鎘對(duì)體外培養(yǎng)雞骨髓基質(zhì)細(xì)胞成骨分化的毒性作用
2021-03-12 06:11:30趙玉田王果帥張雪晴李賽慧閔雯嫣劉宗平顧建紅
畜牧與獸醫(yī) 2021年3期
關(guān)鍵詞:檢測(cè)
趙玉田,王果帥,張雪晴,李賽慧,閔雯嫣,劉宗平,顧建紅*
(1. 揚(yáng)州大學(xué)獸醫(yī)學(xué)院,江蘇 揚(yáng)州 225009;2. 江蘇高校動(dòng)物重要疫病與人獸共患病防控協(xié)同創(chuàng)新中心,江蘇 揚(yáng)州 225009)
鎘是一種不易分解的重金屬環(huán)境污染物,其生物半衰期為10~30年,進(jìn)入環(huán)境后不能被微生物降解,并可通過食物鏈蓄積在體內(nèi)[1-2]。急性或慢性鎘暴露能引起肝腎及骨骼等多種器官和組織的損傷。因此,鎘的研究愈來愈廣泛。骨作為鎘毒性作用的主要靶器官之一,接觸高濃度鎘會(huì)降低骨密度(bone mineral density,BMD),通常表現(xiàn)為骨質(zhì)疏松、關(guān)節(jié)炎等癥狀[3]。近年來,由于工業(yè)的快速發(fā)展,大氣和飼料中的鎘污染日益嚴(yán)重,對(duì)動(dòng)物健康造成了極大的威脅[4]。
骨髓基質(zhì)細(xì)胞(bone marrow stromal cells,BMSCs)是一種骨髓多能細(xì)胞,在體內(nèi)處于相對(duì)靜止?fàn)顟B(tài),當(dāng)受到各種生理或病理刺激時(shí)能分化為成骨細(xì)胞(osteoblast,OB)、軟骨細(xì)胞、脂肪細(xì)胞、心肌細(xì)胞和骨骼肌細(xì)胞等[5-6]。BMSCs純化需時(shí)短,并可在體外迅速擴(kuò)增,作為骨髓微環(huán)境的重要組成成分,對(duì)于維持正常的骨代謝具有極其重要的作用[7-8]。鎘引起骨質(zhì)疏松的確切機(jī)制尚不清楚[9]。有文獻(xiàn)報(bào)道,氯化鎘可促進(jìn)人BMSCs的成脂分化、抑制其成骨分化[10]。大鼠試驗(yàn)結(jié)果表明,低劑量的鎘可以通過RANKL/OPG信號(hào)下調(diào)BMSCs的成骨分化基因[11],也可通過Wnt/β-catenin通路抑制BMSCs的成骨分化[12]。