擴展青霉侵染對蘋果果實膜磷脂代謝的影響
2021-03-02 07:06:06魏亞楠宗元元DovPRUSKYEdwardSIONOV
食品科學 2021年3期
彭 慧,龔 迪,魏亞楠,楊 乾,宗元元,Dov PRUSKY,,Edward SIONOV,畢 陽,*
(1.甘肅農業大學食品科學與工程學院,甘肅 蘭州 730070;2.以色列農業研究組織農產品采后科學部,以色列 里雄萊錫安 7505101)
由擴展青霉(Penicillium expansum)引起的青霉病是蘋果的重要采后病害,青霉病不僅導致蘋果腐爛及品質劣變,而且會在果實體內積累棒曲霉毒素[1]。P.expansum主要經由果實表面的傷口侵染,通過分泌胞外酶致使傷口處組織潰爛[2]。但該病原菌侵染如何影響果實細胞膜完整性和與其相關的磷脂代謝尚不清楚。前人研究表明,多種真菌侵染均會增加果實的細胞膜透性[3],P.expansum侵染會導致蘋果果實細胞膜透率升高[4];輪紋病菌(Botryosphaeria berengriana)和褐腐病菌(Monilinia fructigena)侵染也會增加梨果實的膜透性[5];在膠孢炭疽菌(Colletotrichum gloeosporioides)侵染的柑橘果實中也可觀察到類似的現象[6]。進一步的研究表明,病原菌侵染所導致的植物細胞膜透性增加與磷脂酶活性的提高、膜磷脂的降解[7]、磷脂酸(phosphatidic acid,PA)和脂肪酸的釋放[8]有關。焦腐病菌(Lasiodiplodia theobromae)侵染會增加龍眼果實的磷脂酶D(phospholipase D,PLD)活性、降低磷脂酰膽堿(phosphatidylcholine,PC)和磷脂酰肌醇(phosphatidylinositol,PI)的含量、提高PA和飽和脂肪酸含量[9];同樣,霜疫霉菌(Peronophythora litchii)侵染也顯著提高了荔枝果實的PLD活性[10],增加了棕櫚酸、硬脂酸、油酸和亞油酸含量[11];此外,擬莖點霉(Phomopsis longanae)侵染龍眼后果實飽和脂肪酸含量增加、不飽和脂肪酸含量和脂肪酸不飽和度降低[12]。由此表明,真菌侵染能夠激活果實的膜磷脂代謝,導致果實PA和脂肪酸的釋放[13-14]。盡管已有真菌侵染對果實細胞膜完整性及磷脂代謝影響的報道,但在不同侵染階段果實細胞膜完整性以及膜磷脂代謝如何變化鮮見報道。本研究以‘元帥’蘋果為試材,觀察P.expansum接種后病斑直徑的變化,測定侵染對果實細胞膜透性的影響,分析膜磷脂代謝關鍵酶活性和底物產物含量的變化,以期為揭示真菌侵染對果實細胞膜磷脂代謝的原理提供理論依據。
