玉米赤霉烯酮誘導山羊子宮內膜基質細胞凋亡的研究
2021-02-27 04:57:20王宗捷張瑞雪劉守勤靳亞平林鵬飛
畜牧獸醫學報 2021年2期
王宗捷,張瑞雪,劉守勤, 靳亞平,林鵬飛
(西北農林科技大學動物醫學院,楊凌 712100)
ZEN主要是由鐮刀菌屬物種經過聚合酮途徑產生,常存在于霉變的谷物上。ZEN化學性質穩定,因此在谷物的加工過程中不易被降解,動物常因食用污染了ZEN的霉變飼料而中毒[1]。ZEN和雌激素的化學結構具有相似性,因此,ZEN可以和內源性雌激素競爭性結合雌激素受體,引起體內生殖激素調節紊亂,導致雌性動物不孕、流產、死胎等[2]。研究顯示,低劑量的ZEN具有和雌激素相同的作用,兩者均能促進卵泡發育,但ZEN可引起PCNA、Bcl-2及Bax基因高表達,致使ZEN誘導的卵泡常常表現異常發育狀態[3-5]。ZEN對牛、豬和小鼠卵母細胞的減數分裂進程具有抑制作用,會引起卵母細胞成熟延遲以及染色質異常[6-8]。ZEN能夠增加小鼠胚胎的致死率[9]。近期的研究顯示,ZEN還能夠影響胚胎的植入過程[10]。
近年來,關于ZEN對哺乳動物產生毒性的作用機制研究也越來越多,用ZEN處理小鼠睪丸間質細胞,發現細胞死亡形式主要是細胞凋亡[11]。進一步研究發現,ZEN能夠通過多種途徑誘導生殖細胞的凋亡,ZEN可以通過激活Fas-FasL信號通路誘導大鼠支持細胞凋亡,還可以通過干擾基因組的穩定性促進顆粒細胞的凋亡[12-13]。當機體發生內質網應激(ERS)且穩態不能平衡時,細胞會激活內在的凋亡途徑,導致細胞凋亡[14]。ERS通過以下通路介導細胞的凋亡:1)PERK-ATF4-CHOP-Bcl-2-Bax通路導致細胞凋亡;2)IRE1α-JNK-Bcl-2通路誘導細胞調亡;3)ATF6-CHOP-Bcl-2通路介導細胞凋亡[15]。先前的研究揭示了ZEN通過內質網應激途徑調節大鼠睪丸支持細胞、TM4細胞以及牛乳腺上皮細胞的凋亡過程,表明ERS在ZEN引起的細胞凋亡中發揮重要的作用[16-18]。
雖然目前ZEN對動物生殖系統的影響已有很多報道,但是關于ZEN在母體子宮方面的報道仍不完善。子宮內膜細胞在保護母體不受病原體侵害以及胚胎附植方面都發揮了積極的作用。因此,本試驗選用山羊ESCs作為試驗材料,探究ZEN誘導細胞凋亡的相關信號通路,為研究ZEN在雌性哺乳動物中引起生殖毒性的機制提供依據。
1 材料與方法
1.1 試劑
玉米赤霉烯酮(Z2125)購自Sigma公司。胎牛血清(Z7186FBS-500)購自ZETA公司。RNA Trizol試劑(9109)購自TaKaRa公司。SYBR Green Master Mix定量試劑盒(Q311-02)購自諾唯贊公司。反轉錄試劑盒(AG11705)購自艾科瑞公司。BCA蛋白含量檢測試劑盒(KGP902)和全蛋白提取試劑盒(KGP2100)以及細胞凋亡試劑盒(KGA1017)購自凱基生物公司。CCK-8試劑盒(C0037)購自碧云天公司。β-actin抗體(JC-PA001)購自晶彩生物。CHOP抗體(2895)購自美國CST公司。GRP78抗體(ab21685)和IRE1抗體(ab124945)購自Abcam公司。ECL發光液(32109)購自賽默飛世爾公司。
1.2 細胞培養