然而,人和大鼠屬于哺乳類動(dòng)物,其骨骼與家禽存在差異。禽類骨骼中空,壁薄而輕,不僅在支撐機(jī)體中發(fā)揮重要作用,在蛋殼形成中亦發(fā)揮重要作用。目前,鎘對(duì)禽類BMSCs成骨分化的研究未見報(bào)道。本研究將在誘導(dǎo)雞胚來源BMSCs成骨分化基礎(chǔ)上,加入不同濃度的醋酸鎘,觀察其對(duì)雞體外培養(yǎng)BMSCs成骨分化的影響,為禽類骨代謝失衡疾病的研究奠定基礎(chǔ)。
1 材料與方法
1.1 實(shí)驗(yàn)動(dòng)物
SPF雞蛋由山東濟(jì)南鑫盛達(dá)科技有限公司購入,在揚(yáng)州大學(xué)獸醫(yī)學(xué)院內(nèi)科實(shí)驗(yàn)室孵化至18日齡。
1.2 雞胚BMSCs的分離與培養(yǎng)
無菌條件下,分離雞胚股骨、脛骨,用不含血清的α-MEM培養(yǎng)基沖出骨髓,沖至發(fā)白,300目濾網(wǎng)過濾,離心留沉淀于管底的細(xì)胞,用α-MEM培養(yǎng)基(含10%FBS)重懸,以3×106個(gè)/mL密度接種于細(xì)胞瓶,37 ℃、5%CO2,飽和濕度的培養(yǎng)箱中培養(yǎng)。
BMSCs在24 h完全貼壁,換液去除未貼壁細(xì)胞,待細(xì)胞長(zhǎng)至80%~90%時(shí),進(jìn)行傳代培養(yǎng)。α-MEM培養(yǎng)基先于37 ℃水浴加熱15 min,取出細(xì)胞瓶,在超凈臺(tái)內(nèi)緩慢棄去培養(yǎng)基,用磷酸鹽緩沖液(phosphate buffered solution,PBS)清洗2次,再用預(yù)熱的0.25%胰酶37 ℃消化,在顯微鏡下觀察至細(xì)胞間連接消失時(shí),加入含有10%胎牛血清的α-MEM培養(yǎng)基終止消化,用膠頭滴管緩慢吹下貼壁細(xì)胞,PBS清洗后用含10%胎牛血清的α-MEM培養(yǎng)基重懸細(xì)胞,計(jì)數(shù),以3×106個(gè)/mL接種于細(xì)胞瓶,置于37 ℃,5%CO2,飽和濕度的培養(yǎng)箱中培養(yǎng)。
1.3 雞BMSCs的形態(tài)學(xué)觀察及成骨分化能力檢測(cè)
