培養基配方對貝萊斯芽胞桿菌FJAT-45028脂肽產量和抑菌活性的影響
2021-02-26 11:13:20陳燕萍陳梅春藍江林鄭梅霞劉波王階平
福建農業科技 2021年12期
陳燕萍 陳梅春 藍江林 鄭梅霞 劉波 王階平


摘 要:探究培養基配方對貝萊斯芽胞桿菌FJAT-45028脂肽產量和抑菌活性的影響,篩選出菌株產脂肽的最佳培養基配方。通過酸沉淀、醇溶法提取菌株FJAT-45028發酵產生的粗脂肽,利用液相色譜串聯四極桿飛行時間質譜(LC-QTOF-MS)測定脂肽組成和含量,并以龍眼致腐菌小新殼梭孢菌為指示菌,評估脂肽的抑菌活性。結果表明:貝萊斯芽胞桿菌FJAT-45028在供試的12種培養基中發酵后均能產生脂肽,粗脂肽產量范圍為2.28~4.86 g·L-1,抑菌圈直徑范圍為12.46~20.02 mm。LC-QTOF-MS分析結果顯示,菌株FJAT-45028產生的脂肽由伊枯草菌素、豐原素和表面活性素組成,不同培養基中產生的伊枯草菌素、豐原素和表面活性素的含量范圍分別是0.46~63.49 mg·L-1、34.07~918.86 mg·L-1和7.00~53.24 mg·L-1。相關性分析表明,伊枯草菌素和豐原素含量以及脂肽總量均與抑菌活性呈顯著正相關(P<0.01)。以抑菌活性和脂肽產量為指標,篩選出貝萊斯芽胞桿菌FJAT-45028產脂肽的最佳培養基配方為:蔗糖20 g·L-1、酵母浸膏20 g·L-1、牛肉浸膏15 g·L-1、MgSO4·7H2O 0.6 g·L-1、FeSO4·7H2O 9.0 mg·L-1。該研究篩選到了適于產抑菌脂肽的培養基,為菌株FJAT-45028抑菌脂肽的生產奠定了基礎。
關鍵詞:貝萊斯芽胞桿菌FJAT-45028;脂肽產量;培養基組分;抑菌活性
中圖分類號:S 476.1?? 文獻標志碼:A?? 文章編號:0253-2301(2021)12-0026-06
DOI: 10.13651/j.cnki.fjnykj.2021.12.005
Effect of Culture Medium Formula on Yield and Antibacterial Activity of theLipopeptides Produced by Bacillus velezensis FJAT-45028
CHEN Yan-ping, CHEN Mei-chun, LAN Jiang-lin, ZHENG Mei-xia, LIU Bo, WANG Jie-ping*
(Institute of Agricultural Biological Resources, Fujian Academy ofAgricultural Sciences, Fuzhou, Fujian 350003, China)
Abstract: In this study, effect of culture medium formula on yield and antibacterial activity of the lipopeptides produced by Bacillus velezensis FJAT-45028 was investigated to obtain the optimal medium for production of the antibacterial lipopeptides. The crude lipopeptides were extracted by acid precipitation and alcoholysis. Their component and content were determined using a liquid chromatography coupled with a quadrupole time-of-flight tandem mass spectrometry (LC-QTOF-MS). The antifungal activity of the lipopeptides was evaluated byusing Neofusicoccum parvum as an indicator, which is a rot-causing fungus of the Longan fruits. The results showed that the strain FJAT-45028 could yield lipopeptides in the twelve tested culture media. Moreover, content of the crude lipopeptides varied from 2.28 g·L-1 to 4.86 g·L-1, and the diameters of inhibition zone varied from 12.46 mm to 20.02 mm. The results of LC-QTOF-MS detection indicated that the lipopeptides produced by FJAT-45028 were composed by iturin, fengycin and surfactin, and their contents in the twelve culture media varied within the range of 0.46-63.49 mg·L-1, 34.07-918.86 mg·L-1, and 7.00-53.24 mg·L-1, respectively. The correlation analysis indicated that the contents of iturin, fengycin and total lipopeptides were significantly positive correlation with the antifungal activities (P<0.01). Finally, an optimal medium formula for production of the lipopeptides with antifungal activities by B. velezensis FJAT-45028 was as follow: Sucrose 20 g·L-1, yeast extract 20 g·L-1, beef extract 15 g·L-1, MgSO4·7H2O 0.6 g·L-1, FeSO4·7H2O 9.0 mg·L-1, according to the antifungal activities and yield of the lipopeptides. In conclusion,the suitable medium for production of the antifungal lipopeptides was established, which laying the foundation for the industrial production of the antifungal lipopeptides from the strain FJAT-45028.