1 材料與方法
1.1 材料與試劑
供試‘元帥’(Malus domesticaL.cv.Delicious)商業成熟果實于2018年10月采自甘肅省條山集團景泰縣果園。選取外觀整齊、大小一致、無病蟲傷和機械傷的果實,單果套網套后裝入瓦楞紙包裝箱,于當天運抵甘肅農業大學食品科學與工程學院采后生物與技術實驗室,在常溫(22±2)℃、相對濕度50%~60%下貯藏待用。
P.expansum(T01)菌株由中國科學院植物學研究所果實逆境應答及生物控制技術研究組提供,PDA上保存待用。
PC、PI、PA、磷脂酶A2(phospholipase A2,PLA2)、磷脂酶C(phospholipase C,PLC)、PLD試劑盒 上海酶聯生物科技有限公司;棕櫚酸、硬脂酸、油酸、亞油酸、亞麻酸(標準品) 美國Sigma-Aldrich公司。
1.2 儀器與設備
LDZX-30KBS型立式壓力蒸汽滅菌鍋 上海申安醫療器械廠;SW-CJ-2FD型超凈工作臺 蘇凈集團蘇州安泰空氣技術有限公司;DHP-9082型恒溫培養箱 上海一恒科學儀器有限公司;A11 basic S025型研磨機 廣州艾卡儀器設備有限公司;DW-HL218型超低溫冰箱 中科美菱低溫科技有限公司;DDS-307A型電導率儀 上海儀電科學儀器股份有限公司;H2050R型臺式高速冷凍離心機 長沙湘儀離心機有限公司;UV-2450紫外-可見光分光光度計 日本島津公司;450-GC氣相色譜儀美國VARIAN公司。
1.3 方法
1.3.1 果實損傷接種
孢子懸浮液配制參照Wang Kaituo等[15]的方法。取25 ℃下培養7 d的P.expansum一皿,加入10 mL無菌水,用無菌涂布器輕輕刮下培養基表面的真菌孢子,經4 層紗布過濾后,將濾液轉入50 mL三角瓶中,加入25 μL體積分數0.05% Tween-80后搖勻,再漩渦振蕩數秒,使孢子分布均勻,采用血球計數板法計算出孢子濃度,最終將孢子懸浮液稀釋至1.0×106個孢子/mL。
損傷接種參照Ge Yonghong等[16]的方法。果實先用自來水沖洗干凈,然后用體積分數為1%的次氯酸鈉消毒2 min,再用清水沖洗晾干。用體積分數70%的乙醇棉球對果實表面消毒,稍作晾干后,用滅菌鐵釘于赤道部位對稱打孔2 個,孔內分別接入20 μL上述孢子懸浮液(接種組)和20 μL無菌水(對照組)。稍作晾干裝入聚乙烯薄膜袋中,于室溫((22±2)℃、相對濕度85%~90%)條件下貯藏待用,于貯藏的第0、1、2、4、6天測定相關指標。每處理用果實30 個,重復3 次。