取第二代雞BMSCs,以每孔3×105個(gè)接種于六孔板,待長(zhǎng)至75%左右改為成骨誘導(dǎo)液(10%FBS、地塞米松10 nmol/L、β-甘油磷酸鈉10 mmol/L、抗壞血酸50 μg/mL的DMEM),誘導(dǎo)7 d時(shí),進(jìn)行堿性磷酸酶(alkaline phosphatase,ALP)染色,誘導(dǎo)21 d時(shí)進(jìn)行礦化結(jié)節(jié)茜素紅染色并拍照記錄。
1.4 流式細(xì)胞儀檢測(cè)BMSCs特異性表面標(biāo)志物
第二代細(xì)胞長(zhǎng)至約80%時(shí),棄去培養(yǎng)基,PBS洗2次,加入2 mL胰蛋白酶消化30 s,含10% FBS的α-MEM培養(yǎng)基終止消化,收集細(xì)胞,1 000 r/min,離心10 min,PBS洗2次,用100 μL PBS重懸。對(duì)照組不加任何抗體,實(shí)驗(yàn)組分別加入2 μL 抗體(CD44-FITC、CD45-PE、CD44-FITC和CD45-PE),冰上孵育30 min后加入1 mL PBS清洗2次,用300 μL PBS重懸細(xì)胞轉(zhuǎn)移至流式管,F(xiàn)ACS LSRFortessa流式細(xì)胞分析儀檢測(cè)。
1.5 CCK-8法檢測(cè)增殖率
取第二代細(xì)胞,按照上述方法傳至96孔板(接種密度7 500個(gè)/孔),待其長(zhǎng)至75%左右,改為成骨誘導(dǎo)培養(yǎng)基(10%FBS、地塞米松10 nmol/L、β-甘油磷酸鈉10 mmol/L、抗壞血酸50 μg/mL的DMEM),并加入0,1,2,5,10和20 μmol/L的醋酸鎘處理72 h,按照CCK-8試劑盒說明書,在450 nm處采用酶標(biāo)儀測(cè)定吸光度值。
1.6 ALP染色測(cè)定ALP活性
取第二代細(xì)胞以每孔3×105個(gè)細(xì)胞接種至6孔板,待長(zhǎng)至75%左右改為成骨誘導(dǎo)培養(yǎng)基(10%FBS、地塞米松10 nmol/L、β-甘油磷酸鈉10 mmol/L、抗壞血酸50 μg/mL的DMEM),并加入0和5 μmol/L的醋酸鎘,處理72 h,按BCIP?/NBT液體底物系統(tǒng)試劑盒操作。
1.7 實(shí)時(shí)熒光定量PCR