Key words: Bacillus velezensis FJAT-45028; Yield of lipopeptide; Components of the culture medium; Antibacterial activity
芽胞桿菌脂肽具有廣譜抗菌活性、穩定性好、不易產生耐藥性、對人畜毒性低且對環境友好等優點,可用于防治水果采后病害。例如,枯草芽胞桿菌fmbJ產生的脂肽可延緩桃子的腐爛,防腐效率達76.5%[1];貝萊斯芽胞桿菌HN-2的脂肽對芒果炭疽菌具有強拮抗作用[2];解淀粉芽胞桿菌BA168產生的豐原素fengycin能夠有效抑制蘋果采后病害青霉病原菌的生長[3]。
芽胞桿菌脂肽是通過非核糖體途徑合成的,優化菌株培養條件,能夠大幅度提升脂肽產量。例如,枯草芽胞桿菌YHI在無碳源供應時,菌體幾乎不產脂肽,添加淀粉+米糠油為碳源后,脂肽產量比以葡萄糖為碳源高出104.4%[4];枯草芽胞桿菌T500在基礎培養基中添加蛋白胨和酵母粉進行優化后,表面活性素surfactin的產量提高了48.2%,伊枯草菌素iturin的產量提高了180.9%,同時還檢測到了基礎培養基發酵時無法產生的豐原素[5]。本實驗室篩選獲得了1株能夠有效抑制龍眼采后致腐病原菌小新殼梭孢菌Neofusicoccum parvum生長的貝萊斯芽胞桿菌FJAT-45028,該菌發揮拮抗作用的主要活性成分為脂肽。為提升該菌株的脂肽產量和抑菌活性,本研究利用液相色譜串聯四極桿飛行時間質譜(LC-QTOF-MS/MS)技術,分析了菌株FJAT-45028在12種不同培養基中的脂肽組成和產量,并結合抑菌活性,篩選出菌株FJAT-45028產脂肽的最佳培養基配方,為脂肽在龍眼采后病害的防控應用奠定基礎。
1 材料與方法
1.1 供試材料
供試菌株:貝萊斯芽胞桿菌Bacillus velezensis FJAT-45028、龍眼采后致腐菌小新殼梭孢菌Neofusicoccum parvum FJAT-3531,均由福建省農業科學院農業生物資源研究所實驗室分離、保存。
本試驗所需的培養基配方見表1,各培養基試劑均購買于北京奧博星生物技術有限責任公司。
1.2 試驗方法
1.2.1 菌株培養 (1)貝萊斯芽胞桿菌FJAT-45028培養:菌株FJAT-45028接種至50 mL LB液體培養基中,30℃、170 r·min-1 振蕩培養24 h,作為種子液。按1%接種量接種至50 mL不同的培養基中(表1),25℃、170 r·min-1 振蕩培養48 h。(2)小新殼梭孢菌FJAT-3531培養:FJAT-3531接種至50 mL的PDA培養基中,30℃、170 r·min-1培養48 h,濃度調整為107 CFU·mL-1備用。
