金鐘連翹與東北連翹種間雜交F1代表型性狀遺傳分析
2021-03-22 09:21:08吳雨桐趙崢畑申建雙張啟翔潘會(huì)堂
植物研究 2021年3期
關(guān)鍵詞:模型
徐 洋 吳雨桐 趙崢畑 申建雙 張啟翔 潘會(huì)堂
(花卉種質(zhì)創(chuàng)新與分子育種北京市重點(diǎn)實(shí)驗(yàn)室,國家花卉工程技術(shù)研究中心,城鄉(xiāng)生態(tài)北京實(shí)驗(yàn)室,北京林業(yè)大學(xué)園林學(xué)院,北京 100083)
連翹屬(Forsythia)植物是我國傳統(tǒng)的早春觀花灌木,姿態(tài)優(yōu)美,花色鮮艷,在園林中大量應(yīng)用。連翹屬現(xiàn)有11 個(gè)種,除1 種原產(chǎn)歐洲東南部外,其余均產(chǎn)自亞洲東部,我國產(chǎn)7 種、1 變型,其中1 種系栽培種[1]。19 世紀(jì),連翹屬植物被引到西方國家后,連翹新品種開始大量出現(xiàn)[2]。1885 年,人們發(fā)現(xiàn)了連翹屬的天然雜交種——金鐘連翹(F.intermedia),認(rèn)為它是連翹(F.suspensa)和金鐘花(F.viridissima)的天然雜交后代[3]。1906年,人們利用連翹變種(F.suspensa var. sieboldii)與金鐘花雜交獲得著名的金鐘連翹品種‘Spectabilis’(F.intermedia‘Spectabilis’),該品種在各地得到廣泛栽植。隨后人們發(fā)現(xiàn)了‘Spectabilis’的芽變品種——‘Lynwood’[4]。Flemer 以金鐘連翹品種‘Spectabilis’為母本,卵葉連翹品種‘Tetra Gold’(F.ovata‘Tetra Gold’)為父本進(jìn)行種間雜交,培育出花大色艷、抗寒性強(qiáng)的連翹新品種‘Princeton Gold’[5]。國內(nèi)的連翹育種起步于20 世紀(jì)90 年代,劉瑋等以白丁香和東北連翹為親本進(jìn)行雜交,使用胚培養(yǎng)的方法,獲得了雜交后代植株[6],但未見培育出品種。Shen 等以金鐘連翹品種‘Courtaneur’(F.intermedia‘Courtaneur’)為母本,朝鮮白連翹(Abeliophyllum distichum)為父本,通過遠(yuǎn)緣雜交培育出具有明顯花香的連翹新品種‘春香’[7]。馬帥等以連翹屬9個(gè)種或品種和華北紫丁香作為親本進(jìn)行遠(yuǎn)緣雜交,獲得了雜交后代[8]。目前培育連翹新品種最直接有效的手段仍然是傳統(tǒng)雜交育種方法,主要的育種目標(biāo)性狀集中在抗寒性、生長習(xí)性和花部性狀上。