取第二代細(xì)胞以每孔3×105個(gè)細(xì)胞接種至6孔板,待長(zhǎng)至75%左右改為成骨誘導(dǎo)培養(yǎng)基(10%FBS、地塞米松10 nmol/L、β-甘油磷酸鈉10 mmol/L、抗壞血酸50 μg/mL的DMEM),并加入0、5、10和20 μmol/L的醋酸鎘,處理72 h,TRizol裂解法提取各組總RNA。使用qPCR儀檢測(cè)成骨相關(guān)標(biāo)志分子:Runt相關(guān)轉(zhuǎn)錄因子2(Runx2)、ALP、成骨細(xì)胞特異性轉(zhuǎn)錄因子(osterix,OSX)、骨鈣素(osteocalcin,OCN)、骨橋蛋白(osteopontin,OPN)、Ⅰ型膠原(type Ⅰ collagen,COL1);破骨細(xì)胞生成和抑制因子:核因子κB受體活化因子配體(RANKL)和骨保護(hù)素(osteoprotegerin,OPG)mRNA表達(dá)水平。以GAPDH為參照,按照RT-PCR Kit進(jìn)行qRT-PCR反應(yīng)。引物序列見如表1。

表1 成骨相關(guān)基因引物序列
1.8 數(shù)據(jù)分析
所有數(shù)值均使用GraphPad Prism軟件分析作圖,數(shù)據(jù)用“平均值±標(biāo)準(zhǔn)差”表示。P<0.05表示組間差異顯著,P<0.01表示組間差異極顯著。
2 結(jié)果
2.1 BMSCs形態(tài)觀察及成骨分化能力檢測(cè)
剛接種的原代BMSCs呈圓形,24 h后開始貼壁,貼壁細(xì)胞呈梭形,體積增大但混有大量紅細(xì)胞、白細(xì)胞等不貼壁橢圓形或圓形細(xì)胞。傳至第一代,細(xì)胞形態(tài)趨于均一,紡錘形細(xì)胞增多,圓形細(xì)胞減少。傳至第二代細(xì)胞形態(tài)更加均一,紡錘形細(xì)胞占80%以上(圖1)。成骨誘導(dǎo)培養(yǎng)基培養(yǎng)7 d ALP染色結(jié)果顯示,細(xì)胞呈紫黑色(圖2)。成骨誘導(dǎo)至21 d時(shí),茜素紅染色可見橘紅色的礦化結(jié)節(jié)形成(圖3)。

圖1 雞BMSCs形態(tài)(100×)

圖2 雞BMSCs ALP 染色(100×)