1.2.2 脂肽制備 貝萊斯芽胞桿菌FJAT-45028發酵液離心棄去菌體得上清液,上清液加2 mol·L-1鹽酸至pH<2,4℃靜置24 h,離心得沉淀,加入磷酸鹽緩沖溶液(PBS)溶解后,經低溫真空冷凍干燥后為粗脂肽粉末,稱重記錄所得粗脂肽產量。
1.2.3 脂肽組成測定 參照文獻Chen等[6]方法,利用LC-QTOF-MS/MS技術進行脂肽組成和含量測定。脂肽iturin、fengycin和surfactin的含量定義為每升上清液所含有的脂肽量(mg·L-1)。
1.2.4 抑菌活性檢測 采用抑菌圈法進行貝萊斯芽胞桿菌FJAT-45028脂肽對小新殼梭孢菌FJAT-3531的抑菌活性測定。吸取準備好的病原菌懸浮液適量,加入到50℃左右的半固體PDA培養基中(瓊脂含量0.9%),使得病原菌終濃度為5×105 CFU·mL-1,并倒置于已凝固的PDA固體培養基上(瓊脂含量1.8%)。待培養基凝固后,采用直徑為7 mm的打孔器進行打孔,分別在孔中注入甲醇溶解的濃度為30 mg·mL-1的脂肽溶液80 μL,無菌甲醇溶液為空白對照,以0.5 mg·mL-1的潮霉素為陽性對照,每個處理重復3次,30℃培養3 d后,測抑菌圈直徑。
1.2.5 數據分析 采用DPS 18.10軟件,Duncan新復極差法進行方差分析,多元回歸法進行脂肽含量和抑菌活性相關性分析。
2 結果與分析
2.1 培養基配方對貝萊斯芽胞桿菌FJAT-45028粗脂肽產量的影響
由表2可知,菌株FJAT-45028在12種培養基中均能產生脂肽,不同培養基的粗脂肽產量范圍為2.28~4.86 g·L-1,其中最高產量的是最低產量的2.13倍;菌株FJAT-45028在K、L、F和C這4種培養基中的粗脂肽產量顯著高于其他培養基(P<0.05),這4種培養基的脂肽產量無顯著差異(P>0.05);而利用H和B培養基發酵的脂肽產量最低,僅為2.31 g·L-1和2.28 g·L-1。對培養基組分進行分析,發現在培養基中添加甘油或山梨醇能顯著提高脂肽的產量。
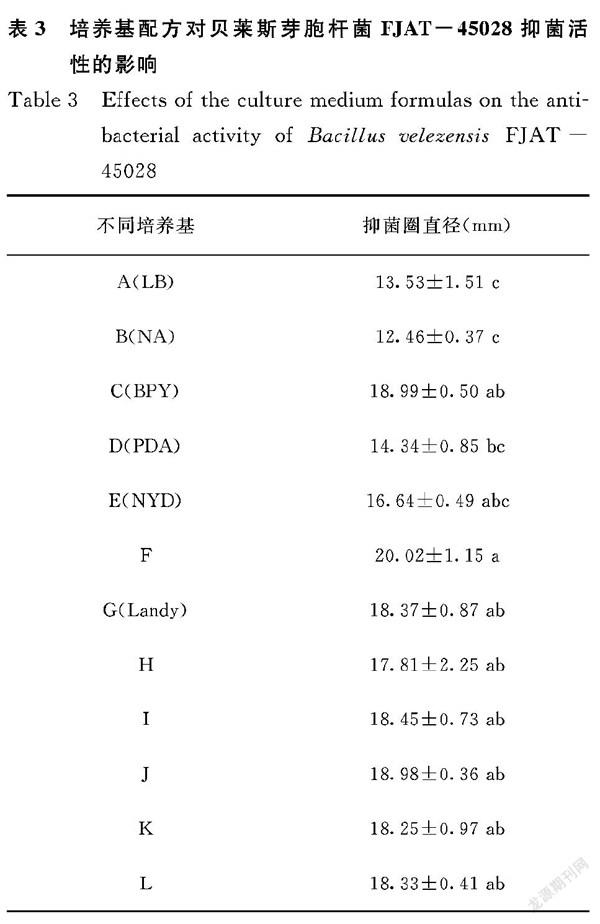
2.2 培養基配方對貝萊斯芽胞桿菌FJAT-45028抑菌活性的影響