觀賞植物的表型性狀是基因型和環(huán)境共同作用的結(jié)果,對(duì)生長于同一環(huán)境中的雜交群體的表型性狀進(jìn)行測(cè)定和分析,對(duì)開展雜交育種工作具有重要的指導(dǎo)意義[9]。楊彥伶等以紫薇雜交F1代群體為研究對(duì)象,對(duì)親本和F1代植株的花徑、花序長、花朵數(shù)、株高、冠幅等性狀進(jìn)行了統(tǒng)計(jì)分析,為合理選配紫薇雜交親本和新品種選育提供了依據(jù)[10]。周利君等通過對(duì)月季雜交F1代群體的花部和葉片形態(tài)性狀進(jìn)行了分析,為挖掘控制表型性狀的優(yōu)良基因及輔助選擇育種提供了幫助[11]。張飛等對(duì)菊花雜交F1代6個(gè)花部性狀進(jìn)行遺傳分析,了解了菊花花部性狀的雜種優(yōu)勢(shì)和遺傳基礎(chǔ)[12]。Wang 等通過對(duì)觀賞鳳梨雜交F1代群體的株高、冠幅、分枝數(shù)等性狀進(jìn)行調(diào)查分析,為這些性狀的QTL 定位及輔助選擇育種提供了依據(jù)[13]。表型性狀多樣性是遺傳多樣性和環(huán)境多樣性的綜合表現(xiàn),而雜交育種依賴于對(duì)表型性狀遺傳分離規(guī)律的準(zhǔn)確把握[14]。在連翹方面,Wang 等對(duì)連翹黃色葉性狀的形成和性狀分離情況進(jìn)行了研究,為培育不受或少受環(huán)境影響的黃葉色連翹新品種提供了依據(jù)[15]。目前,有關(guān)連翹雜交后代性狀遺傳規(guī)律的研究仍然較少,很多性狀的遺傳特性尚不清楚,研究連翹主要性狀的遺傳規(guī)律對(duì)連翹新品種定向選育具有重要的指導(dǎo)意義。
本研究對(duì)金鐘連翹品種‘Lynwood’與東北連翹種間雜交獲得的F1代群體的花冠口直徑、花裂片長度、抗寒性等12個(gè)表型性狀進(jìn)行測(cè)定,分析這些性狀的遺傳規(guī)律,并利用植物數(shù)量性狀的主基因+多基因混合遺傳模型[16]對(duì)各性狀進(jìn)行了遺傳分析,以期為連翹的定向育種提供參考。
1 材料與方法
1.1 植物材料
以金鐘連翹品種‘Lynwood’×東北連翹的130株雜交后代和雜交親本作為研究對(duì)象,親本的主要信息見表1 和圖1。該雜交群體露地栽植于國家花卉工程技術(shù)研究中心小湯山基地,該基地位于北京市昌平區(qū)小湯山鎮(zhèn),年平均氣溫11.8℃,年平均日照時(shí)數(shù)2 684 h,年平均降水量550.3 mm。栽植地的溫度、光照、水分、土壤等環(huán)境因子基本相同,植株生長健壯、表現(xiàn)穩(wěn)定。
1.2 雜交