永生化的山羊ESCs細胞系由本實驗室建立和保存[19]。將ESCs 細胞在37 ℃、5%CO2環境下培養于 DMEM/F-12 培養基中(添加10%的胎牛血清),當細胞貼壁生長融合度為80%~90%時進行傳代。將ESCs以3×105個·孔-1的密度接種于六孔板中,培養24 h后,加入0、10、20、30、40 μg·mL-1ZEN,處理24 h后,在倒置熒光顯微鏡下,觀察細胞形態。
1.3 CCK-8
將傳代細胞消化后,用移液器重懸混勻,之后對細胞進行計數。ESCs以5×103個·孔-1的細胞密度接種在96孔板中,注意每個孔重復6次;培養24 h 后,用0、5、10、20、30、40、60 μg·mL-1的ZEN處理24 h,之后將CCK8以10 μL·孔-1的量加入到細胞中;37 ℃孵育1 h后,通過酶標儀在450 nm處檢測吸光度值。2 μmol·L-14-PBA和10 μmol·L-1Irestatin9389 在ZEN處理前1 h加入。
1.4 細胞凋亡率檢測
用“1.2”中不同濃度的ZEN處理ESCs 24 h后,棄上清,并且用不含EDTA的0.25%的胰蛋白酶消化ESCs,輕輕吹打收集;ESCs懸液離心5 min,用冷的PBS洗滌兩次,將細胞密度調整至1×104個·mL-1;將細胞重懸于50 μL結合緩沖液中,加入5 μL的7-AAD并將混合物在黑暗中靜置15 min;加入450 μL結合緩沖液和1 μL的Annexin V-PE并在黑暗中溫育15 min;在1 h內進行流式細胞儀檢測。
1.5 Real-time PCR
用“1.2”中不同濃度的ZEN處理山羊ESCs 24 h后,棄掉上清,使用Trizol試劑提取細胞總RNA。cDNA合成按照反轉錄試劑盒說明書進行。在Bio-Rad CFX96 PCR儀中使用SYBR Green Master Mix進行Real-time PCR檢測。用 2-△△Ct法進行mRNA定量。β-actin為內參基因。引物序列詳見表1。

表1 Real-time PCR 引物序列
1.6 Western blot
用“1.2”中不同濃度的ZEN處理山羊ESCs 24 h 后,收集樣品。棄掉上清,用PBS清洗細胞兩次后,用細胞刷收集細胞,用1 mL的PBS洗滌、收集細胞;離心棄掉PBS后,加入蛋白裂解液,提取全蛋白,根據BCA試劑盒說明書來測定總蛋白的濃度;按照每個蛋白樣本30 μg加到凝膠電泳分離泳道中;然后,將目的蛋白樣品轉移到PVDF膜上,用10%脫脂牛奶封閉PVDF膜2 h;TBTS清洗膜之后,用β-actin(1∶5 000)、phosphoIRE1(1∶1 000)、GRP78(1∶1 000)、CHOP(1∶1 000)抗體分別4 ℃孵育過夜;TBST洗滌5次,5 min·次-1;在室溫下使其與辣根過氧化物酶標記二抗共同孵育1 h,TBST洗滌5次,5 min·次-1;最后,1∶1配制ECL發光液,均勻滴加到膜上,使用凝膠成像系統(Tannon Biotech)檢測。
1.7 數據統計與分析
所有的試驗至少重復3次。應用GraphPad Prism 8統計軟件對數據進行顯著性t檢驗和單因素方差(ANOVA)統計分析,得到的結果用“平均值± 標準差”表示,P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結 果
2.1 ZEN對山羊ESCs增殖和凋亡的影響