由表3可知,菌株FJAT-45028在不同培養基中產生的脂肽對龍眼致腐菌小新殼梭孢菌均有抑菌活性,其抑菌圈直徑范圍為12.46~20.02 mm;菌株FJAT-45028在培養基F中產生的脂肽抑菌活性顯著高于其他培養基(P<0.05),抑菌圈直徑為20.02 mm;其次,其他培養基所產脂肽抑菌活性僅次于培養基F的有C、J、I、G、L、K、H,該菌在這7組培養基中脂肽抑菌活性無顯著性差異(P>0.05);而利用培養基A和B所產脂肽的抑菌活性最差,抑菌圈直徑僅為13.53 mm和12.46 mm。對培養基組分進行分析,發現富含碳源、氮源和無機鹽的培養基產生的脂肽具有較好的抑菌效果。綜合考慮粗脂肽產量和抑菌活性,篩選的貝萊斯芽胞桿菌FJAT-45028產脂肽最佳培養基配方為F培養基,即蔗糖20 g·L-1、酵母浸膏20 g·L-1、牛肉浸膏15 g·L-1、MgSO4·7H2O 0.6g·L-1、FeSO4·7H2O 9.0 mg·L-1。
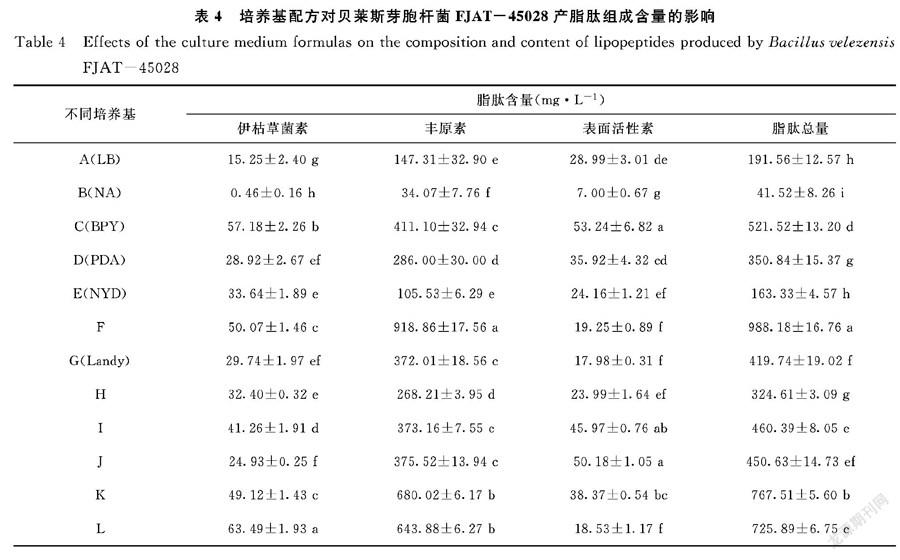
2.3 培養基配方對貝萊斯芽胞桿菌FJAT-45028產脂肽組成含量的影響
從表4可知,12種培養基產生的脂肽均含有伊枯草菌素、豐原素、表面活性素三大家族化合物,脂肽總含量范圍在41.52~988.18 mg·L-1,最大含量是最低含量的23.8倍。伊枯草菌素含量范圍在0.46~63.49 mg·L-1,其中菌株FJAT-45028在培養基中伊枯草菌素含量大小前四者的依次為L>C>F>K;豐原素含量范圍在34.07~918.86 mg·L-1,各培養基中豐原素含量大小前四者的依次為F>K>L>C;表面活性素含量范圍在7.00~53.24 mg·L-1,各培養基中表面活性素含量大小前四者的依次為C>J>I>K。分析發現,伊枯草菌素和豐原素含量在前四者的培養基均為F、K、L、C,伊枯草菌素、豐原素、表面活性素含量最低者均為B培養基,說明富含碳源、氮源及無機鹽的培養基有利于貝萊斯芽胞桿菌FJAT-45028脂肽組分中伊枯草菌素和豐原素的合成。
2.4 貝萊斯芽胞桿菌FJAT-45028脂肽組分含量與抑菌活性相關性分析
脂肽對龍眼致腐菌小新殼梭孢菌FJAT-3531的抑菌活性與伊枯草菌素含量、豐原素含量及總脂肽含量間相關系數分別為0.7678**、0.7351**和0.7625**,均呈極顯著正相關(P<0.01),而與表面活性素含量相關系數為0.3759,兩者間相關性不顯著(P>0.05),表明菌株FJAT-3531粗脂肽中發揮抑菌作用的活性成分可能為伊枯草菌素和豐原素。
3 結論與討論