東北連翹開花比金鐘連翹品種‘Lynwood’早約10 d。于2013 年3 月中旬初花期采集東北連翹花粉,置于變色硅膠中干燥,4℃冰箱保存。授粉前采用鐵軍等的方法[17]離體培養(yǎng)花粉檢測(cè)花粉活力,萌發(fā)率在25%以上的花粉用來雜交授粉。在‘Lynwood’開花前一天,人工去雄,套上硫酸紙袋隔離。第二天10:00~12:00 柱頭分泌粘液時(shí)進(jìn)行授粉。授粉后20 d 左右,柱頭開始萎蔫,子房開始膨大時(shí),將硫酸紙袋換成網(wǎng)袋。10 月初果實(shí)成熟后收種子,室內(nèi)陰干存放。11 月下旬,種子經(jīng)100 mg·L-1赤霉素處理6 h 后,點(diǎn)播于裝有草炭基質(zhì)的穴盤中。2014 年3 月上盆,5 月移栽于大田,對(duì)獲得的雜交F1代進(jìn)行大田常規(guī)管理。最終獲得包括130株后代的群體。
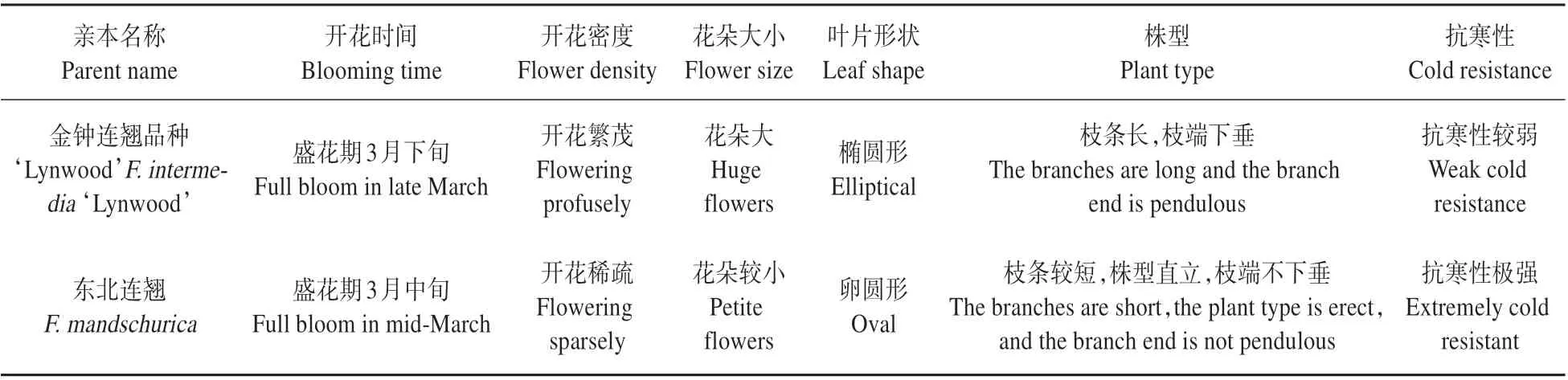
表1 金鐘連翹品種‘Lynwood’和東北連翹的主要性狀Table 1 Main traits of F.intermedia‘Lynwood’and F.mandschurica
1.3 表型指標(biāo)和抗寒指標(biāo)的測(cè)定
表型指標(biāo)測(cè)定:根據(jù)國家林業(yè)局2010 年發(fā)布的《植物新品種特異性、一致性、穩(wěn)定性測(cè)試指南——連翹屬》,于2019 年3~4 月連翹花期,每個(gè)株系選取3朵完全開放的花,用游標(biāo)卡尺測(cè)定花冠口直徑(X1)、花裂片長度(X2)和寬度(X3),并計(jì)算長寬比(X4);選3 個(gè)成熟枝條,測(cè)定10cm 枝條長度內(nèi)花芽的數(shù)量作為著花密度(X5)指標(biāo);8 月,選枝條從頂端向下數(shù)第5~7 節(jié)處的成熟葉片,測(cè)定葉片長度(X6)和寬度(X7),并計(jì)算葉片長寬比(X8);12 月植株落葉后,測(cè)定植株的株高(X9)和冠幅,以植株南北向和東西向的寬度作為冠幅(X10)指標(biāo),選3 個(gè)當(dāng)年生枝條測(cè)定枝條長度(X11)。每個(gè)指標(biāo)重復(fù)3次。
抗寒指標(biāo)測(cè)定:參考Lim&Arora的方法[18],采用電解質(zhì)滲出率擬合Logistic 方程,計(jì)算低溫半致死溫度(LT50)。植株經(jīng)自然抗寒鍛煉后,于2020年1 月上旬,選雜交植株上生長狀況一致、粗細(xì)均勻的1年生枝條,每個(gè)株系剪取8~10個(gè)枝條,剪取的枝條長15~20 cm,用去離子水沖洗干凈,擦干,橫切成0.5 cm 的小段,分7 組用錫箔紙包好,置于5℃冰箱適應(yīng)2 d。取出其中1份材料測(cè)定電導(dǎo)率作為對(duì)照,另外6份采用人工梯度降溫法降溫,溫度梯度為-10、-15、-20、-25、-30、-35℃,降溫速率為5℃/h,降至目標(biāo)溫度后維持12 h,取出1份材料測(cè)定電導(dǎo)率,其他材料繼續(xù)降溫。冷凍的枝條在5℃下放置2 h解凍,然后稱取0.1 g裝入15 mL的離心管中,加入5 mL的去離子水,搖床震蕩24 h,采用DDS-307A電導(dǎo)儀測(cè)定初電導(dǎo)值(C1),將其放入沸水浴中30 min,冷卻至室溫后測(cè)定終電導(dǎo)值(C2)。每種枝條重復(fù)測(cè)定3次,取其平均值[19~20]。計(jì)算公式為:
以各個(gè)株系相對(duì)電導(dǎo)率值y 和相應(yīng)處理溫度x,擬合logistic回歸方程:
式中:k 代表細(xì)胞傷害率的飽和容量,即k=100,依據(jù)方程得到參數(shù)a、b,擬合度顯著時(shí),進(jìn)而求得LT50=-lna/b,作為抗寒性(X12)指標(biāo)[21]。
1.4 數(shù)據(jù)統(tǒng)計(jì)分析
利用Excel2016、SPSS25 進(jìn)行數(shù)據(jù)處理和分析,對(duì)表型性狀進(jìn)行描述性統(tǒng)計(jì)并計(jì)算變異系數(shù),繪制頻率分布圖,進(jìn)行相關(guān)性分析。采用蓋鈞鎰等[16]的植物數(shù)量性狀主基因+多基因混合遺傳模型分析方法,利用SEA 軟件統(tǒng)計(jì)11 種遺傳模型的AIC 值(Akaike’s information criterion),選擇最適合的模型,根據(jù)最小二乘法估算加性效應(yīng)、顯性效應(yīng)、主基因遺傳力等遺傳參數(shù)[22]。
2 結(jié)果與分析
2.1 F1代群體表型性狀的遺傳變異
對(duì)F1代群體植株的花冠口直徑、花裂片長度、抗寒性等12 個(gè)性狀進(jìn)行描述性統(tǒng)計(jì)(見表2)。雜交F1代個(gè)體的不同性狀均有一定程度的差異,存在一定的變異幅度,12 個(gè)性狀的遺傳變異系數(shù)為11.33%~36.89%,變異系數(shù)均超過10%。群體性狀中變異程度最大的是冠幅,變異程度最小的是花裂片長度。著花密度和冠幅的變異系數(shù)超過30%,變異較大。除花裂片長度、花裂片寬度、葉片長度外,其他性狀的變異系數(shù)在15%~30%,為中等變異水平。株高的標(biāo)準(zhǔn)差最大,群體中極易出現(xiàn)極值個(gè)體,葉片長寬比標(biāo)準(zhǔn)差最小。
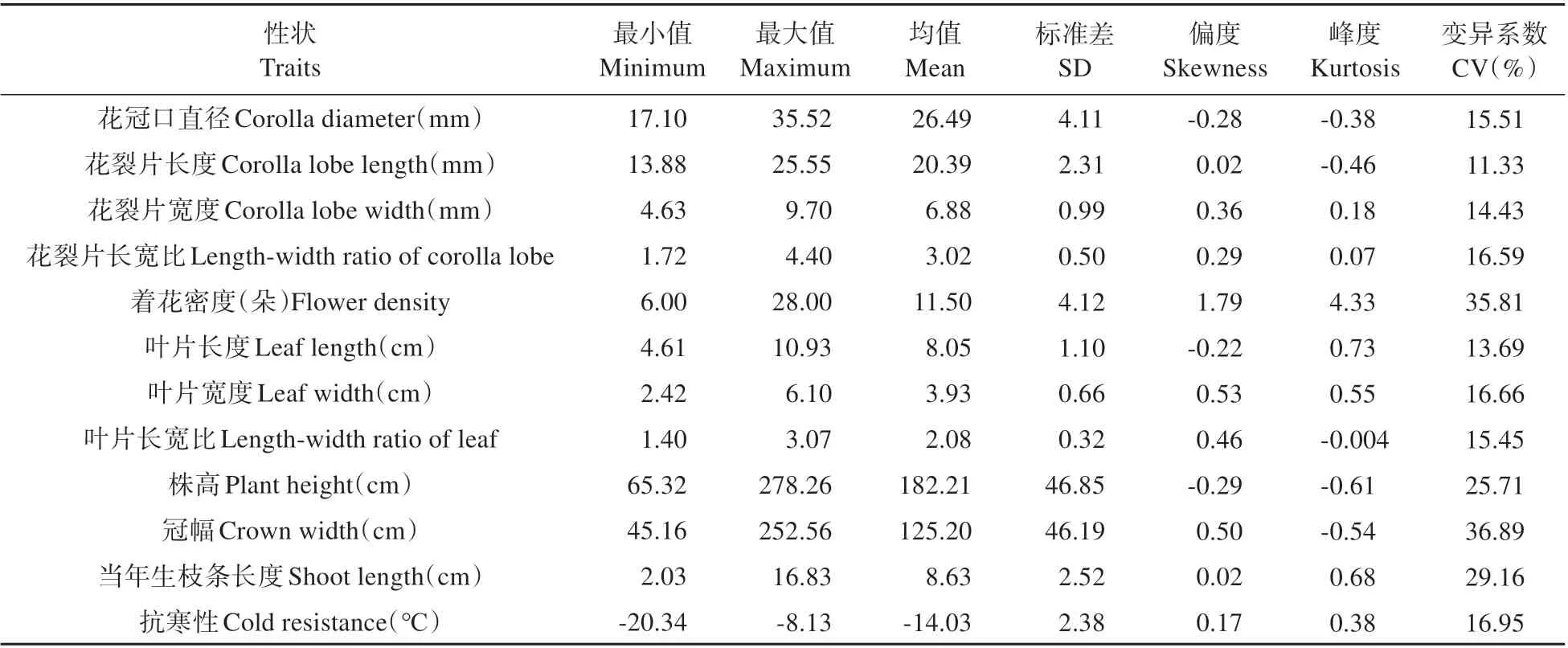
表2 金鐘連翹品種‘Lynwood’×東北連翹群體表型性狀的描述性統(tǒng)計(jì)Table 2 Descriptive statistics of phenotypic traits in F1 population of F.intermedia‘Lynwood’×F.mandschurica
對(duì)F1代群體的表型性狀進(jìn)行頻率分布分析(見圖2),結(jié)果表明,12個(gè)性狀都呈現(xiàn)單峰分布,接近正態(tài)分布,符合數(shù)量性狀的特點(diǎn)。除著花密度外,其他11 個(gè)性狀的偏度和峰度絕對(duì)值均小于1,與正態(tài)曲線擬合較好。著花密度的偏度和峰度大于1,呈現(xiàn)出明顯的正偏性和極端性。