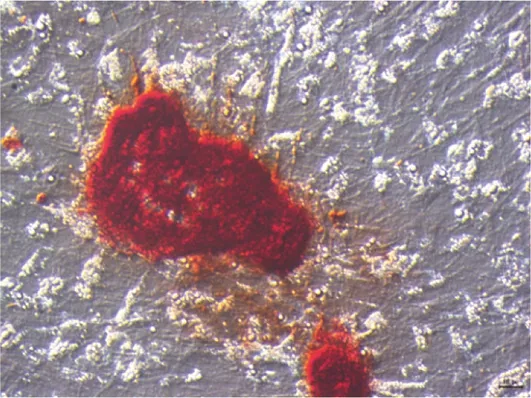
圖3 礦化結(jié)節(jié)茜素紅染色(100×)
2.2 流式檢測(cè)特異性標(biāo)志物
流式細(xì)胞術(shù)檢測(cè)第二代BMSCs,結(jié)果顯示,CD44(64.9%)為陽性(圖4),CD45(0.5%)為陰性(圖5),符合基質(zhì)細(xì)胞表型特征。
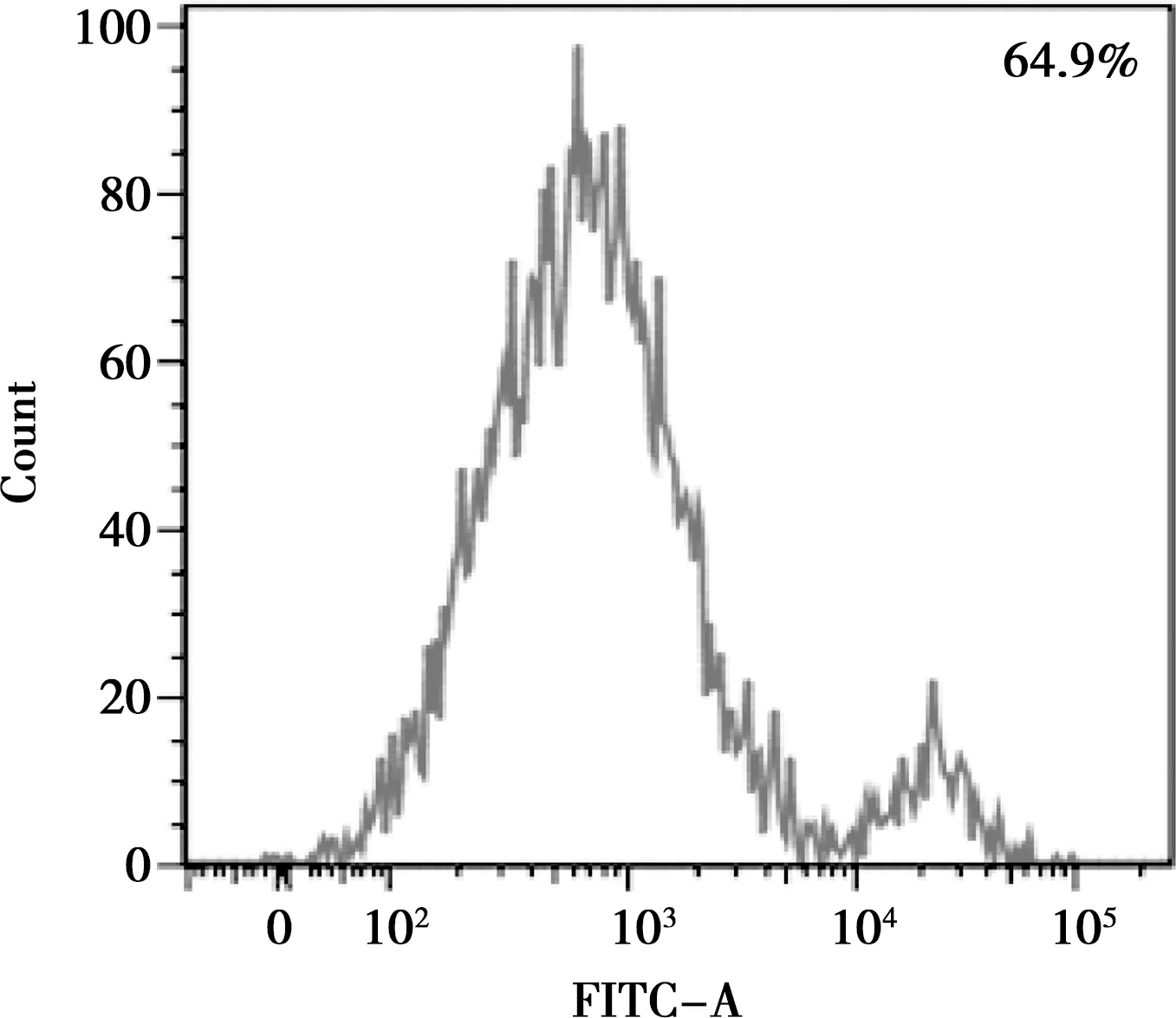
圖4 CD44-FITC
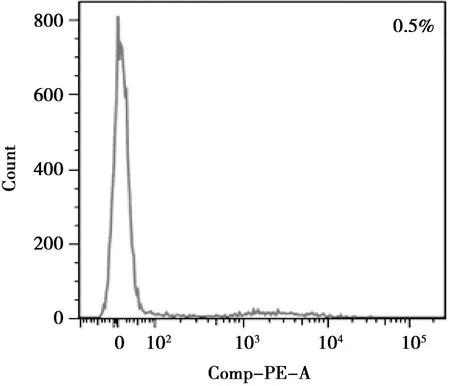
圖5 CD45-PE
2.3 鎘對(duì)BMSCs增殖率的影響
CCK-8法檢測(cè)不同濃度醋酸鎘處理72 h的細(xì)胞增殖率,以確定醋酸鎘的細(xì)胞毒性。結(jié)果顯示,低濃度的鎘對(duì)細(xì)胞的存活率有顯著的促進(jìn)作用(P<0.05),當(dāng)醋酸鎘濃度達(dá)到20 μmol/L時(shí),可顯著抑制細(xì)胞活力(P<0.05)(圖6)。
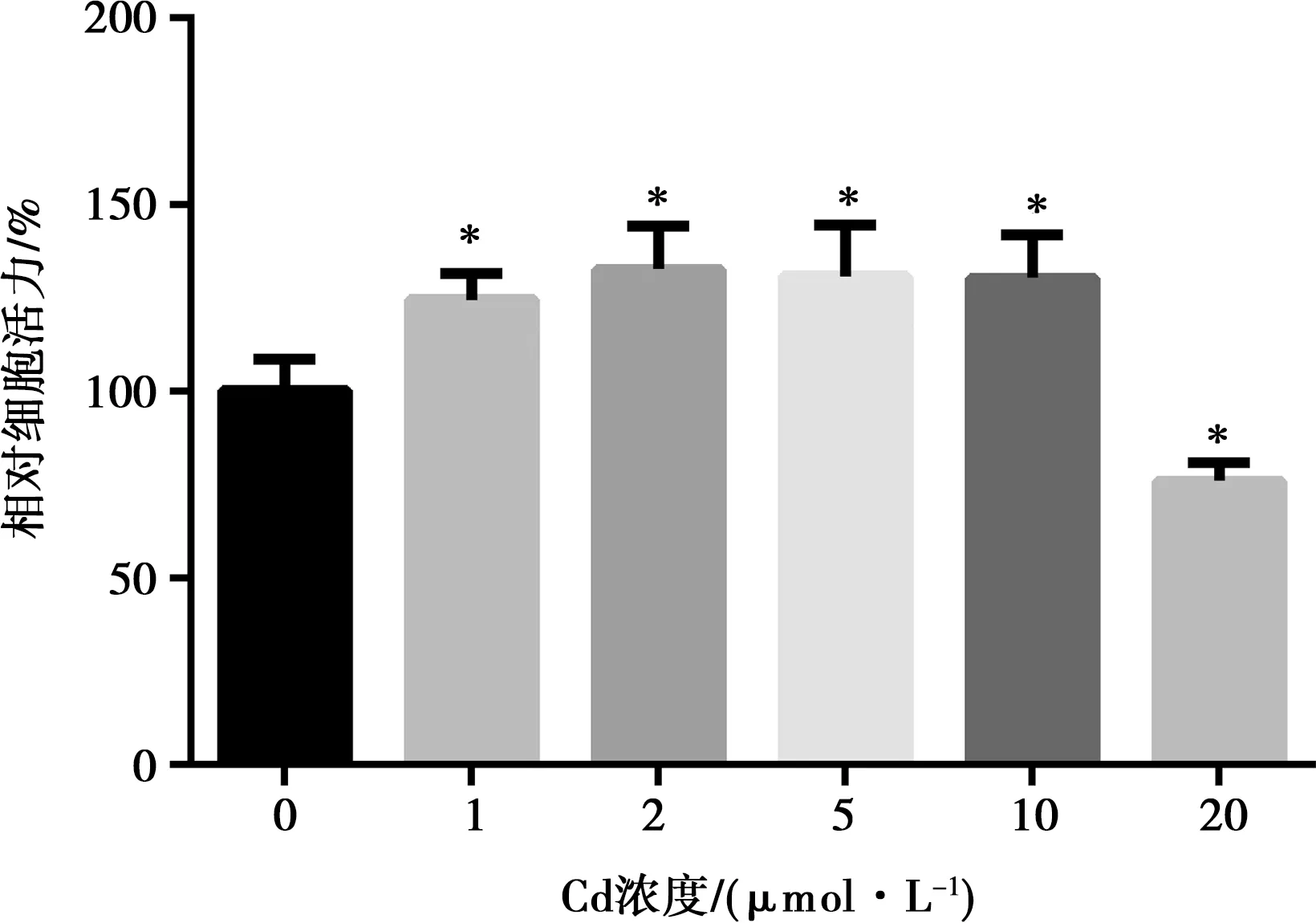
注:*表示與空白對(duì)照組比較,差異顯著(P<0.05)
2.4 鎘對(duì)BMSCs成骨分化過程中ALP活性的影響
ALP染色法檢測(cè)細(xì)胞堿性磷酸酶的活性,結(jié)果顯示,空白對(duì)照組細(xì)胞存在大量紫黑色的染色顆粒(圖7A),醋酸鎘處理組紫黑色面積較少(圖7B),說明鎘處理組ALP活性明顯降低。
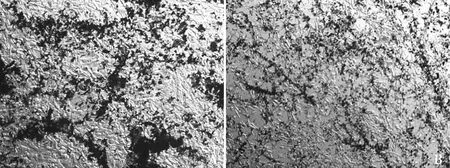
A. 空白組;B. 鎘處理組
2.5 RT-PCR檢測(cè)OB相關(guān)基因
熒光定量PCR結(jié)果顯示,OPN、COL1、Runx2、OSX、ALP、OCN和OPG mRNA的表達(dá)量隨著鎘濃度的升高劑量依賴性降低,除了5 μmol/L鎘組OPN差異不顯著外,其余各組差異均極顯著(P<0.01)(圖8A-H)。RANKL mRNA的表達(dá)量呈濃度依賴性升高,差異均極顯著(P<0.01)(圖8A~H)。