1.3.2 病斑直徑測定
采用十字交叉法測定病斑直徑,每處理用果實3 個,重復3 次。
1.3.3 細胞膜透過率測定
細胞膜透過率測定參照Ren Yalin等[17]的方法。用直徑10 mm打孔器打取果實病斑邊緣外以外12 mm以內健康組織,切取皮下2~5 mm處果肉組織圓片3.0 g,用去離子水將其沖洗至少3 次,用濾紙吸干多余水分,放入盛有30 mL去離子水的燒杯中,用電導率儀分別測初始和室溫條件下放置3 h后的電導率。然后將燒杯及組織置于95 ℃水浴鍋中水浴30 min,待冷卻至室溫后測定最終電導率。細胞膜透過率用公式(1)計算。

1.3.4 生化指標測定取樣
樣品前處理參照Gong Di等[4]的方法并作修改。用無菌手術刀切取果實病斑以外12 mm以內健康部位果皮及其皮下2~4 mm組織,液氮速凍后用研樣機研為粉末,裝入50 mL離心管,于-80 ℃超低溫冰箱中保存待用。
1.3.5 磷脂酶活力的測定
粗酶液的提取參照Alferez等[18]的方法并略修改。取0.5 g冷凍粉末,加入4.5 mL 0.1 mol/L pH 7.4的磷酸緩沖液,振蕩混勻,4 ℃、3 000×g離心15 min,上清液即為粗酶液。PLA2、PLC、PLD活力均采用相關試劑盒說明書的方法進行測定,單位均為U/g。
1.3.6 PC、PI和PA含量的測定
磷脂的提取參照許佳妮等[19]的方法并略修改。取0.1 g冷凍粉末,加入1 mL氯仿/甲醇溶液(2∶1,V/V)超聲提取30 min后,室溫下靜置30 min,在4 ℃、3 000×g下離心20 min,取上清液待測。
PC、PI、PA含量測定均參照相應試劑盒方法,單位均為μg/g。
1.3.7 脂肪酸含量的測定
脂肪酸含量測定參照胡妍蕓等[20]的方法并略修改。取冷凍粉末2.0 g,加入5 mL石油醚-乙醚(4:3,V/V)混合液在0~4 ℃下浸提24 h,再加入5 mL 0.4 mol/L的氫氧化鉀-甲醇溶液,室溫下甲酯化2 h,重蒸餾水分離得到有機相,氮氣吹干,1 mL氯仿溶解,無水硫酸鈉除水過0.45 μm膜,裝入進樣小瓶中用氣相色譜儀分析。色譜柱為極性柱CP-WAX(30 m×0.32 mm,0.5 μm);升溫程序:120 ℃下保持2 min,以6 ℃/min升至2 3 0 ℃,保持1 0 m i n;載氣(高純N2)流速2 mL/min,進樣量1 μL;分流比為20∶1,進樣口和檢測器溫度均為250 ℃。在相同條件下,將一定質量濃度的不同脂肪酸標品和待測樣品分別依次進樣,通過比較保留時間進行定性,通過峰面積歸一化法定量。脂肪酸不飽和度利用公式(2)計算。