對(duì)F1代群體的12個(gè)性狀與親本進(jìn)行比較分析(見表3),總體上看,雜交后代分離廣泛,各個(gè)性狀均出現(xiàn)大于高親或小于低親的超親個(gè)體。雜交F1代群體中花裂片長寬比的均值大于中親值,花裂片長度、著花密度、葉片長寬比的均值與中親值的比值接近100%,其余性狀的均值小于中親值。除花冠口直徑、花裂片長寬比、葉片長度、冠幅、抗寒性以外,其他性狀介于雙親之間的個(gè)體所占比值最大。F1代群體中多數(shù)個(gè)體(74.8%)的花冠口直徑小于低親,說明花冠口直徑有變小的趨勢(shì)。64.57%的個(gè)體花裂片長寬比大于高親,說明花裂片形狀有偏向更狹長的趨勢(shì)。F1代群體沒有出現(xiàn)冠幅大于高親的個(gè)體,有66.15%的個(gè)體小于低親,證明冠幅有偏向更小的趨勢(shì)。抗寒性大于高親的比例為51.11%,說明抗寒性遺傳偏向于抗寒性較弱的‘Lynwood’,也有相當(dāng)?shù)谋壤?8.89%)介于雙親之間,能夠從中篩選出抗寒性較強(qiáng)的植株,但群體中并未出現(xiàn)抗寒性大于東北連翹的植株。
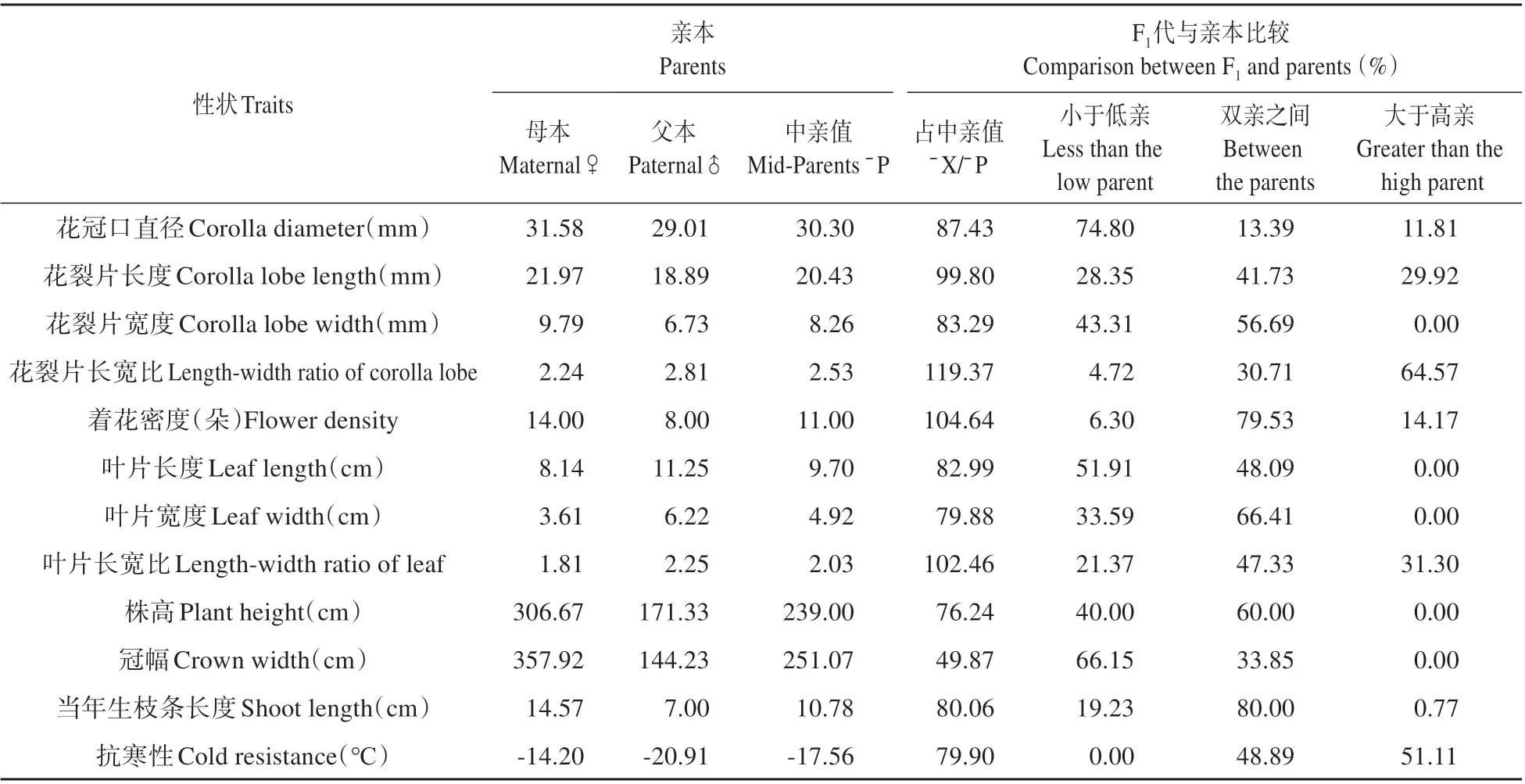
表3 金鐘連翹品種‘Lynwood’×東北連翹群體表型性狀的遺傳分析Table 3 The genetic analysis of phenotypic traits in F1 population of F.intermedia‘Lynwood’×F.mandschurica

表4 金鐘連翹品種‘Lynwood’×東北連翹群體表型性狀的相關(guān)性分析Table 4 Correlation analysis of phenotypic traits in F1 population of F.intermedia‘Lynwood’×F.mandschurica
2.2 F1代群體表型性狀間的相關(guān)性
對(duì)連翹的表型性狀進(jìn)行相關(guān)性分析(見表4)。花冠口直徑與著花密度呈極顯著負(fù)相關(guān)關(guān)系,表明著花密度越大的株系,其花冠口直徑越小。著花密度與株高、冠幅、當(dāng)年生枝條長度均呈現(xiàn)極顯著負(fù)相關(guān)關(guān)系,說明著花密度與株型性狀之間存在著十分緊密的相關(guān)性。當(dāng)年生枝條長度與株高、冠幅呈極顯著正相關(guān)關(guān)系,說明植株的長勢(shì)對(duì)株高和冠幅均有顯著影響。抗寒性與著花密度呈極顯著正相關(guān)關(guān)系,與花冠口直徑、冠幅呈極顯著負(fù)相關(guān)關(guān)系。