A. OPN mRNA的相對(duì)表達(dá);B. COL1 mRNA的相對(duì)表達(dá);C. Runx2 mRNA的相對(duì)表達(dá);D. OSX mRNA的相對(duì)表達(dá);E. ALP mRNA的相對(duì)表達(dá);F. OCN mRNA的相對(duì)表達(dá);G. OPG mRNA的相對(duì)表達(dá);H. RANKL mRNA的相對(duì)表達(dá)。**表示與對(duì)照組比較差異極顯著(P<0.01)
3 討論
目前報(bào)道BMSCs分離的方法不止一種,包括差速貼壁分離法、密度梯度離心和流式細(xì)胞術(shù)分離法等[13]。王翔毅等[14]發(fā)現(xiàn)體外密度梯度離心法和貼壁靜置可以獲得大量高度均一、表型符合BMSCs的細(xì)胞。根據(jù)BMSCs與其他細(xì)胞貼壁能力、沉降速度的差異,首次換液時(shí)間格外重要,接種后24 h通過首次全量換液以防止雜細(xì)胞貼壁,傳代時(shí)控制胰酶消化時(shí)間為30 s[15]。目前還沒發(fā)現(xiàn)BMSCs的特異表面標(biāo)志,主要根據(jù)其可表達(dá)CD44、CD54、CD106等間質(zhì)內(nèi)皮細(xì)胞標(biāo)志,但不表達(dá)CD34、CD45等造血細(xì)胞標(biāo)志進(jìn)行鑒定[16-18]。本試驗(yàn)采用全骨髓差速貼壁法,接種后24 h全量換液,待其長(zhǎng)至80%時(shí),傳代培養(yǎng)。原代細(xì)胞可見大量圓形或橢圓形雜細(xì)胞,傳至第一代時(shí)圓形細(xì)胞減少,細(xì)胞呈典型的紡錘形,體積較大,邊緣光亮,可穩(wěn)定傳代培養(yǎng)。成骨誘導(dǎo)培養(yǎng)基培養(yǎng)第二代BMSCs 7 d細(xì)胞ALP染色陽性,培養(yǎng)21 d可形成礦化結(jié)節(jié)。流式細(xì)胞術(shù)檢測(cè)顯示第二代細(xì)胞CD44陽性,CD45陰性,符合BMSCs特征。表明本試驗(yàn)獲得的細(xì)胞為雞BMSCs,可用于下一步試驗(yàn)。
