基于表面等離子體共振(SPR)技術實時分析RGD基序與整聯蛋白的相互作用
2021-03-01 09:08:22李鵬飛龔真莉杜曉華
畜牧獸醫學報 2021年1期
李鵬飛, 龔真莉,杜曉華,蔣 韜*
(1.甘肅農業大學 動物醫學院,蘭州 730070;2.中國農業科學院 蘭州獸醫研究所 家畜疫病病原生物學國家重點實驗室國家口蹄疫參考實驗室,蘭州 730046)
口蹄疫(foot-and-mouth disease,FMD)分布于世界各地,是一種具有高度傳染性的病毒性疾病[1-2],因其具有重大的經濟影響,被認為是影響畜牧業發展最重要的疫病之一。口蹄疫病毒(foot-and-mouth disease virus,FMDV)是引起FMD的病原體,屬于小RNA病毒科,口蹄疫病毒屬,由O、A、C、SAT1、SAT2、SAT3和Asia1 7種血清型組成[3]。FMD完整的病毒粒子為正二十面體對稱結構, 由VP1、VP2、VP3、VP44種結構蛋白各60個拷貝組成, RNA分子被包裹在內[4]。VP1~VP3 3種結構蛋白暴露在病毒顆粒表面且聚集成蛋白質三聚體復合物,較小的結構蛋白VP4隱藏于內部[5]。VP1蛋白是FMDV顆粒表面的主要結構蛋白,在其易變的 G-H 環上不僅含有病毒主要的抗原位點,而且含有一個高度保守的RGD基序(精氨酸-甘氨酸-天門冬氨酸),該基序是病毒粒子與細胞表面整聯蛋白結合引起感染的主要結合位點。盡管RGD依賴的整聯蛋白受體通過RGD基序識別配體,但FMDV-VP1蛋白G-H環中VP1側面的氨基酸殘基在FMDV感染中同樣起重要作用。如RGD依賴性整聯蛋白αvβ5和α5β1并未充當FMDV的受體[6]。VP1 S154D突變導致Asia1 型FMDV使用豬整聯蛋白受體αvβ6和αvβ8的能力增加,從而增加了其在宿主細胞中的復制水平,增強了對宿主豬的致病性[7]。
整聯蛋白是哺乳動物細胞黏附和信號傳導的關鍵蛋白之一[8]。迄今為止,已鑒定18種不同的α亞基和8種β亞基非共價結合構成24種整聯蛋白異二聚體跨膜糖蛋白受體,可與多種細胞或細胞外配體結合,通常整聯蛋白不具有活性,在活化后才能成為黏附劑[9]。整聯蛋白可以通過結合細胞外配體或通過細胞內結構域與細胞骨架相互作用,在細胞膜上雙向傳遞信號[10]。在非活性狀態下,整聯蛋白的胞外結構域不與配體結合,以彎曲構象形式存在。然而,來自細胞的信號引起構象變化,暴露外部配體結合位點,結合配體并將信號從細胞外傳遞到細胞內[11]。在體外,至少有4種整聯蛋白(αvβ1、αvβ3、αvβ6和αvβ8)可作為FMDV受體來介導FMDV感染宿主細胞[12]。研究發現,整聯蛋白αvβ6可能是決定FMDV 組織嗜性的主要受體,且FMDV可能會激發整聯蛋白αvβ6在上皮細胞中的表達,然后以網格蛋白依賴的方式內化。表明整聯蛋白αvβ6不僅是FMDV的吸附受體,而且在病毒與受體結合后的脫殼、復制等過程中也發揮著重要作用[6]。研究證實,可溶性截短形式的牛整聯蛋白αvβ6通過二價陽離子依賴的方式結合FMDV-RGD,可用作FMDV的捕獲試劑,并不影響病毒的結合[13]。除整聯蛋白外,在細胞培養中,FMDV可以迅速適應利用替代細胞受體: 硫酸乙酰肝素(HS)[14]和尚未確認的替代受體[15]。研究證實,整聯蛋白α 和 β 亞基 N-末端形成的球形區域為胞外配體結合域[16]。
表面等離子體共振(surface plasmon resonance,SPR)技術是一種基于光學的實時監測方法,且因SPR技術無需標記、靈敏度高、特異性高、實時和快速等優勢被廣泛應用于各種生物分子相互作用的研究[17]。目前,尚未見國內外采用SPR技術研究FMDV RGD基序與豬源整聯蛋白αvβ6胞外區結構域、αv亞基胞外區結構域和β6亞基胞外區結構域的結合動力學方面的報道。本研究在國內首次利用基于SPR技術的Biacore 3000系統,研究了FMDV RGD基序與豬源整聯蛋白αvβ6胞外區結構域、αv亞基胞外區結構域和β6亞基胞外區結構域的相互作用,研究結果將為進一步闡明FMDV與宿主細胞整聯蛋白受體的相互作用機制提供依據。
1 材料與方法
1.1 材料和儀器