2.3 F1代群體表型性狀的遺傳模型
由于連翹自交不親和,基因型高度雜合[23]。其F1代類似于純合親本的F2代,常稱為“假F2代”[24]。因此,可以采用植物數(shù)量性狀混合遺傳模型的主基因+多基因分析方法中的單個(gè)分離世代F2群體的分離分析方法來分析連翹雜交F1代群體表型性狀的主基因遺傳方式,11 個(gè)模型的AIC 值結(jié)果見表5。根據(jù)AIC 準(zhǔn)則挑選AIC 值最小的一個(gè)或相對(duì)較小的幾個(gè)模型,同時(shí),基于均勻性檢驗(yàn)(U12,,U32),Smirnov 檢 驗(yàn)(nW2)及Kolmogorov 檢 驗(yàn)(Dn)對(duì)候選的模型進(jìn)行適合性檢驗(yàn)。比較候選模型的統(tǒng)計(jì)量顯著性情況,數(shù)目最少的確定為最優(yōu)遺傳模型。如果候選模型之間顯著性數(shù)目一樣,則AIC 值小的被認(rèn)定為最優(yōu)遺傳模型[16]。以著花密度為例,AIC 值較小的模型有1MG-AD,2MGADI,2MG-AD 作為備選最適模型。然后利用適合性檢驗(yàn)(見表5~6),結(jié)果顯示3 個(gè)模型顯著數(shù)目都為2,而2MG-AD 模型的AIC 值最小,所以著花密度的最適遺傳模型為2MG-AD。
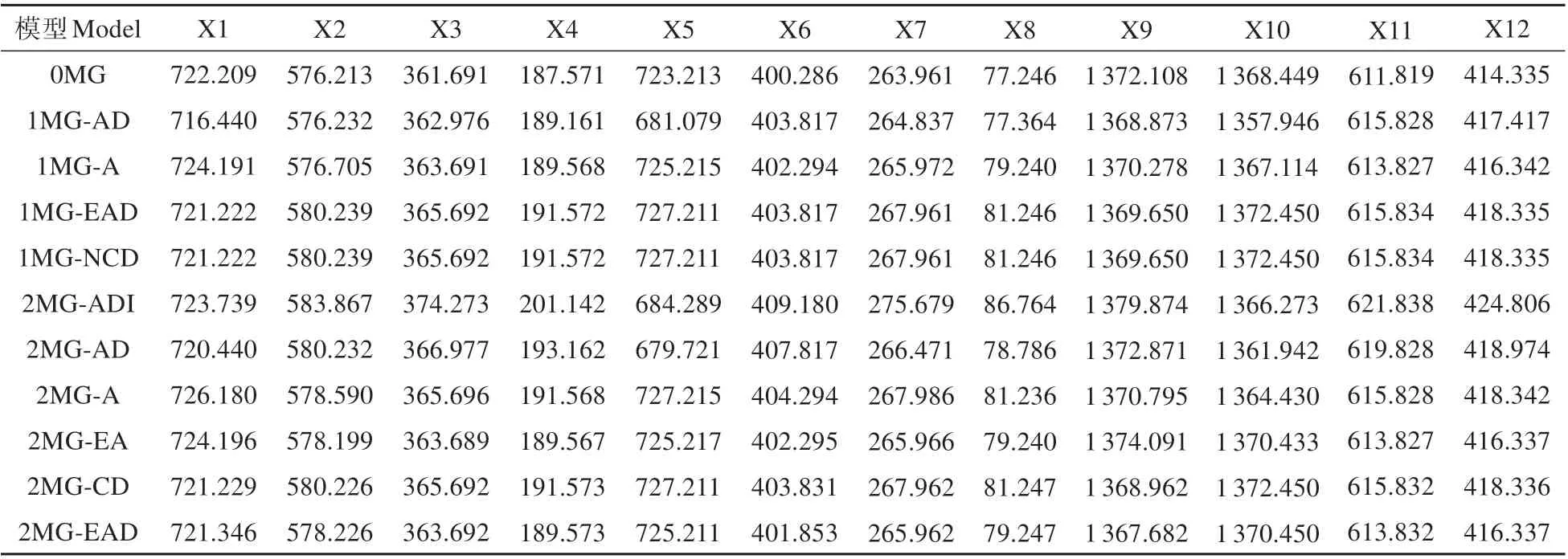
表5 金鐘連翹品種‘Lynwood’×東北連翹群體表型性狀各種遺傳模型的AIC值Table 5 AIC values of various genetic models of phenotypic traits in F1 population of F.intermedia‘Lynwood’×F.mandschurica
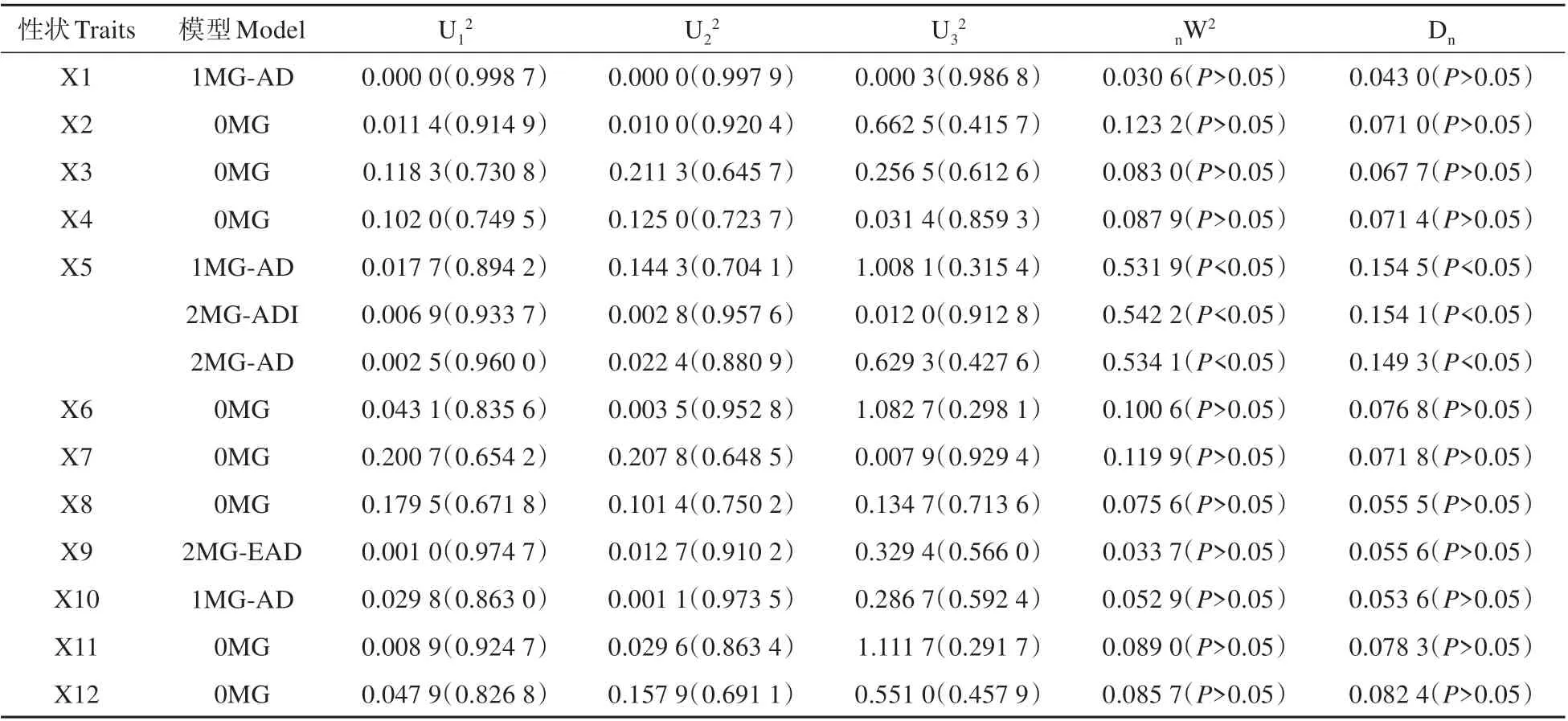
表6 入選模型的適合性檢驗(yàn)Table 6 Test for goodness-of-fit of selected genetic models
對(duì)入選模型進(jìn)行適合性檢驗(yàn)(見表6)。花裂片長度、花裂片寬度、花裂片長寬比、葉片長度、葉片寬度、葉片長寬比、當(dāng)年生枝條長度、抗寒性的最適模型為0MG 模型,這些性狀的遺傳由受環(huán)境影響比較大的多基因控制。花冠口直徑和冠幅的最適模型為1MG-AD 模型,這些性狀由一對(duì)加性—顯性主基因控制。著花密度的最適模型為2MG-AD模型,由兩對(duì)加性—顯性主基因控制。株高的最適模型為2MG-EAD 模型,由兩對(duì)等加性—顯性主基因控制[25]。
2.4 F1代群體表型性狀的遺傳參數(shù)