鎘作為重要的環(huán)境重金屬污染物,可增加人和動(dòng)物骨質(zhì)疏松的風(fēng)險(xiǎn)[19]。高劑量的鎘可明顯損害大鼠脛骨骨微結(jié)構(gòu)[20]。早期研究表明,鎘的骨毒性繼發(fā)于腎損傷,尤其腎小管損傷可致腎小管鈣、磷重吸收障礙;同時(shí)維生素D活化受損,繼發(fā)鈣、磷代謝異常,引發(fā)骨丟失和骨質(zhì)疏松癥狀[21]。最近研究表明,鎘可直接作用于OB,引起骨損傷[22]。然而,鎘是否影響禽BMSCs成骨分化,從而導(dǎo)致骨骼疾病仍不清楚。OB發(fā)育包括活化增殖期、細(xì)胞外基質(zhì)成熟期和基質(zhì)礦化期3個(gè)階段。ALP是OB成熟及活性的標(biāo)志[23]。Runx2是促進(jìn)骨形成的關(guān)鍵調(diào)控因子,為成骨分化早期標(biāo)志物。OSX是Runx2在成骨過程中的下游靶點(diǎn),激活COL1A1和OCN的轉(zhuǎn)錄,實(shí)現(xiàn)骨形成[24]。OCN由成熟的礦化OB合成,是OB分化中晚期標(biāo)志物,其可通過與鈣及羥基磷灰石結(jié)合促進(jìn)骨形成[25]。OPN主要表達(dá)于骨形成晚期,調(diào)控礦化組織的重建[26];此外,表達(dá)于OB或BMSCs的RANKL/OPG信號(hào)是近20年發(fā)現(xiàn)的調(diào)控破骨細(xì)胞生成和骨溶解的關(guān)鍵信號(hào)軸[27]。因此ALP、RUNX2、OSX、COL1A1、OCN、OPN的表達(dá)水平反映著BMSCs的成骨分化能力,RANKL和OPG表示骨吸收能力。本研究在體外誘導(dǎo)雞BMSCs成骨分化過程中用不同濃度醋酸鎘處理,觀察了鎘對(duì)BMSCs增殖及上述成骨標(biāo)志物表達(dá)的影響。結(jié)果顯示,低濃度(≤10 μmol/L)鎘顯著促進(jìn)BMSCs增殖,高濃度(大于10 μmol/L)鎘顯著抑制BMSCs增殖;然而,5 μmol/L鎘暴露均可顯著或極顯著下調(diào)成骨相關(guān)指標(biāo)(COL1、OSX、RUNX2、ALP、OCN、OPN)以及OPG的表達(dá),上調(diào)RANKL基因的表達(dá),且呈劑量依賴效應(yīng)。研究表明,鎘能夠抑制體外培養(yǎng)雞BMSCs向OB的分化,低濃度鎘對(duì)BMSCs增殖的促進(jìn)作用主要是反饋調(diào)節(jié)引起。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