脂肽是由非核糖體途徑合成,不同菌株所產脂肽組分不一樣,但同一菌株在不同的培養條件下所產脂肽組分和含量也會有所不同。李興玉等[7]報道枯草芽胞桿菌XF1菌株在5種不同的培養基發酵液中所含環脂肽類化合物色譜圖基本一致,但脂肽產量最高者為最低者的2.94倍。張曉云等[8]分析了不同碳、氮源對枯草芽胞桿菌BAB-1菌株脂肽產量的影響,結果表明,熊果苷是BAB-1菌株產生表面活性素的最佳碳源,其含量是L阿拉伯糖的5.60倍;熊果苷與葡萄糖利于該菌株產生豐原素,顯著高于D木糖、D核糖及L阿拉伯糖。本研究結果顯示,貝萊斯芽胞桿菌FJAT-45028在12種培養基中所產的脂肽組分均含有伊枯草菌素、豐原素、表面活性素,但各組分的含量均不同,且差異顯著。
金屬離子對抑菌脂肽的產生有一定影響,鐵、鎂、錳離子能促進抑菌脂肽的產生。劉宇帥等[9]研究表明,較高濃度的葡萄糖和無機鹽及較低濃度的Mg2+能夠顯著提高解淀粉芽胞桿菌TF28抑菌脂肽的產量;黃翔峰等[10]研究發現,添加Fe2+能大幅提高脂肽產量。本研究發現,FJAT-45028菌株在加入Mg2+和Fe2+的F和K培養基產生的脂肽含量顯著高于未添加Mg2+和Fe2+的A和B培養基,其中F發酵液中脂肽含量為B培養基的23.8倍,說明Mg2+和Fe2+也能促進FJAT-45028菌株的脂肽的合成。
研究表明,豐原素和伊枯草菌素具有與化學農藥相當的生物活性,能顯著抑制植物病原真菌的生長。付瑞敏等[3]研究發現,解淀粉芽胞桿菌BA168產生的豐原素是抑制蘋果采后病害擴展青霉病原菌的脂肽成分;Calvo等[11]研究表明,解淀粉芽胞桿菌BUZ14脂肽伊枯草菌素A是防控桃子褐腐病和蘋果藍腐病的主要抑菌成分;蔣夢曦等[12]用脂肽伊枯草菌素A處理櫻桃番茄后,腐爛率可降低55%;朱弘元等[13]研究發現,解淀粉芽胞桿菌B15生產的伊枯草菌素A、豐原素對葡萄灰霉病菌有較好的抑菌效果。本研究結果顯示,菌株FJAT-45028粗脂肽對龍眼致腐菌小新殼梭孢菌的抑制活性與伊枯草菌素和豐原素呈極顯著正相關,表明粗脂肽發揮抑菌作用的活性組分可能為伊枯草菌素和豐原素,該結果與前人報道相吻合。綜合考慮脂肽產量和抑菌活性,貝萊斯芽胞桿菌FJAT-45028產脂肽的最佳培養基配方為蔗糖20 g·L-1、酵母浸膏20 g·L-1、牛肉浸膏15 g·L-1、MgSO4·7H2O 0.6 g·L-1、FeSO4·7H2O 9.0 mg·L-1。本研究結果為脂肽在龍眼采后病害防控的應用奠定基礎。
參考文獻:
[1]黃現青,別小妹,呂鳳霞,等.枯草芽胞桿菌fmbJ產脂肽抑制點青霉效果及其桃防腐試驗[J].農業工程學報,2008(1):263-267.