根據(jù)已選出的最適模型,采用最小二乘法估計(jì)除OMG 模型性狀以外的其余性狀的一階、二階遺傳參數(shù)(見表7)。控制花冠口直徑和冠幅的一對(duì)主基因的加性效應(yīng)值分別為5.27 和40.48,顯性效應(yīng)值分別為0.77和-37.97,加性效應(yīng)均大于顯性效應(yīng),主基因遺傳力分別為76.05%和60.3%。控制著花密度的兩對(duì)主基因的加性效應(yīng)值分別為4.99 和2.58,顯性效應(yīng)值分別為-7.02 和-0.91,說明兩對(duì)主基因的加性效應(yīng)均為正向增效,而顯性效應(yīng)均為負(fù)向效應(yīng)。控制株高的主基因的等加性—顯性效應(yīng)值為30.71,主基因遺傳力為64.75%。
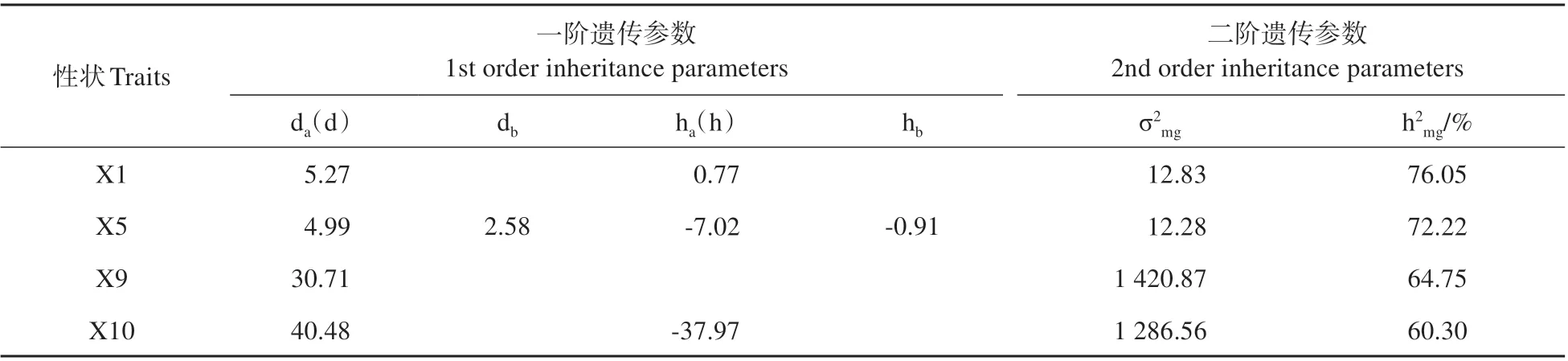
表7 不同表型性狀在各最適模型下的遺傳參數(shù)估計(jì)值Table 7 Estimation of genetic parameters of phenotypic traits at its optimal genetic model
3 討論
表型性狀的多樣性是育種工作的基礎(chǔ),了解和掌握雜交群體的表型性狀的多樣性水平和變異程度,對(duì)于新品種培育具有重要意義[26]。焦垚等通過對(duì)紫薇雜交F1代性狀的統(tǒng)計(jì)分析,發(fā)現(xiàn)雜交后代在株高、冠幅上更容易產(chǎn)生變異[27]。Dhananjaya 等通過測(cè)定分析香石竹與中國石竹種間雜交F1代表型性狀的遺傳變異,發(fā)現(xiàn)了切花效果優(yōu)良、產(chǎn)量高的新品種,并且掌握了遺傳規(guī)律用以指導(dǎo)后續(xù)育種工作[28]。本研究所測(cè)定的金鐘連翹與東北連翹雜交F1代群體的12 個(gè)表型性狀均分離廣泛,變異系數(shù)為11.33%~36.89%。根據(jù)李娟娟等對(duì)遺傳變異度判定的標(biāo)準(zhǔn)[29],本研究中花裂片長度、花裂片寬度、葉片長度遺傳變異度較小,遺傳改良潛力一般。而花冠口直徑、花裂片長寬比、著花密度、葉片寬度、葉片長寬比、株高、冠幅、當(dāng)年生枝條長度、抗寒性等性狀遺傳變異度均達(dá)到中等水平以上,具有較大的遺傳改良潛力,性狀分離廣泛,能夠在后代群體中篩選出觀賞性和抗寒性優(yōu)良的連翹新品種。
通過對(duì)表型性狀之間的相關(guān)性研究,可在育種中合理利用不同性狀間的關(guān)系,來預(yù)測(cè)相關(guān)的性狀表現(xiàn),有效提高育種效率,加快育種進(jìn)程。Kumar 等通過分析發(fā)現(xiàn),唐菖蒲每個(gè)穗上花的數(shù)量與花朵直徑呈顯著正相關(guān),開花的時(shí)長與穗長呈顯著負(fù)相關(guān)關(guān)系[30]。何貴平等通過試驗(yàn)發(fā)現(xiàn),杉木的結(jié)頂率與凍害率存在顯著負(fù)相關(guān)關(guān)系,可以通過了解結(jié)頂率指標(biāo)間接進(jìn)行抗寒性材料的初選[31]。在本研究中植株抗寒性與花冠口直徑、冠幅呈極顯著的負(fù)相關(guān),與著花密度呈極顯著正相關(guān)關(guān)系。因此,在抗寒性育種過程中,可以選取比較容易測(cè)定的性狀進(jìn)行參考,以提高抗寒性狀的育種效率。