1.4 數據統計與分析
以上每項指標測定均重復3 次。全部數據用Microsoft Excel 2010軟件計算平均值和標準偏差,采用Origin 9.0軟件作圖,利用SPSS 19.0軟件進行Duncan’s多重差異顯著性分析和Pearson相關分析(P<0.05)。
2 結果與分析
2.1 P.expansum侵染對果實病斑直徑的影響
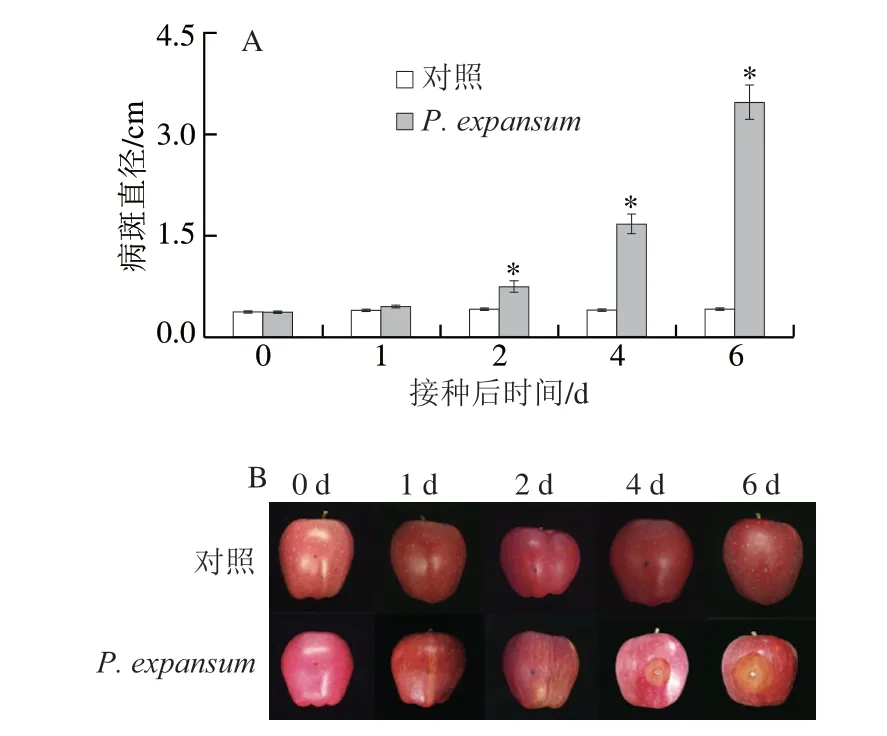
圖1 P.expansum侵染后蘋果果實病斑直徑(A)和外觀(B)Fig.1 Effect of P.expansum infection on lesion diameter (A)and appearance (B) of apple fruits
由圖1可知,P.expansum侵染后果實病斑直徑逐漸增大,而對照果實的病斑直徑基本保持穩定。侵染果實的病斑直徑在前2 d與對照相比無顯著差異,2 d后均顯著高于對照組,第4天時高出對照組3.19 倍(圖1)。上述結果表明,果實和病原菌互作早期(前2 d)果實抗性占優勢,而2 d后病原菌的致病性明顯提高,病斑明顯增大。
2.2 P.expansum侵染對果實細胞膜透過率的影響
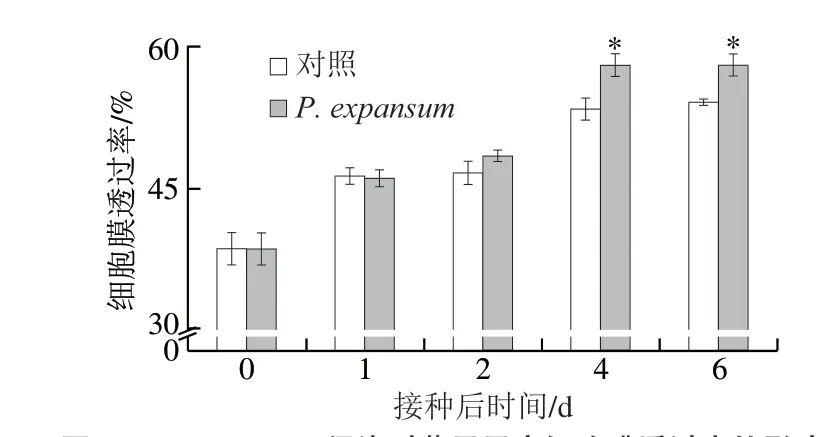
圖2 P.expansum侵染對蘋果果實細胞膜透過率的影響Fig.2 Effect of P.expansum infection on cell membrane permeability of apple fruits
細胞膜透過率是反映果實細胞膜完整性的重要指標,病原菌侵染會破壞果實細胞膜的完整性[21]。無論是接種組還是對照組,果實細胞膜透過率整體均呈上升趨勢,侵染早期(前2 d)果實細胞膜透過率與對照組無明顯差異,而侵染后期(2 d后)顯著高于對照組(P<0.05)(圖2)。由此表明,P.expansum侵染早期對細胞膜透過率影響不大,但在侵染后期明顯破壞了細胞膜的完整性。
2.3 P.expansum侵染對果實膜磷脂代謝的影響
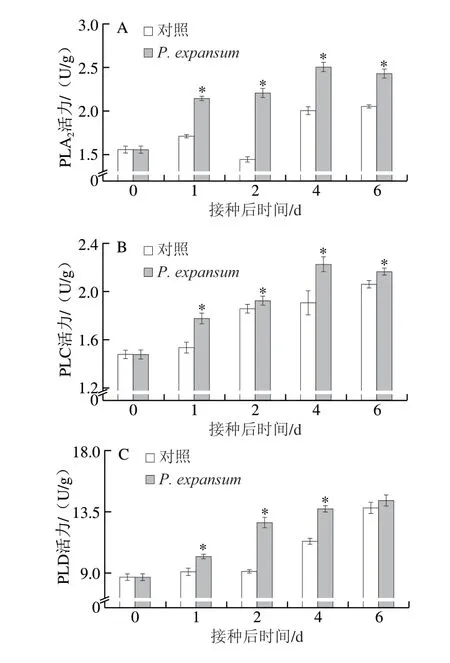