[2]王雨,譚崢,韋丹丹,等.貝萊斯芽胞桿菌HN2的鑒定.及對杧果炭疽菌的抑菌活性研究[J].中國生物防治學報,2020,36(2):220-230.
[3]付瑞敏,常慧萍,邢文會,等.解淀粉芽胞桿菌所產脂肽豐原素是防治蘋果采后青霉病的關鍵物質[J].江蘇農業科學,2018,46(3):80-85.
[4]沈玉江,雷亞峰,劉健,等.產表面活性素枯草芽胞桿菌YHI的培養基優化研究[J].黑龍江八一農墾大學學報,2020,32(5):69-76,128.
[5]喬俊卿,劉郵洲,張榮勝,等.枯草芽胞桿菌T500產脂肽類抗生素的搖瓶發酵工藝優化[J].南京農業大學學報,2018,41(3):447-455.
[6]CHEN M C,WANG J P,ZHU Y J,et al.Antibacterial activity against Ralstonia solanacearum of the lipopeptides secreted from the Bacillus amyloliquefaciens strain FJAT-2349[J].Journal of Applied Microbiology,2019,126(5):1519-1529.
[7]李興玉,毛自朝,吳毅歆,等.不同培養基對Bacillus subtilis XF1產脂肽抗生素的影響[J].內蒙古農業大學學報(自然科學版),2014,35(6):49-53.
[8]張曉云,郭慶港,王培培,等.不同碳氮源對枯草芽胞桿菌BAB1產抗菌脂肽的影響[J].中國生物防治學報,2021,37(2):251-258.
[9]劉宇帥,孟利強,陳靜宇,等.解淀粉芽胞桿菌TF28產抗菌脂肽培養基優化[J].微生物學雜志,2017,37(3):52-58.
[10]黃翔峰,詹鵬舉,彭開銘,等.培養基中鐵離子對枯草芽胞桿菌CICC 23659發酵產脂肽的影響研究[J].中國生物工程雜志,2013,33(6):52-61.
[11]CALVO H,MENDIARA I,ARIAS E,et al.The role of iturin A from B.amyloliquefaciens BUZ14 in the inhibition of the most common postharvest fruit rots[J].Food Microbiol,2019,82:62-69.
[12]蔣夢曦,林福興,別小妹,等.Iturin A與肉桂精油復配提升櫻桃番茄貯藏品質[J].食品與發酵工業,2019,45(19):206-212.
[13]朱弘元,康健,范昕,等.解淀粉芽胞桿菌B15產脂肽的分離鑒定及抑菌機理[J].江蘇農業科學,2016,44(5):186-189.
(責任編輯:柯文輝)
收稿日期:2021-10-12
作者簡介:陳燕萍,女,1985年生,碩士,助理研究員,主要從事生物防治研究。
通信作者:王階平,男,1972年生,博士,研究員,主要從事微生物生物技術與農業生物藥物(E-mail:781063449@qq.com)。
基金項目:福建省科技計劃項目——省屬公益類科研院所基本科研專項(2019R1034-5、2019R1034-2);福建省農業科學院對外合作項目(DEC2020-06)。
