圖5)。信號肽分析發現不存在信號肽結構(圖6)。

圖2 Actyr基因的cDNA序列及推導的氨基酸序列

圖3 CDS分析結果
把本試驗所克隆的Actyr基因編碼的氨基酸序列與其他物種絡氨酸酶基因編碼的氨基酸序列進行同源性比較分析,發現其在兩個銅離子結合區(CuA和CuB)高度保守(圖7)。
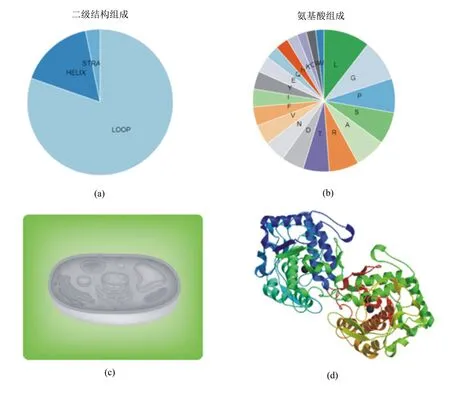
圖4 Actyr基因的生物信息學分析結果
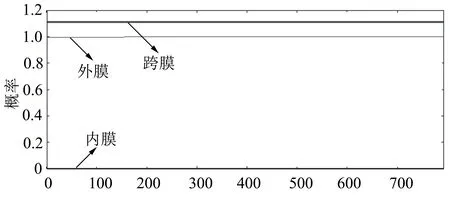
圖5 跨膜結構域分析結果
圖6 信號肽預測結果
2.3 不同發育時期Actyr基因表達的分析
以ACTIN基因為內參基因,對玉木耳不同發育時期Actyr基因的表達量進行分析,發現在玉木耳菌絲生長時期,幾乎檢測不到Actyr基因的表達量;隨著玉木耳的生長發育,其表達量不斷上升,到子實體直徑為3 cm時,Actyr基因的表達量達到最高;隨著玉木耳的進一步發育,其轉錄表達水平逐漸下降(圖8)。
3 討論與結論
本實驗采用基因克隆的手段,獲得了玉木耳的Actyr基因,明確了該基因的基本結構和氨基酸序列。通過生物信息學分析,發現該蛋白屬于絡氨酸酶基因家族成員,具有其典型特征,即在活性中心含有兩個銅離子結合區。此外,通過對其理化性質、結構和功能等方面進行分析預測,結果表明,Actyr基因編碼的蛋白無跨膜區,不存在信號肽;細胞定位預測結果表明其存在于細胞質中,與大部分真菌的絡氨酸酶性質一致[12]。絡氨酸酶廣泛存在于真菌中,在真菌中具有不同的功能,如參與孢子的形成及穩定、防御和毒性機理,參與子實體褐變及色素沉著[1-3,13-15]。此外, Kitamoto等的研究表明,雙核菌絲在黑暗條件下培養時,絡氨酸酶的活性較可見光條件下培養時低[16]。本研究通過分離、克隆玉木耳絡氨酸酶基因,分析其基因表達特征,為進一步明確絡氨酸酶基因在玉木耳中的功能及遺傳育種奠定了基礎。
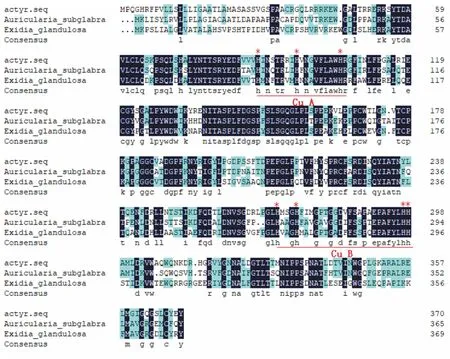
“*”表示保守的組氨酸(His)殘基。
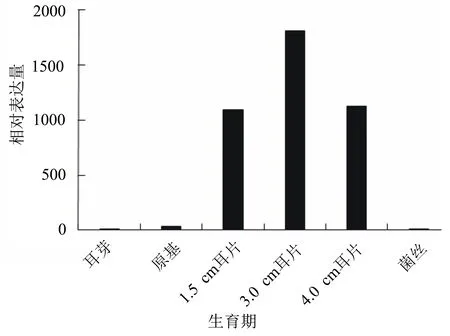
圖8 玉木耳不同生長發育階段Actyr基因的相對表達量
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