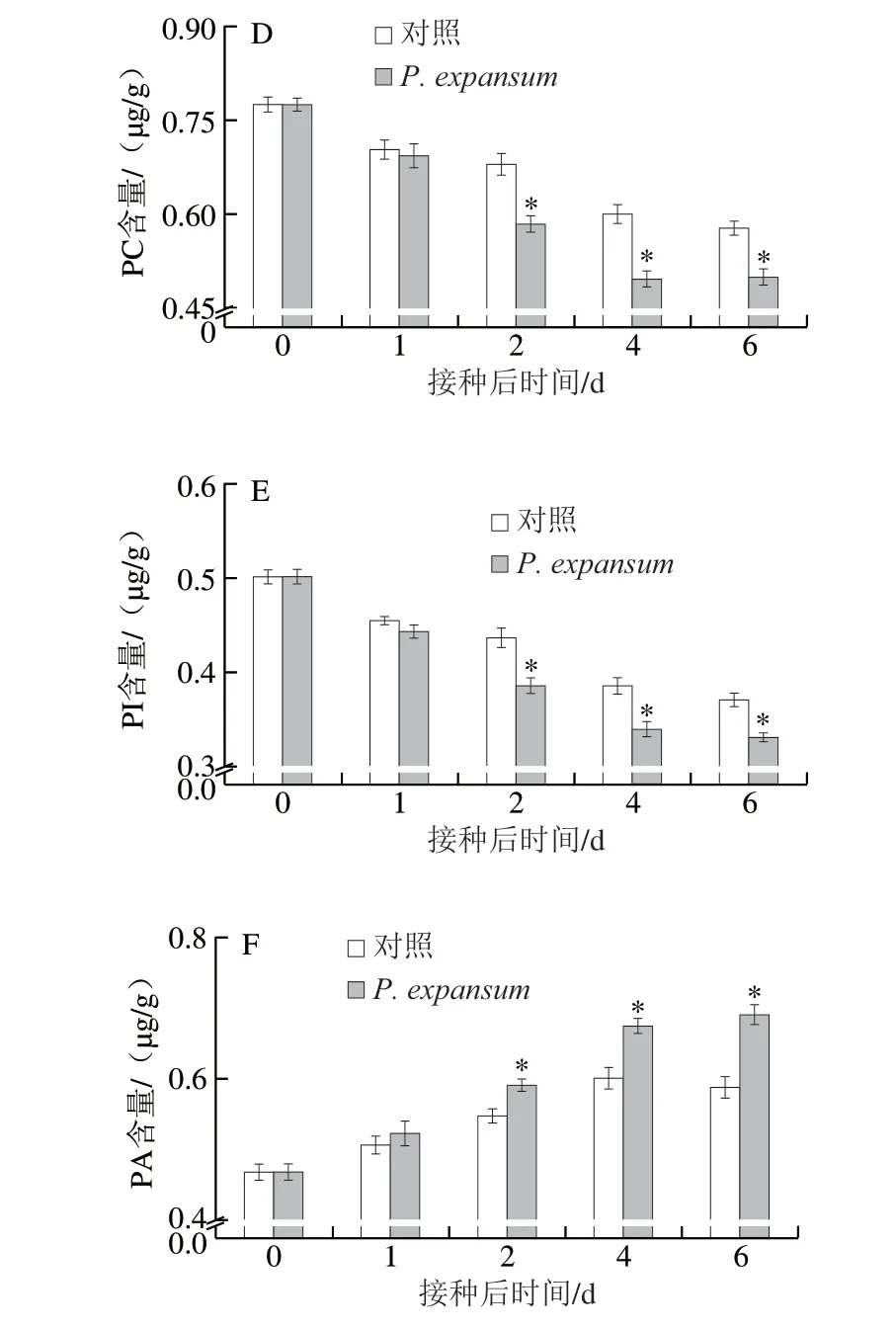
圖3 P.expansum 侵染對蘋果果實PLA2(A)、PLC(B)、PLD(C)活力以及PC(D)、PI(E)和PA(F)含量的影響Fig.3 Effect of P.expansum infection on the activities of PLA2 (A),PLC (B) and PLD (C) and the contents of PC (D), PI (E) and PA (F)in apple fruits
PLA2、PLC和PLD是降解膜磷脂的主要酶,PLA2專一性水解PC的sn-2位酯鍵,釋放脂肪酸;PLC可專一性水解PI產生三磷酸肌醇和二酰甘油;PLD可將PC水解為PA和膽堿[22]。由圖3可知,蘋果果實的PLA2活力在侵染后先持續增高,第4天時達到峰值,再小幅下降。對照果實的PLA2活力先小幅升降,后明顯增高。侵染果實的PLA2活力均顯著高于對照組,第2天時高于對照組52.46%(P<0.05)(圖3A)。侵染果實和對照果實的PLC活力整體上均呈逐漸升高趨勢,但侵染果實PLC活力顯著高于對照組,第4天時侵染果實PLC活力高于對照組16.36%(P<0.05)(圖3B)。侵染果實PLD活力持續升高,對照果實PLD活力2 d前基本保持穩定,2 d后逐漸升高。侵染果實PLD活力顯著高于對照組,其中第2天達到最大差異且高于對照組39.28%(P<0.05)(圖3C)。侵染和對照果實的PC和PI含量均逐漸降低,侵染果實的PC和PI含量在早期(2 d前)與對照組無顯著差異,后期(2 d后)顯著低于對照組,第4天時分別低于對照組16.91%和11.8%(P<0.05)(圖3D、E)。侵染和對照果實的PA含量均逐漸升高,侵染果實的PA含量早期(2 d前)與對照組無顯著差異,后期(2 d后)顯著高于對照組,第4天時高出對照組12.74%(P<0.05)(圖3F)。上述結果表明,P.expansum侵染伊始,果實的PLA2、PLC和PLD活力就快速升高,但侵染果實的PC、PI和PA含量在侵染后期(2 d后)才顯著低于或高于對照組。
2.4 P.expansum侵染對果實脂肪酸含量和脂肪酸不飽和度的影響