如圖1所示,隨添加ZEN濃度的升高,山羊ESCs的細胞活力逐漸下降。與對照組(0 μg·mL-1ZEN)相比,添加10 μg·mL-1ZEN顯著地抑制了山羊ESCs細胞活力(P<0.05),而20 μg·mL-1ZEN極顯著地抑制了山羊ESCs細胞活力(P<0.01)。當ZEN濃度大于40 μg·mL-1時,60%以上的ESCs增殖受到抑制。FCM檢測結果同樣證實,隨ZEN濃度的增加,ESCs細胞凋亡率顯著上升(表2)。40 μg·mL-1ZEN處理山羊ESCs 24 h后,50%左右ESCs發生了凋亡。
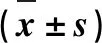
表2 ZEN對山羊ESCs細胞凋亡的FCM檢測結果
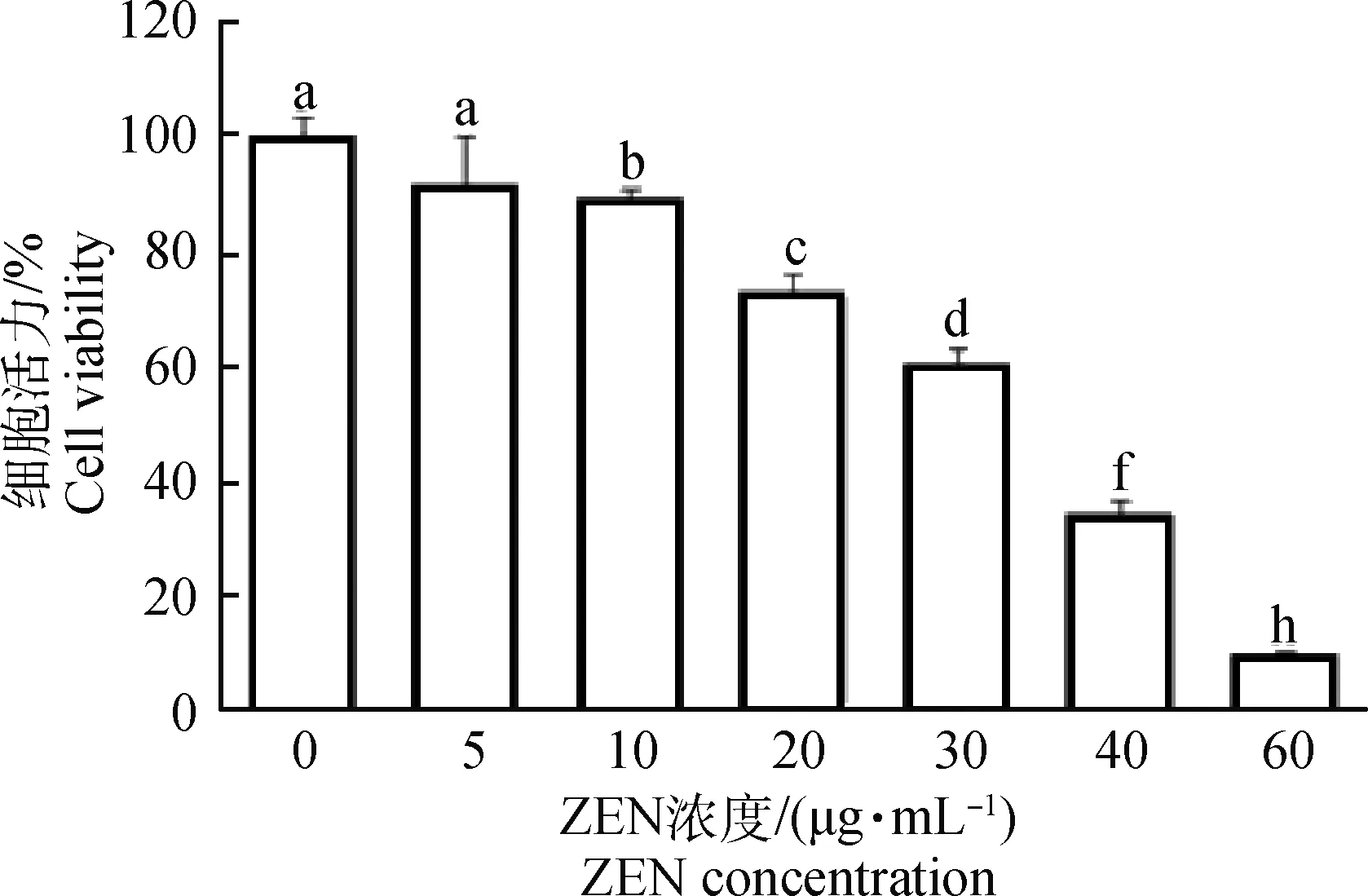
相鄰字母表示差異顯著,P<0.05;相隔兩字母表示差異極顯著,P<0.01。下同
2.2 ZEN對山羊ESCs凋亡相關基因的影響
圖2顯示,隨著ZEN濃度的升高,Caspase-9、Caspase-8和Bax/Bcl-2的mRNA水平表達量逐漸增加。30和40 μg·mL-1ZEN組顯著上調了Caspase-9 mRNA表達水平(P<0.05)。當添加10 μg·mL-1ZEN時,Bax/Bcl-2 mRNA表達水平即顯著增加;而30和40 μg·mL-1ZEN極顯著上調了Bax/Bcl-2 mRNA表達水平(P<0.01)。與對照組相比,10~30 μg·mL-1ZEN對Caspase-8 mRNA水平無顯著上調作用;當添加30 μg·mL-1ZEN時,Caspase-8 mRNA水平顯著增加(P<0.05)。

A.Caspase-9/β-actin; B.Caspase-8/β-actin; C.Bax/Bcl-2/β-actin
2.3 ZEN對ERS通路關鍵基因表達的影響
如圖3所示,添加30和40 μg·mL-1ZEN處理山羊ESCs 24 h后,GRP78 mRNA的表達水平顯著增加(P<0.05)。ZEN顯著上調了CHOPmRNA表達水平且呈劑量依賴性關系,10 μg·mL-1ZEN即顯著誘導了CHOPmRNA表達(P<0.05),而40 μg·mL-1ZEN極顯著上調了CHOPmRNA表達水平(P<0.01)。當ZEN濃度大于20 μg·mL-1時,ATF6 mRNA表達水平顯著增加(P<0.05)。10~30 μg·mL-1ZEN對PERKmRNA表達水平無顯著上調作用;當添加40 μg·mL-1ZEN時,PERKmRNA水平顯著增加(P<0.05)。而ZEN且呈劑量依賴性的顯著上調了IRE1 mRNA表達水平。Western blot結果進一步證實,添加10 μg·mL-1ZEN顯著增加了GRP78、CHOP和phosphoIRE1α蛋白表達水平(圖4,P<0.05)。

A.GRP78/β-actin; B.CHOP/β-actin; C.ATF6/β-actin; D.PERK/β-actin; E.IRE/β-actin

A.Western blot; B.GRP78; C.CHOP; D.phosphoIRE1
2.4 ERS對ZEN誘導ESCs凋亡中的調節作用
為了進一步分析ERS在ZEN引起的ESCs凋亡過程中的調控作用,首先采用ERS特異性抑制劑4-PBA和IRE1特異性抑制劑Irestatin9389預處理ESCs 1 h,添加30 μg·mL-1ZEN作用24 h后,CCK-8分析ESCs的增殖情況。圖5所示,添加4-PBA和Irestatin9389均顯著地挽救了ZEN對ESCs增殖的抑制作用(P<0.05)。