豬源整聯蛋白αvβ6胞外區、αv亞基胞外區和β6亞基胞外區由本實驗室原核表達并純化、鑒定得到。RGD(Arg-Gly-Asp)由南京金斯瑞生物科技有限公司合成。Biacore 3000生物傳感器;Biacore 3000控制軟件;氨基偶聯試劑盒;CM5 Sensor Chip(CM5傳感器芯片);不同 pH (4.0、4.5、5.0 和 5.5)的10 mmol·L-1醋酸鈉緩沖液;HBS-EP 緩沖液;再生試劑盒均購自GE Healthcare公司。
1.2 方法
1.2.1 芯片表面預處理 開機后,首先將系統溫度設置為25 ℃,取出Maintaince 芯片,插入 CM5 芯片并固定,使用primer命令用純水沖洗管道,再換HBS-EP緩沖液,再次使用primer命令沖洗管道,最后通過HBS-EP緩沖液,直到基線穩定,為下一步的試驗做好準備。
1.2.2 偶聯pH的選擇 分別使用氨基偶聯試劑盒中pH4.0、4.5、5.0、5.5的10 mmol·L-1醋酸鈉緩沖液稀釋目的蛋白樣品整聯蛋白αvβ6胞外區、αv亞基胞外區和β6亞基胞外區至20 μg·mL-1,使其帶上不同量的電荷。再將整聯蛋白樣品以10 μL·min-1的流速流過芯片表面,注射2 min,通過整聯蛋白樣品與芯片的結合曲線確定合適的偶聯pH。最后,以20 μL·min-1流速注射30 μL 50 mmol·L-1NaOH洗脫液完全去除芯片表面吸附的樣品蛋白。
1.2.3 在CM5芯片上偶聯整聯蛋白 使用預處理篩選得到的最合適的pH偶聯緩沖液稀釋目的蛋白至20 μg·mL-1,應用蛋白固定自動化程序進行氨基偶聯。
1.2.4 結合試驗 將RGD基序溶解在HBS-EP運行緩沖液中,并稀釋至低濃度(31.25 μg·mL-1)和高濃度(500 μg·mL-1),以10 μL·min-1的流速進樣3 min,觀察響應值,看是否有結合。每次進樣結束后,用Glycin-HCl(pH 3.0)再生30 s。
1.2.5 動力學測定 首先,將RGD樣品在HBS-EP緩沖液中稀釋至500、250、125、62.5、31.25 μg·mL-1的系列濃度。然后,以最大稀釋量開始,以最小稀釋量結束,分別在整聯蛋白固定表面和參考表面以30 μL·min-1的流速注射稀釋的RGD樣品3 min。最后,停止注射RGD樣品,觀察其在5 min內的解離率,在此期間芯片上只通過HBS-EP緩沖液。每個濃度的測定設置2個循環,在每個結合循環后,以30 μL·min-1的注射速度注入Glycin-HCl(pH 3.0),對芯片表面進行再生。
1.2.6 數據分析 將所得的實時SPR傳感圖導入Biaevaluation 分析軟件,選擇1∶1 Lamgmuir 結合模型進行局部擬合。
2 結 果
2.1 芯片表面預濃縮-偶聯pH的選擇
通過Biacore 3000生物傳感器信號響應值的變化,根據預濃縮得到的結果選擇最合適的偶聯pH。結果表明,整聯蛋白αvβ6胞外區、αv亞基胞外區和β6亞基胞外區均在pH4.0的偶聯緩沖液中與芯片表面產生最佳的靜電相互作用。
2.2 在CM5芯片上偶聯整聯蛋白
通過蛋白固定自動化氨基偶聯程序,最終得到樣品蛋白的實際偶聯量:整聯蛋白αvβ6胞外區為13 333.3 RU、αv亞基胞外區為15 000.8 RU、β6亞基胞外區為10 000.6 RU。
2.3 結合試驗與動力學測定
結合試驗RU值表明RGD溶液與整聯蛋白αvβ6胞外區和αv亞基胞外區有結合,且在低濃度下結合量少,在高濃度下結合量多,而RGD與β6亞基胞外區在兩種濃度下均基本沒有結合。由動力學曲線RU值可知,隨著反應質量濃度的增加,結合信號也逐漸升高,說明結合量增大,且都表現為快結合快解離(圖1)。采用 1∶1 Lamgmuir 結合模型擬合曲線,得到RGD基序與豬源整聯蛋白αvβ6胞外區結構域和αv亞基胞外區的結合動力學常數(表1)。由平衡解離常數可知,RGD基序與整聯蛋白αvβ6胞外區結構域的親和力比與整聯蛋白αv亞基胞外區結構域的親和力高。
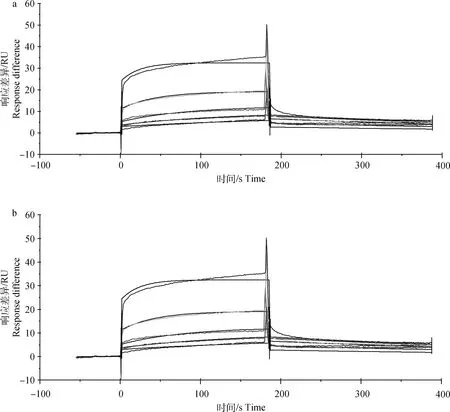