較高的不飽和脂肪酸含量對維持果實的細胞膜流動性和完整性具有重要作用[23]。此外,細胞膜不飽和脂肪酸含量越高,果實的抗病性也越強[24]。侵染果實和對照果實油酸、亞油酸、亞麻酸含量均在貯藏初期迅速升高,在第1天時達到峰值,然后逐漸降低。在貯藏的第2天,侵染果實的油酸、亞油酸和亞麻酸分別比對照分別高31.58%、55.57%和43.67%(P<0.05)(圖4A、B、C)。侵染果實棕櫚酸和硬脂酸含量在貯藏過程中逐漸升高,且在后期升高的速率加快,對照果實的含量在侵染早期(前2 d)保持穩定,后期(2 d后)略有升高。侵染果實的棕櫚酸和硬脂酸含量在第6天時分別高出對照組49.46%和43.39%(P<0.05)(圖4D、E)。侵染和對照果實的脂肪酸不飽和度整體呈先升高后降低的趨勢,侵染早期(前2 d)果實的脂肪酸不飽和度顯著高于對照組,第2天時高于對照組37.54%。侵染后期(4 d后)顯著低于對照組,第6天時低于對照組16.26%(P<0.05)(圖4F)。上述結果表明,P.expansum侵染早期促進了果實不飽和脂肪酸的釋放,而侵染后期促進了飽和脂肪酸釋放,早期的不飽和脂肪酸釋放與果實抗病性相關,而后期的飽和脂肪酸釋放則與P.expansum的致病相關。
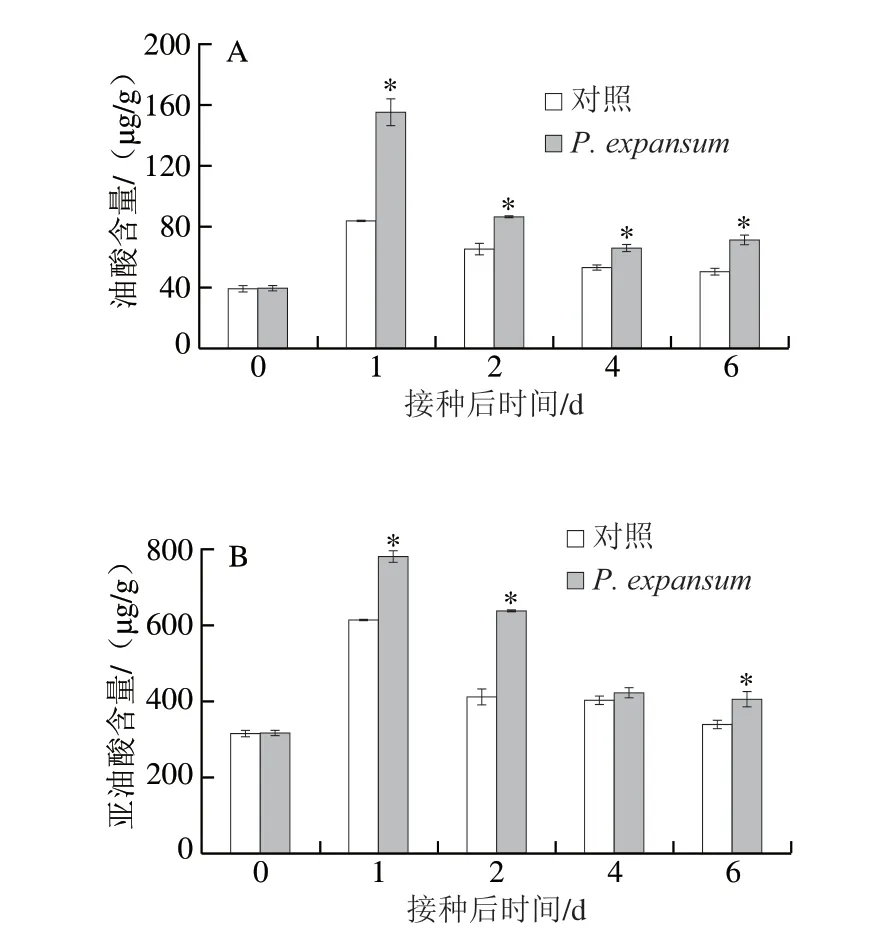
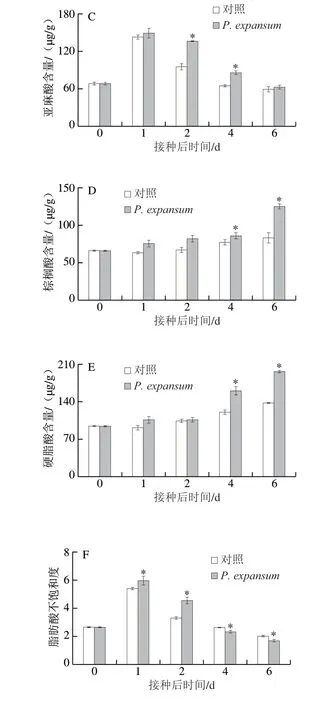
圖4 P.expa nsum侵染對蘋果果實油酸(A)、亞油酸(B)、亞麻酸(C)、棕櫚酸(D)、硬脂酸(E)含量和脂肪酸不飽和度(F)的影響Fig.4 Effect of P.expansum infection on the contents of oleic acid (A),linoleic acid (B), linolenic acid (C), palmitic acid (D), stearic acid (E)and the degree of unsaturation of fatty acids (F) in apple fruits
2.5 P.expansum侵染果實細胞膜透率與膜磷脂代謝相關指標的相關性
由表1可見,P.expansum侵染果實細胞膜透過率與PLA2、PLC和PLD活力呈極顯著正相關,相關系數分別為0.938、0.989和0.953;與PC和PI含量呈極顯著負相關,相關系數分別為-0.949和-0.955;與PA含量極顯著正相關,相關系數為0.985。此外,侵染果實細胞膜透率還與棕櫚酸和硬脂酸含量顯著極正相關,相關系數分別為0.781和0.901。
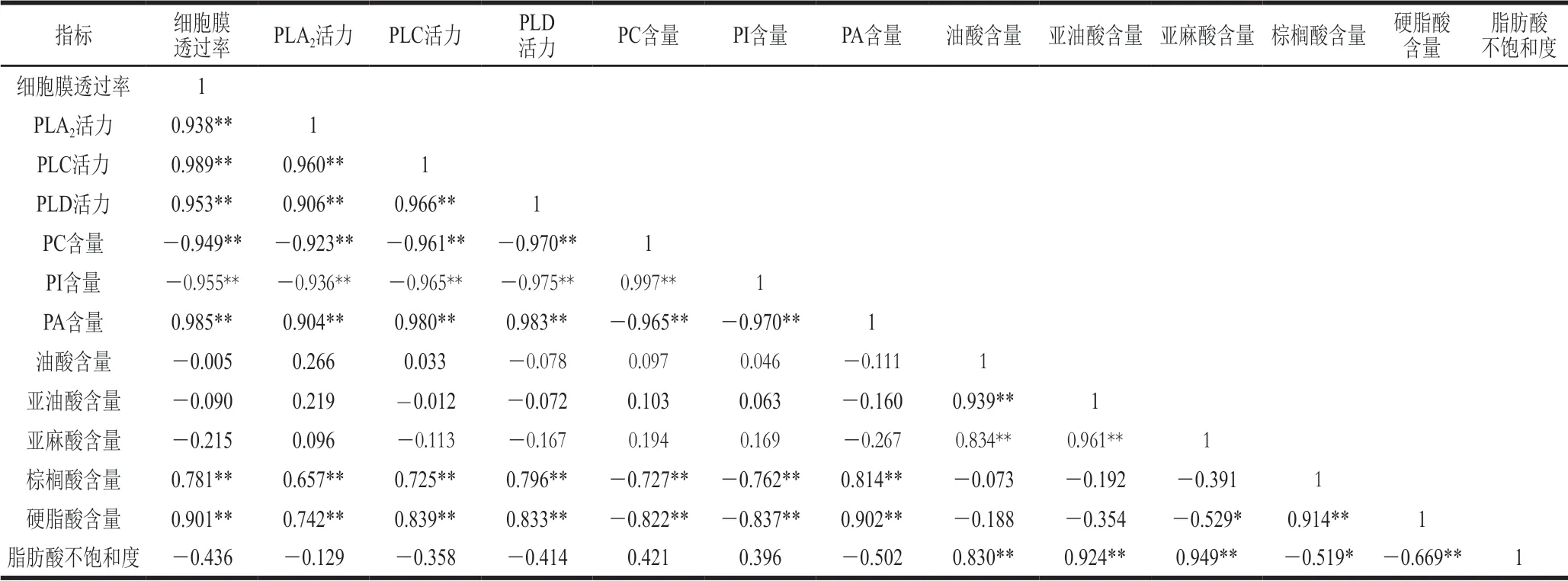