圖5 ERS特異性抑制劑對ZEN抑制ESCs增殖的調節作用
3 討 論
越來越多的研究表明,ZEN對雌性動物生殖系統具有顯著的毒性作用,ZEN可誘發斷奶小母豬子宮內膜和肌層增厚、腺體數量增多,導致子宮形態發生改變[20]。ZEN還增加了大鼠患多囊卵巢綜合征的風險[21]。還有研究證實,ZEN對小鼠的胎盤發育會產生不利的影響,還可導致胚胎早期附植失敗[22-23]。近期研究表明,ZEN可誘導不同時期小鼠子宮細胞發生凋亡,顯著上調凋亡關鍵基因Caspase-3、Caspase-8、Caspase-9以及Bcl-2的表達[24]。ZEN可以通過激活JNK/Drp1途徑刺激線粒體裂變從而降低人ESCs的存活和遷移活性[25]。目前,有關ZEN對反芻動物生殖毒性的報道較少,本試驗通過分析ZEN對山羊ESCs增殖和凋亡的影響,以期為揭示ZEN對山羊胚胎附植和妊娠的生殖毒性作用奠定基礎。
現已證實,ZEN可通過引起細胞凋亡進而引發生殖障礙。ZEN可通過激活JAK2/STAT3信號通路和抑制抗凋亡相關lncRNAs表達誘導豬顆粒細胞發生凋亡[26-27]。ZEN能夠通過激活線粒體途徑誘導大鼠睪丸支持細胞發生凋亡[28]。本試驗發現,ZEN對山羊ESCs的毒性作用呈現劑量依賴性的關系并能顯著誘發山羊ESCs凋亡;同時,凋亡基因Caspase-8、Caspase-9和Bax/Bcl-2 mRNA水平均被顯著上調。
先前的研究發現,ZEN可以通過內源性凋亡通路和線粒體凋亡通路誘導細胞凋亡[26,29]。本實驗室前期研究表明,ZEN可以通過ERS通路誘導小鼠卵巢顆粒細胞和巨噬細胞凋亡[30-31]。采用ERS抑制劑預處理細胞可顯著減輕ZEN誘導的人肝細胞凋亡和自噬[32]。當發生ERS時,機體會啟動一些反應來維持細胞穩態,當ERS超過內質網修復能力時就會啟動細胞凋亡機制。本研究發現,ZEN可以顯著上調ERS標志基因GRP78和CHOPmRNA和蛋白表達水平,并呈劑量依賴性。同時,ERS三條通路關鍵基因ATF6、PERK和IRE1的表達水平均顯著增加,尤其是IRE1通路上調最為顯著。添加ERS特異性抑制劑4-PBA以及IRE1特異性抑制劑Irestatin9389的結果進一步表明,ZEN可以通過ERS通路誘導ESCs發生凋亡。另外,ZEN具有類雌激素的毒性,它可以和內源性雌激素競爭性結合雌激素受體[33]。先前的研究表明,ZEN可以通過雌激素受體激活Wnt-1/β-catenin通路促進卵巢的發育[4]。ZEN可以通過調節雌激素受體信號和核受體Nur77的表達來抑制小鼠睪丸間質細胞中睪酮的生物合成[34]。山羊ESCs細胞中存在雌激素受體,ZEN是否通過雌激素受體對山羊ESCs產生毒性作用需要進一步的研究。
4 結 論
ZEN對山羊ESCs有顯著毒性作用,可劑量依賴性地抑制ESCs增殖和誘導ESCs凋亡;ZEN可以通過激活ERS通路誘導ESCs凋亡,且以IRE1通路最為顯著。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
學苑創造·A版(2015年11期)2016-01-14 09:03:27
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
七彩語文·畫刊(2012年4期)2012-04-29 00:00:00