a.整聯蛋白αvβ6胞外區;b.整聯蛋白αv亞基胞外區a. αvβ6 extracellular domain; b. αv subunit extracellular domain圖1 RGD基序與整聯蛋白αvβ6胞外區和整聯蛋白αv亞基胞外區的結合動力學曲線Fig.1 kinetic curves of RGD motif binding to the αvβ6 extracellular domain and αv subunit extracellular domain

表1 RGD基序與整聯蛋白αvβ6胞外區和整聯蛋白αv亞基胞外區相互作用的動力學常數Table 1 Kinetic constants of the interaction between the RGD motif and the αvβ6 extracellular domain and αv subunit extracellular domain
3 討 論
FMDV通過細胞受體介導的內吞作用進入宿主,這一過程是由病毒與細胞表面受體(主要是整聯蛋白)的結合啟動的[18]。研究證實,在體外病毒通過結構蛋白VP1上的RGD基序與細胞表面整聯蛋白結合并誘導內化[19]。另外,研究發現,形成FMDV病毒顆粒的衣殼蛋白中氨基酸殘基的變化不僅會影響病毒與細胞受體之間的吸附能力,還可能影響病毒的生物學特性[7,20-22]。
本研究結果表明,RGD基序與整聯蛋白β6亞基胞外區結構域之間幾乎沒有結合;RGD基序與整聯蛋白αvβ6胞外區結構域的親和力最強,平衡解離常數KD為7.33×10-6M;與整聯蛋白αv亞基胞外區結構域之間的親和力小于與整聯蛋白αvβ6胞外區結構域的親和力,平衡解離常數KD為9.79×10-5M。結合常數表明RGD基序與整聯蛋白αvβ6胞外區結構域的結合速度快于與αv亞基胞外區結構域的結合速度。同時,動力學曲線表明,RGD基序與整聯蛋白αvβ6胞外區結構域和整聯蛋白αv亞基胞外區結構域的結合過程均為快結合快解離,有部分原因可能是由于本試驗使用的RGD基序缺少上下游氨基酸,同時在體外整聯蛋白構象改變,從而導致結合不穩定。
分子模擬FMDV RGD結構域的氨基酸與整聯蛋白氨基酸之間可能的相互作用,發現RGD結構域的精氨酸(Arg143)與家牛α-整聯蛋白的保守結構域(SIPLQ)的亮氨酸(Leu679)相互作用,除此之外其他氨基酸之間也存在相互作用,如FMDV與β-整聯蛋白之間的其他相互作用包括Asn47-Gln1710、Arg152-Asn1643、Gly202-Gln1187和 Ser154-Lys1669;對不同牛種α-整聯蛋白外顯子區體外擴增的研究表明,嗜性位點(SIPLQ)具有保守性[23]。在本研究中并未得到RGD基序與整聯蛋白β6亞基胞外區結構域之間的結合能力,可能是由于本研究中使用的是單一的Arg-Gly-Asp3肽,并沒有RGD上下游的氨基酸,因此,并沒有體現出與上述研究報道相似的結果。有研究表明,FMDV VP1衣殼蛋白中GH環的RGD序列可與細胞整聯蛋白形成極其穩定的復合物,并且該復合物對EDTA具有抗性,特定氨基酸的替換表明,與αvβ6結合的穩定性取決于靠近RGD序列的螺旋結構,RGD +1和RGD +4位點的兩個亮氨酸殘基是這種穩定相互作用的關鍵位置[24]。
此外,有研究報道,整聯蛋白對RGD基序的特異性識別是通過α亞基β螺旋槳結構域與精氨酸結合,天冬氨酸通過Mg2+或Mn2+在金屬離子結合位點(MIDAS)與整聯蛋白β亞基β1結構域結合[25-26]。本研究的樣品中并沒有加入Mg2+或Mn2+等金屬離子。研究證實,整聯蛋白β亞基βI結構域含有多個二價金屬陽離子結合位點(MIDAS、ADMIDAS、LIMBS),配體與整聯蛋白的結合受到二價金屬陽離子的調節,二價金屬陽離子(Ca2+、Mg2+和Mn2+)不僅是整聯蛋白與配體結合的前提,而且還可以影響整聯蛋白的激活狀態[17,27],同時,二價金屬陽離子對配體與αv整聯蛋白上RGD位點結合的影響在同一配體的整聯蛋白和同一整聯蛋白的配體之間變化[28]。本研究結果將為進一步闡明FMDV與宿主細胞整聯蛋白受體的相互作用機制提供依據。
4 結 論
利用SPR技術從動力學角度實時分析了RGD基序分別與豬源整聯蛋白αvβ6胞外區結構域、αv亞基胞外區結構域和β6亞基胞外區結構域的相互作用,通過數據擬合分析得到特異性相互作用的動力學常數,結果表明,SPR技術能夠實時、快速地檢測相互作用過程。