表1 P.expansum侵染果實細胞膜透過率與膜磷脂代謝各指標間的相關性分析(n=15)Table 1 Correlation analysis between cell membrane permeability and phospholipid metabolism indices in apple fruits infected by P.expansum (n= 15)
3 討 論
本研究發現,P.expansum侵染果實的病斑直徑在侵染早期(前2 d)無明顯變化,侵染后期(2 d后)快速升高(圖1)。由此表明,侵染早期果實對P.expansum侵染表現抗性,而侵染后期果實抗性減弱,病原菌的致病性明顯提高。蘋果果實早期對P.expansum的抗性涉及活性氧、抗菌物質和病程相關蛋白等諸多方面[25]。而P.expansum對蘋果的致病性主要源自細胞壁降解酶[2]。本研究中侵染后期P.expansum顯著提高蘋果果實細胞膜透過率的結果(圖2)與P.longanae侵染龍眼和尖孢炭疽菌(Colletotrichum acutatum)侵染番石榴后的現象[26-27]類似。果實細胞膜透率的增大一方面是因為侵染導致活性氧大量積累,破壞了細胞膜的完整性[28];另一方面是因為侵染促進果實乙烯釋放[29],進而能誘導磷脂酶活力增加,加速膜磷脂降解[30]。
侵染早期,果實PLA2活力快速升高(圖3A)。該結果與晚疫病菌(Phytophthora infestans)侵染馬鈴薯塊莖后PLA2活力增加的結果[31]類似。侵染果實細胞膜透率與PLA2活力之間呈極顯著正相關,表明PLA2在分解膜磷脂中具有重要作用。PLA2是甘油磷脂sn-2位酯酰基水解酶,能夠水解產生油酸、亞油酸和亞麻酸等不飽和脂肪酸。關于真菌侵染如何誘導果實PLA2活力升高的機制研究不多。由于P.expansum侵染會促進蘋果的乙烯釋放[4],推測乙烯可能參與了PLA2活力的誘導。P.expansum侵染早期油酸、亞油酸和亞麻酸迅速釋放(圖4A~C),該結果與P.litchii侵染荔枝觀察到的不飽和脂肪酸含量升高結果[11]基本一致。有研究表明,較高油酸、亞油酸和亞麻酸含量有利于維持獼猴桃和柑橘果實細胞膜的完整性[23,32-33]。此外,不飽和脂肪酸組成和含量還與枇杷和荔枝果實的抗病力密切相關[24,34]。Feussner等[13]發現,不飽和脂肪酸可通過脂氧合酶途徑生成茉莉酸及其衍生物,進而誘導植物抗病力。蘋果果實的棕櫚酸和硬脂酸含量在侵染后期迅速升高(圖4D、E),該結果與P.litchii侵染荔枝果實后觀察到的現象[11]基本類似。侵染果實的細胞膜透率與飽和脂肪酸含量極顯著正相關。飽和脂肪酸的釋放是磷脂酶A1(phospholipase A1,PLA1)作用的結果[35],但PLA1如何在侵染果實中發揮作用尚有待進一步研究。