植物大多數(shù)表型性狀都是由微效多基因控制的數(shù)量性狀,在同一雜交后代群體的不同個(gè)體中往往表現(xiàn)出連續(xù)的數(shù)量差異,易受環(huán)境影響,不易明確分組歸類,需要應(yīng)用統(tǒng)計(jì)學(xué)方法對(duì)整個(gè)群體進(jìn)行測(cè)定和分析[32]。張琳等利用主基因+多基因混合遺傳模型對(duì)牡丹雜交F1代20個(gè)表型性狀進(jìn)行分析,結(jié)果表明株高、冠幅等8 個(gè)性狀由微效多基因控制,新枝長度、花朵數(shù)等12個(gè)性狀受到一對(duì)或兩對(duì)主基因控制[33]。Zhang 等對(duì)菊花雜交群體的初始開花時(shí)間和開花持續(xù)時(shí)間進(jìn)行了混合遺傳模型分析,結(jié)果發(fā)現(xiàn)兩個(gè)性狀的主基因遺傳力分別為65%和72%,遺傳力較大,在早期育種就可進(jìn)行選擇[34]。本研究發(fā)現(xiàn)花裂片長度、花裂片寬度、花裂片長寬比、葉片長度、葉片寬度、葉片長寬比、當(dāng)年生枝條長度、抗寒性等8個(gè)性狀的遺傳無主基因控制,受環(huán)境影響較大的微效多基因控制;花冠口直徑和冠幅由一對(duì)加性—顯性主基因控制;著花密度由兩對(duì)加性—顯性主基因控制;株高由兩對(duì)等加性—顯性主基因控制。花冠口直徑、冠幅、著花密度和株高的主基因遺傳力分別為76.05%、60.3%、72.22%和64.75%,均大于50%,屬于高度遺傳力,受環(huán)境影響較小,在早期世代即可進(jìn)行選擇[35]。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網(wǎng)絡(luò)安全與數(shù)據(jù)管理(2022年1期)2022-08-29 03:15:20
導(dǎo)航定位學(xué)報(bào)(2022年4期)2022-08-15 08:27:00
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:36
成都醫(yī)學(xué)院學(xué)報(bào)(2021年2期)2021-07-19 08:35:14
新世紀(jì)智能(數(shù)學(xué)備考)(2020年9期)2021-01-04 00:25:14
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