果實的PLC活力在P.expansum侵染期間持續升高(圖3B)。該結果與葉霉病菌(Cladosporium fulvum)侵染番茄葉片后PLC 4和PLC 6基因表達量升高的結果[36]類似。PLC是一種質膜蛋白,非特異性PC-PLC可水解PC生成二酰甘油(diacylglycerol,DAG)和磷酸膽堿,特異性PI-PLC水解4,5-二磷酸肌醇生成1,4,5-三磷酸肌醇和DAG[36]。DAG可通過DAG激酶磷酸化為PA[37]。P.expansum侵染果實PLD活力持續力升高(圖3C)。該結果與L.theobromae侵染的龍眼果實PLD活力升高結果[9]基本類似。PLD能夠專一力水解結構磷脂PC,生成PA和膽堿陽離子自由基團,其活力決定了磷脂的水解強度[38]。乙烯能夠誘導番茄果實的PLC和PLD轉錄及活力升高[39-40],表明P.expansum侵染蘋果期間PLC和PLD激活可能被乙烯促進。此外,病原菌侵染早期激活的PLD還參與了擬南芥、水稻和番茄的防御反應[41]。由于侵染果實的PC和PI含量分別與PLA2、PLC和PLD活力呈極顯著負相關,表明PLA2、PLC和PLD在分解膜磷脂中作用突出。PC是一種主要的膜磷脂,與其他脂質如PI等共同維持細胞膜結構[42]。P.expansum侵染導致果實貯藏后期PC和PI含量快速降低(圖3D、E),該結果與L.theobromae侵染龍眼果實后觀察到的結果[9]基本一致。細胞膜磷脂可在果實抵御病原菌侵染過程中提供物質和能量,增強抗病力[43]。PA作為PC、PI的部分水解產物,在侵染后期含量快速升高(圖3F)。前人研究表明,PA也是植物中重要的脂質信號分子,被稱為“脂質第二信使”,參與多種脅迫相關的信號傳導[44]。據此推測,PA可能在果實抗病性中發揮重要作用。
4 結 論
P.expansum侵染蘋果早期(前2 d),果實病斑直徑無明顯變化,細胞膜透過率與對照組無顯著差異,侵染后期(2 d后)果實病斑直徑和細胞膜透過率明顯增大。P.expansum侵染激活了果實的PLA2、PLC和PLD活力,加速了膜磷脂降解,破壞了細胞膜的完整性。侵染早期促進了果實不飽和脂肪酸的釋放,侵染后期促進了飽和脂肪酸的釋放,侵染早期不飽和脂肪酸的迅速釋放可能與果實抗性相關。
